
From Morphology to Multi-Omics: A New Age of Fusarium Research

 , , , , and
, , , , and
Abstract
1. Fusarium: A Genus of Global Agricultural and Phytopathological Significance
2. Taxonomy and Phylogenetics of Fusarium Species
2.1. From Morphology to Genomics: Evolution of Fusarium Classification
2.2. Rise and Relevance of Fusarium Species Complexes: Evolutionary Divergence, Taxonomic Challenges, Intraspecific Variability and Pathogenic Implications
2.3. The Neocosmospora Debate: Taxonomy Reshuffled, or Genus Redefined?
2.4. Emerging Fusarium Pathogens in Niche Crops and Geographies

{kind=link}
{kind=link}
| Fusarium Species | Species Complex | Primary Host(s) | Crop Type | Region | References |
|---|---|---|---|---|---|
| F. zanthoxyli | FTOSC | Zanthoxylum bungeanum | Spice crop (woody) | Northern China | [67,68] |
| F. torreyae | FTOSC | Torreya taxifolia | Tree (conifer) | North America | [69] |
| F. oxysporum f. sp. cubense TR4 | FOSC | Banana (Musa spp.) | Fruit crop (perennial herb) | Southeast Asia, Middle East, Africa, Latin America | [71,72,73] |
| Novel FIESC spp. | FIESC | Various herbs and crops | Medicinal, legumes | China | [76] |
| FIESC spp. | FIESC | Rice, soybean, legumes | Cereals, legumes | South America, Africa, Europe | [77,79,80] |
| F. acuminatum, F. avenaceum, F. tricinctum | FTSC | Soybean, medicinal herbs, fruits, cereals | Multiple | Asia, Europe | [80,82,86] |
| F. avenaceum | FTSC | Raspberry, pulses, cereals, soybean | Berries, legumes | Europe, North-Estern China | [7,80,85,87] |
2.5. Phylogenomics of Fusarium: Insights from Whole-Genome Data
3. Genomic Insights and Functional Genomics: Exploring Recent Genomic Studies and Their Implications for Understanding Pathogenicity and Resistance Mechanisms
3.1. Genomic Architecture of Fusarium Species
3.2. Secondary Metabolites, Genome Plasticity, and Host Specialization
3.2.1. Functional Genomics and Pathogenicity Mechanisms
3.2.2. Proteomics and Secreted Virulence Factors
3.3. Resistance Mechanisms in Host Plants
3.4. Phylogenomics and Comparative Genome Analysis Across Species Complexes
4. Pathogenicity and Host Interactions
4.1. Fusarium Pathogenicity
4.2. Host-Specificity and Formae Speciales in Fusarium Species
4.3. Plant Immune Responses and Fusarium Counter-Defenses
5. Toxins Produced by Fusarium: Global Burden in Major Staple Crops
5.1. Overview of Major Fusarium-Derived Toxins in Staple Crops
5.2. Regional Analysis of Mycotoxin Burden and Exposure Risk
5.3. Mycotoxin Detection
6. Management and Diagnostic Strategies of Fusarium-Induced Plant Diseases
| Fungicide (Common Name) | Commercial Name (If Specificied) | FRAC Category/Chemical Group | Mode of Application | Target Fusarium Species/Disease | References |
|---|---|---|---|---|---|
| Azoxystrobin | Azimut, Amistar, Dynasty, Ortiva | Strobilurin (QoI, FRAC group 11) | Seed coating, Culture plates, Seed treatment, Foliar spray, in vitro | F. acutatum, F. oxysporum f. sp. cepae, Fusarium spp., F. virguliforme, F. subglutinans, F. temperatum, F. graminearum, F. oxysporum, F. pseudograminearum | [200,234,235,236] |
| Carbendazim | Antracol | Benzimidazole (Systemic, Broad-spectrum) | Seed treatment, Soil mixture, Culture media | Fusarium spp., F. oxysporum f. sp. vasinfectum, F. oxysporum f. sp. lentis, F. oxysporum (maize), F. equiseti, F. chlamydosporum, F. pseudograminearum | [227,228,229,230,231,232,233,234,235,236,237,238,239] |
| Cyclobutrifluram | - | SDHI (FRAC group 7) | In vitro | F. pseudograminearum | [240] |
| Difenoconazole | Dividend XL RTA | Azole (DMI) | Culture plates, Seed treatment, in vitro | F. solani, F. proliferatum, F. oxysporum, F. circinatum, F. avenaceum, F. culmorum, F. poae, F. sporotrichioides, F. subglutinans, F. temperatum, F. pseudograminearum | [241,242,243,244] |
| Fludioxonil | Vibrance, Maxim 4 FS, MaximQuattro, Celest XL, Celest Quattro | Phenylpyrrole | Seed coating, Culture plates, Seed treatment | F. acutatum, F. oxysporum f. sp. cepae, Rhizoctonia, Fusarium spp., F. graminearum, F. virguliforme, F. solani, F. oxysporum (dry rot), Sclerotinia sclerotiorum, F. verticillioides, F. pseudograminearum | [200,213,223] |
| Fluopyram | ILeVO | SDHI (FRAC group 7) | Seed treatment | F. virguliforme | [221] |
| Phenamacril | - | Myosin inhibitor | In vitro | F. pseudograminearum | [226,240] |
| Prochloraz | Sportak | Azole (DMI) | Seed coating, Potted sprout irrigation, Culture plates | F. oxysporum f. sp. cepae, F. acutatum, F. subglutinans, F. temperatum, F. oxysporum (banana wilt), F. graminearum (FHB), F. culmorum (FHB), F. oxysporum f. sp. lycopersici, F. pseudograminearum | [235,244] |
| Prothioconazole | Redigo, Proline, Prosaro | Azole (DMI, FRAC group 3) | Seed treatment, Foliar fungicide, in vitro | Fusarium spp., F. virguliforme, F. graminearum (FHB), F. poae, F. pseudograminearum | [222,245,246] |
| Pyraclostrobin | Stamina, BAS 580 | Strobilurin (QoI, FRAC group 11) | Seed treatment, Foliar fungicide, in vitro | Fusarium spp., F. virguliforme, F. graminearum, F. pseudograminearum | [223,240,245] |
| Tebuconazole | Orius 25, Azimut, Raxil 250 FL, Raxil MD, Raxil T, Folicur, Nativo SC300, Twinstar 75 WG, Prosaro | Azole (DMI, FRAC group 3) | Culture plates, Seed coating, In vitro, Foliar spray | F. acutatum, F. oxysporum f. sp. cepae, F. subglutinans, F. temperatum, F. graminearum (FHB), F. culmorum (FHB), F. poae, Fusarium equiseti, F. chlamydosporum, Fusarium spp., F. pseudograminearum | [243,244,247] |
| Thiabendazole | Mertect 340F, Rival, Tecto, MaximQuattro, Trilex AL (part of) | Benzimidazole (FRAC group 1) | Culture plates, Seed treatment, In vitro, In vivo, In situ | F. solani, Fusarium spp., F. oxysporum, F. graminearum (part of combination), F. verticillioides (part of combination) | [241,248] |
| Trifloxystrobin | Trilex, Fortix | Strobilurin (QoI, FRAC group 11) | Seed treatment, Foliar fungicide, in vitro | Fusarium spp., F. graminearum, F. virguliforme, F. chlamydosporum, F. asiaticum | [243,247,249] |
Molecular Diagnostics
7. Conclusions and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Aoki, T.; O’Donnell, K.; Geiser, D.M. Systematics of key phytopathogenic Fusarium species: Current status and future challenges. J. Gen. Plant Pathol. 2014, 80, 189–201. [Google Scholar] [CrossRef]
- O’Donnell, K.; Whitaker, B.K.; Laraba, I.; Proctor, R.H.; Brown, D.W.; Broders, K.; Kim, H.-S.; McCormick, S.P.; Busman, M.; Aoki, T. DNA sequence-based identification of Fusarium: A work in progress. Plant Dis. 2022, 106, 1597–1609. [Google Scholar] [CrossRef]
- Kamil, D.; Mishra, A.K.; Das, A.; Nishmitha, K. Genus Fusarium and Fusarium species complexes. In Biodiversity, Bioengineering, and Biotechnology of Fungi; Academic Press: Cambridge, MA, USA, 2025; pp. 209–225. [Google Scholar] [CrossRef]
- Torbati, M.; Arzanlou, M.; da Silva Santos, A.C. Fungicolous Fusarium species: Ecology, diversity, isolation, and identification. Curr. Microbiol. 2021, 78, 2850–2859. [Google Scholar] [CrossRef]
- Armer, V.J.; Kroll, E.; Darino, M.; Smith, D.P.; Urban, M.; Hammond-Kosack, K.E. Navigating the Fusarium species complex: Host-range plasticity and genome variations. Fungal Biol. 2024, 128, 2439–2459. [Google Scholar] [CrossRef]
- Khuna, S.; Kumla, J.; Thitla, T.; Nuangmek, W.; Lumyong, S.; Suwannarach, N. Morphology, molecular identification, and pathogenicity of two novel Fusarium species associated with postharvest fruit rot of cucurbits in Northern Thailand. J. Fungi 2022, 8, 1135. [Google Scholar] [CrossRef]
- Moparthi, S.; Burrows, M.; Mgbechi-Ezeri, J.; Agindotan, B. Fusarium spp. associated with root rot of pulse crops and their cross-pathogenicity to cereal crops in Montana. Plant Dis. 2021, 105, 548–557. [Google Scholar] [CrossRef]
- Moparthi, S.; Perez-Hernandez, O.; Burrows, M.E.; Bradshaw, M.J.; Bugingo, C.; Brelsford, M.; McPhee, K. Identification of Fusarium spp. associated with chickpea root rot in Montana. Agriculture 2024, 14, 974. [Google Scholar] [CrossRef]
- Dean, R.; van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J. The top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef]
- Ekwomadu, T.I.; Akinola, S.A.; Mwanza, M. Fusarium mycotoxins, their metabolites (free, emerging, and masked), food safety concerns, and health impacts. Int. J. Environ. Res. Public Health 2021, 18, 11741. [Google Scholar] [CrossRef]
- Okungbowa, F.I.; Shittu, H.O. Fusarium wilts: An overview. Environ. Res. J. 2012, 6, 83–102. [Google Scholar]
- Arie, T. Fusarium diseases of cultivated plants, control, diagnosis, and molecular and genetic studies. J. Pestic. Sci. 2019, 44, 275–281. [Google Scholar] [CrossRef]
- Ma, L.-J.; Geiser, D.M.; Proctor, R.H.; Rooney, A.P.; O’Donnell, K.; Trail, F.; Gardiner, D.M.; Manners, J.M.; Kazan, K. Fusarium pathogenomics. Annu. Rev. Microbiol. 2013, 67, 399–416. [Google Scholar] [CrossRef]
- Coleman, J.J. The Fusarium solani species complex: Ubiquitous pathogens of agricultural importance. Mol. Plant Pathol. 2016, 17, 146–158. [Google Scholar] [CrossRef]
- Chandra, N.S.; Wulff, E.G.; Udayashankar, A.C.; Nandini, B.P.; Niranjana, S.R.; Mortensen, C.N.; Prakash, H.S. Prospects of molecular markers in Fusarium species diversity. Appl. Microbiol. Biotechnol. 2011, 90, 1625–1639. [Google Scholar] [CrossRef]
- Infantino, A.; Grottoli, A.; Bergamaschi, V.; Oufensou, S.; Burgess, L.W.; Balmas, V. FusaHelp: A web site program for the morphological identification of Fusarium species. J. Plant Pathol. 2023, 105, 429–436. [Google Scholar] [CrossRef]
- O’Donnell, K.; Ward, T.J.; Robert, V.A.; Crous, P.W.; Geiser, D.M.; Kang, S. DNA sequence-based identification of Fusarium: Current status and future directions. Phytoparasitica 2015, 43, 583–595. [Google Scholar] [CrossRef]
- Desai, S.; Dubey, S.C.; Prasad, R.D. Impacts of climate change on Fusarium species vis-à-vis adaptation strategies. Indian Phytopathol. 2020, 73, 593–603. [Google Scholar] [CrossRef]
- Buxton, E.W. The taxonomy and variation in culture of Fusarium oxysporum from gladiolus. Trans. Br. Mycol. Soc. 1955, 38, 202–208. [Google Scholar] [CrossRef]
- Snyder, W.C.; Hansen, H.N. The species concept in Fusarium. Am. J. Bot. 1940, 27, 64–67. [Google Scholar] [CrossRef]
- Booth, C. The Genus Fusarium; Commonwealth Mycological Institute: Kew, Surrey, 1971; Volume 237. [Google Scholar]
- O’Donnell, K.; Sutton, D.A.; Rinaldi, M.G.; Sarver, B.A.; Balajee, S.A.; Schroers, H.-J.; Summerbell, R.C.; Robert, V.A.; Crous, P.W.; Zhang, N. Internet-accessible DNA sequence database for identifying fusaria from human and animal infections. J. Clin. Microbiol. 2010, 48, 3708–3718. [Google Scholar] [CrossRef]
- Watanabe, M.; Yonezawa, T.; Lee, K.-i; Kumagai, S.; Sugita-Konishi, Y.; Goto, K.; Hara-Kudo, Y. Molecular phylogeny of the higher and lower taxonomy of the Fusarium genus and differences in the evolutionary histories of multiple genes. BMC Evol. Biol. 2011, 11, 322. [Google Scholar] [CrossRef]
- Lizcano Salas, A.F.; Duitama, J.; Restrepo, S.; Celis Ramírez, A.M. Phylogenomic approaches reveal a robust time-scale phylogeny of the Terminal Fusarium Clade. IMA Fungus 2024, 15, 13. [Google Scholar] [CrossRef]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W.; Fungal Barcoding Consortium. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef]
- Singha, I.M.; Kakoty, Y.; Unni, B.G.; Das, J.; Kalita, M.C. Identification and characterization of Fusarium spp. using ITS and RAPD causing fusarium wilt of tomato isolated from Assam, Northeast India. J. Genet. Eng. Biotechnol. 2016, 14, 99–105. [Google Scholar] [CrossRef]
- Zarrin, M.; Ganj, F.; Faramarzi, S. Analysis of the rDNA internal transcribed spacer region of the Fusarium species by polymerase chain reaction-restriction fragment length polymorphism. Biomed. Rep. 2016, 4, 471–474. [Google Scholar] [CrossRef]
- Crous, P.W.; Lombard, L.; Sandoval-Denis, M.; Seifert, K.A.; Schroers, H.-J.; Chaverri, P.; Gené, J.; Guarro, J.; Hirooka, Y.; Bensch, K. Fusarium: More than a node or a foot-shaped basal cell. Stud. Mycol. 2021, 98, 100116. [Google Scholar] [CrossRef]
- van Diepeningen, A.D.; Feng, P.; Ahmed, S.; Sudhadham, M.; Bunyaratavej, S.; de Hoog, G.S. Spectrum of Fusarium infections in tropical dermatology evidenced by multilocus sequencing typing diagnostics. Mycoses 2015, 58, 48–57. [Google Scholar] [CrossRef]
- Oliveira, L.J.M.G.; Rodrigues, A.A.C.; Silva, E.K.C.; Oliveira, A.C.S.; Barros, M.C.; Fraga, E.C.; Nascimento, I.O.; Silva, M.R.M. Morphological and phylogenetic characterization of Fusarium Link. Aust. J. Crop Sci. 2021, 15, 1406–1415. [Google Scholar] [CrossRef]
- Yörük, E.; Yli-Mattila, T. Translation elongation factor 1-alpha sequencing provides reliable tool for identification of Fusarium graminearum species complex members. Diversity 2024, 16, 481. [Google Scholar] [CrossRef]
- Geiser, D.M.; Al-Hatmi, A.M.; Aoki, T.; Arie, T.; Balmas, V.; Barnes, I.; Bergstrom, G.C.; Bhattacharyya, M.K.; Blomquist, C.L.; Bowden, R.L. Phylogenomic analysis of a 55.1-kb 19-gene dataset resolves a monophyletic Fusarium that includes the Fusarium solani species complex. Phytopathology 2021, 111, 1064–1079. [Google Scholar] [CrossRef]
- Sandoval-Denis, M.; Lombard, L.; Crous, P.W. Back to the roots: A reappraisal of Neocosmospora. Persoonia 2019, 43, 90–185. [Google Scholar] [CrossRef]
- Maryani, N.; Sandoval-Denis, M.; Lombard, L.; Crous, P.W.; Kema, G. New endemic Fusarium species hitch-hiking with pathogenic Fusarium strains causing Panama disease in small-holder banana plots in Indonesia. Persoonia 2019, 43, 48–69. [Google Scholar] [CrossRef]
- O’Donnell, K.; Rooney, A.P.; Proctor, R.H.; Brown, D.W.; McCormick, S.P.; Ward, T.J.; Frandsen, R.J.; Lysøe, E.; Rehner, S.A.; Aoki, T. Phylogenetic analyses of RPB1 and RPB2 support a middle Cretaceous origin for a clade comprising all agriculturally and medically important fusaria. Fungal Genet. Biol. 2013, 52, 20–31. [Google Scholar] [CrossRef]
- Manukyan, I.; Khatsaeva, R.; Kosyrev, S.; Nagam, M.A.A. Species diversity of Fusarium fungi on grain crops in the conditions of the foothill zone of the Central Caucasus. BIO Web Conf. 2024, 118, 01018. [Google Scholar] [CrossRef]
- Wang, M.; Crous, P.W.; Sandoval-Denis, M.; Han, S.; Liu, F.; Liang, J.; Duan, W.; Cai, L. Fusarium and allied genera from China: Species diversity and distribution. Persoonia 2022, 48, 1–53. [Google Scholar] [CrossRef]
- Laurence, M.H.; Summerell, B.A.; Burgess, L.W.; Liew, E.C. Genealogical concordance phylogenetic species recognition in the Fusarium oxysporum species complex. Fungal Biol. 2014, 118, 374–384. [Google Scholar] [CrossRef]
- Mi, Z.; Su, J.; Yu, L.; Zhang, T. Comparative mitochondrial genomics of Thelebolaceae in Antarctica: Insights into their extremophilic adaptations and evolutionary dynamics. IMA Fungus 2024, 15, 33. [Google Scholar] [CrossRef]
- Peck, L.D.; Llewellyn, T.; Bennetot, B.; O’Donnell, S.; Nowell, R.W.; Ryan, M.J.; Flood, J.; de la Vega, R.C.R.; Ropars, J.; Giraud, T. Horizontal transfers between fungal Fusarium species contributed to successive outbreaks of coffee wilt disease. PLoS Biol. 2024, 22, e3002480. [Google Scholar] [CrossRef]
- Lombard, L.; Sandoval-Denis, M.; Cai, L.; Crous, P.W. Changing the game: Resolving systematic issues in key Fusarium species complexes. Persoonia 2019, 43, i–ii. [Google Scholar] [CrossRef]
- O’Donnell, K.; Humber, R.A.; Geiser, D.M.; Kang, S.; Park, B.; Robert, V.A.; Crous, P.W.; Johnston, P.R.; Aoki, T.; Rooney, A.P. Phylogenetic diversity of insecticolous fusaria inferred from multilocus DNA sequence data and their molecular identification via FUSARIUM-ID and Fusarium MLST. Mycologia 2012, 104, 427–445. [Google Scholar] [CrossRef]
- Nilsson, R.H.; Larsson, K.-H.; Taylor, A.F.S.; Bengtsson-Palme, J.; Jeppesen, T.S.; Schigel, D.; Kennedy, P.; Picard, K.; Glöckner, F.O.; Tedersoo, L. The UNITE database for molecular identification of fungi: Handling dark taxa and parallel taxonomic classifications. Nucleic Acids Res. 2019, 47, D259–D264. [Google Scholar] [CrossRef]
- Villani, A.; Proctor, R.H.; Kim, H.-S.; Brown, D.W.; Logrieco, A.F.; Amatulli, M.T.; Moretti, A.; Susca, A. Variation in secondary metabolite production potential in the Fusarium incarnatum-equiseti species complex revealed by comparative analysis of 13 genomes. BMC Genom. 2019, 20, 314. [Google Scholar] [CrossRef]
- Ha, M.S.; Ryu, H.; Ju, H.J.; Choi, H.-W. Diversity and pathogenic characteristics of the Fusarium species isolated from minor legumes in Korea. Sci. Rep. 2023, 13, 22516. [Google Scholar] [CrossRef]
- Summerell, B.A. Resolving Fusarium: Current status of the genus. Annu. Rev. Phytopathol. 2019, 57, 323–339. [Google Scholar] [CrossRef]
- Gao, Y.; Zhang, Z.; Ji, M.; Ze, S.; Wang, H.; Yang, B.; Hu, L.; Zhao, N. Identification and pathogenicity of Fusarium species from herbaceous plants on grassland in Qiaojia County, China. Microorganisms 2025, 13, 113. [Google Scholar] [CrossRef]
- Harish, J.; Jambhulkar, P.P.; Bajpai, R.; Arya, M.; Babele, P.K.; Chaturvedi, S.K.; Kumar, A.; Lakshman, D.K. Morphological characterization, pathogenicity screening, and molecular identification of Fusarium spp. isolates causing post-flowering stalk rot in maize. Front. Microbiol. 2023, 14, 1121781. [Google Scholar] [CrossRef]
- Pothiraj, G.; Hussain, Z.; Singh, A.K.; Solanke, A.U.; Aggarwal, R.; Ramesh, R.; Shanmugam, V. Characterization of Fusarium spp. inciting vascular wilt of tomato and its management by a Chaetomium-based biocontrol consortium. Front. Plant Sci. 2021, 12, 748013. [Google Scholar] [CrossRef]
- Zhang, L.; Hou, M.; Zhang, X.; Cao, Y.; Sun, S.; Zhu, Z.; Han, S.; Chen, Y.; Ku, L.; Duan, C. Integrative transcriptome and proteome analysis reveals maize responses to Fusarium verticillioides infection inside the stalks. Mol. Plant Pathol. 2023, 24, 693–710. [Google Scholar] [CrossRef]
- Achari, S.R.; Kaur, J.; Dinh, Q.; Mann, R.; Sawbridge, T.; Summerell, B.A.; Edwards, J. Phylogenetic relationship between Australian Fusarium oxysporum isolates and resolving the species complex using the multispecies coalescent model. BMC Genom. 2020, 21, 248. [Google Scholar] [CrossRef]
- Brankovics, B.; van Dam, P.; Rep, M.; de Hoog, G.S.; van der Lee, T.A.J.; Waalwijk, C.; van Diepeningen, A.D. Mitochondrial genomes reveal recombination in the presumed asexual Fusarium oxysporum species complex. BMC Genom. 2017, 18, 735. [Google Scholar] [CrossRef]
- Coleman, J.J.; Rounsley, S.D.; Rodriguez-Carres, M.; Kuo, A.; Wasmann, C.C.; Grimwood, J.; Schmutz, J.; Taga, M.; White, G.J.; Zhou, S. The genome of Nectria haematococca: Contribution of supernumerary chromosomes to gene expansion. PLoS Genet. 2009, 5, e1000618. [Google Scholar] [CrossRef]
- Homa, M.; Galgóczy, L.; Manikandan, P.; Narendran, V.; Sinka, R.; Csernetics, Á.; Vágvölgyi, C.; Kredics, L.; Papp, T. South Indian isolates of the Fusarium solani species complex from clinical and environmental samples: Identification, antifungal susceptibilities, and virulence. Front. Microbiol. 2018, 9, 1052. [Google Scholar] [CrossRef]
- Šišić, A.; Baćanović-Šišić, J.; Al-Hatmi, A.M.; Karlovsky, P.; Ahmed, S.A.; Maier, W.; de Hoog, G.S.; Finckh, M.R. The ‘forma specialis’ issue in Fusarium: A case study in Fusarium solani f. sp. pisi. Sci. Rep. 2018, 8, 1252. [Google Scholar] [CrossRef]
- Moussa, T.A.A.; Al-Zahrani, H.S.; Kadasa, N.M.S.; Ahmed, S.A.; de Hoog, G.S.; Al-Hatmi, A.M.S. Two new species of the Fusarium fujikuroi species complex isolated from the natural environment. Antonie Van Leeuwenhoek 2017, 110, 819–832. [Google Scholar] [CrossRef]
- Geiser, D.M.; Aoki, T.; Bacon, C.W.; Baker, S.E.; Bhattacharyya, M.K.; Brandt, M.E.; Brown, D.W.; Burgess, L.W.; Chulze, S.; Coleman, J.J. One fungus, one name: Defining the genus Fusarium in a scientifically robust way that preserves longstanding use. Phytopathology 2013, 103, 400–408. [Google Scholar] [CrossRef]
- Inami, K.; Yoshioka-Akiyama, C.; Morita, Y.; Yamasaki, M.; Teraoka, T.; Arie, T. A genetic mechanism for emergence of races in Fusarium oxysporum f. sp. lycopersici: Inactivation of avirulence gene AVR1 by transposon insertion. PLoS ONE 2012, 7, e44101. [Google Scholar]
- Armenta-López, S.E.; Valenzuela-Solano, C.; Hernández-Martínez, R. Identification and molecular analysis of races of Fusarium oxysporum f. sp. lycopersici isolated from tomato in Baja California, Mexico. Mex. J. Phytopathol. 2021, 39, 266–288. [Google Scholar] [CrossRef]
- Haegi, A.; De Felice, S.; Scotton, M.; Luongo, L.; Belisario, A. Fusarium oxysporum f. sp. melonis-melon interaction: Effect of grafting combination on pathogen gene expression. Eur. J. Plant Pathol. 2017, 149, 787–796. [Google Scholar] [CrossRef]
- O’Donnell, K.; Al-Hatmi, A.M.; Aoki, T.; Brankovics, B.; Cano-Lira, J.F.; Coleman, J.J.; de Hoog, G.S.; Di Pietro, A.; Frandsen, R.J.; Geiser, D.M. No to Neocosmospora: Phylogenomic and practical reasons for continued inclusion of the Fusarium solani species complex in the genus Fusarium. mSphere 2020, 5, 00810–00820. [Google Scholar] [CrossRef]
- Ulaszewski, B.; Sandoval-Denis, M.; Groenewald, J.Z.; Costa, M.M.; Mishra, B.; Ploch, S.; Crous, P.W.; Thines, M. Genomic features and evolution of lifestyles support the recognition of distinct genera among fusarioid fungi. Mycol. Prog. 2025, 24, 20. [Google Scholar] [CrossRef]
- van Dam, P.; Fokkens, L.; Schmidt, S.M.; Linmans, J.H.; Kistler, H.C.; Ma, L.J.; Rep, M. Effector profiles distinguish formae speciales of Fusarium oxysporum. Environ. Microbiol. 2016, 18, 4087–4102. [Google Scholar] [CrossRef]
- Li, Y.; Steenwyk, J.L.; Chang, Y.; Wang, Y.; James, T.Y.; Stajich, J.E.; Spatafora, J.W.; Groenewald, M.; Dunn, C.W.; Hittinger, C.T. A genome-scale phylogeny of the kingdom Fungi. Curr. Biol. 2021, 31, 1653–1665.e1655. [Google Scholar] [CrossRef]
- Stajich, J.E. Fungal genomes and insights into the evolution of the kingdom. Microbiol. Spectr. 2017, 5, 10–1128. [Google Scholar] [CrossRef]
- O’Donnell, K.; Ward, T.J.; Aberra, D.; Kistler, H.C.; Aoki, T.; Orwig, N.; Kimura, M.; Bjørnstad, Å.; Klemsdal, S.S. Multilocus genotyping and molecular phylogenetics resolve a novel head blight pathogen within the Fusarium graminearum species complex from Ethiopia. Fungal Genet. Biol. 2008, 45, 1514–1522. [Google Scholar] [CrossRef]
- Jiao, J.; Zhong, S.; Zhao, L.; Yang, X.; Tang, G.; Li, P. Genome-wide characterization of effector proteins in Fusarium zanthoxyli and their effects on plant’s innate immunity responses. BMC Plant Biol. 2025, 25, 298. [Google Scholar] [CrossRef]
- Zhou, X.; O’Donnell, K.; Aoki, T.; Smith, J.A.; Kasson, M.T.; Cao, Z.-M. Two novel Fusarium species that cause canker disease of prickly ash (Zanthoxylum bungeanum) in northern China form a novel clade with Fusarium torreyae. Mycologia 2016, 108, 668–681. [Google Scholar] [CrossRef]
- Zhou, X.; O’Donnell, K.; Kim, H.S.; Proctor, R.H.; Doehring, G.; Cao, Z.M. Heterothallic sexual reproduction in three canker-inducing tree pathogens within the Fusarium torreyae species complex. Mycologia 2018, 110, 710–725. [Google Scholar] [CrossRef]
- Ruan, Z.; Jiao, J.; Zhao, J.; Liu, J.; Liang, C.; Yang, X.; Sun, Y.; Tang, G.; Li, P. Genome sequencing and comparative genomics reveal insights into pathogenicity and evolution of Fusarium zanthoxyli, the causal agent of stem canker in prickly ash. BMC Genom. 2024, 25, 502. [Google Scholar] [CrossRef]
- Dita, M.; Barquero, M.; Heck, D.; Mizubuti, E.S.; Staver, C.P. Fusarium wilt of banana: Current knowledge on epidemiology and research needs toward sustainable disease management. Front. Plant Sci. 2018, 9, 1468. [Google Scholar] [CrossRef]
- Ordóñez, N.; Seidl, M.F.; Waalwijk, C.; Drenth, A.; Kilian, A.; Thomma, B.P.H.J.; Ploetz, R.C.; Kema, G.H.J. Worse comes to worst: Bananas and Panama disease—When plant and pathogen clones meet. PLoS Pathog. 2015, 11, e1005197. [Google Scholar] [CrossRef]
- Liu, S.; Li, J.; Zhang, Y.; Liu, N.; Viljoen, A.; Mostert, D.; Zuo, C.; Hu, C.; Bi, F.; Gao, H.; et al. Fusaric acid instigates the invasion of banana by Fusarium oxysporum f. sp. cubense TR4. New Phytol. 2020, 225, 913–929. [Google Scholar] [CrossRef]
- Afzalinia, S.; Mehrabi-Koushki, M.; Farokhinejad, R. Two new species of Fusarium in the F. incarnatum-equiseti species complex from Oryza sativa in Iran. Antonie Van Leeuwenhoek 2024, 118, 5. [Google Scholar] [CrossRef]
- Matić, S.; Tabone, G.; Guarnaccia, V.; Gullino, M.L.; Garibaldi, A. Emerging leafy vegetable crop diseases caused by the Fusarium incarnatum-equiseti species complex. Phytopathol. Mediterr 2020, 59, 2. [Google Scholar]
- Wang, M.; Chen, Q.; Diao, Y.; Duan, W.; Cai, L. Fusarium incarnatum-equiseti complex from China. Persoonia-Mol. Phylogeny Evol. Fungi 2019, 43, 70–89. [Google Scholar] [CrossRef]
- Avila, C.F.; Moreira, G.M.; Nicolli, C.P.; Gomes, L.B.; Abreu, L.M.; Pfenning, L.H.; Haidukowski, M.; Moretti, A.; Logrieco, A.; Del Ponte, E.M. Fusarium incarnatum-equiseti species complex associated with Brazilian rice: Phylogeny, morphology and toxigenic potential. Int. J. Food Microbiol. 2019, 306, 108267. [Google Scholar] [CrossRef]
- Moreira, G.M.; Nicolli, C.P.; Gomes, L.B.; Ogoshi, C.; Scheuermann, K.K.; Silva-Lobo, V.L.; Schurt, D.A.; Ritieni, A.; Moretti, A.; Pfenning, L.H.; et al. Nationwide survey reveals high diversity of Fusarium species and related mycotoxins in Brazilian rice: 2014 and 2015 harvests. Food Control 2020, 113, 107171. [Google Scholar] [CrossRef]
- Lu, Y.; Qiu, J.; Wang, S.; Xu, J.; Ma, G.; Shi, J.; Bao, Z. Species diversity and toxigenic potential of Fusarium incarnatum-equiseti species complex isolates from rice and soybean in China. Plant Dis. 2021, 105, 2628–2636. [Google Scholar] [CrossRef]
- Petrović, K.; Orzali, L.; Krsmanović, S.; Valente, M.T.; Tolimir, M.; Pavlov, J.; Riccioni, L. Genetic diversity and pathogenicity of the Fusarium species complex on soybean in Serbia. Plant Dis. 2024, 108, 1851–1860. [Google Scholar] [CrossRef]
- Liu, Y.; Tian, Y.; Zhao, X.; Yue, L.; Uwaremwe, C.; Zhou, Q.; Wang, Y.; Zhang, Y.; Dun, Z.; Cui, Z.; et al. Identification of Pathogenic Fusarium spp. Responsible for Root Rot of Angelica sinensis and Characterization of Their Biological Enemies in Dingxi, China. Plant Dis. 2022, 106, 1898–1910. [Google Scholar] [CrossRef]
- Niyongabo Turatsinze, A.; Xie, X.; Ye, A.; Chen, G.; Wang, Y.; Yue, L.; Zhou, Q.; Wu, L.; Zhang, M.; Zhang, Z. Fusarium cross-infection in medicinal herbs alters rhizosphere microbiomes and disrupts mycorrhizal functions under soil physicochemical imbalances. Plant Soil 2025, 1–31. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, R.; Sha, Y. Distribution, pathogenicity and disease control of Fusarium tricinctum. Front. Microbiol 2022, 13, 939927. [Google Scholar] [CrossRef]
- Senatore, M.; Ward, T.; Cappelletti, E.; Beccari, G.; McCormick, S.; Busman, M.; Laraba, I.; O’Donnell, K.; Prodi, A. Species diversity and mycotoxin production by members of the Fusarium tricinctum species complex associated with Fusarium head blight of wheat and barley in Italy. Int. J. Food Microbiol. 2021, 358, 109298. [Google Scholar] [CrossRef]
- Niyongabo Turatsinze, A.; Xie, X.; Chen, G.; Ye, A.; Yue, L.; Wang, Y.; Zhou, Q.; Zhang, M.; Zhang, Z.; Zhao, J.; et al. First Report of Fusarium avenaceum Causing Root Rot of Raspberry (Rubus corchorifolius) in China. Plant Dis. 2024, 108, 3194. [Google Scholar] [CrossRef]
- Okello, P.N.; Mathew, F.M. Cross-pathogenicity studies show South Dakota isolates of Fusarium acuminatum, F. equiseti, F. graminearum, F. oxysporum, F. proliferatum, F. solani, and F. subglutinans from either soybean or corn are pathogenic to both crops. Plant Health Prog. 2019, 20, 44–49. [Google Scholar] [CrossRef]
- Bugingo, C.; Brelsford, M.; Fonseka, D.; Pasche, J.; Burrows, M. Unveiling the diversity and virulence of seedborne Fusarium species in lentil production: Insights from a two-year study in the Northern Great Plains. Plant Health Progress. 2025, 26, 146–154. [Google Scholar] [CrossRef]
- Jambhulkar, P.P.; Bajpai, R.; Reddy, H.J.; Tripathy, P.S.; Varun, P.; Rout, A.K.; Behera, B.K.; Lakshman, D.K.; Nanjundappa, M. Assessment of Genetic Diversity and the Population Structure of Species from the Fusarium fujikuroi Species Complex Causing Fusarium Stalk Rot of Maize. J. Fungi 2024, 10, 574. [Google Scholar] [CrossRef]
- Dewing, C. Comparative Genomics Reveal Processes Implicated in Host-Specificity in Species Within the American Clade of the Fusarium Fujikuroi Species Complex. Master’s Thesis, University of Pretoria (South Africa), Pretoria, South Africa, 2020. Available online: https://www.proquest.com/dissertations-theses/comparative-genomics-reveal-processes-implicated/docview/3122660831/se-2 (accessed on 15 May 2025).
- Witte, T.E.; Villeneuve, N.; Boddy, C.N.; Overy, D.P. Accessory Chromosome-Acquired Secondary Metabolism in Plant Pathogenic Fungi: The Evolution of Biotrophs Into Host-Specific Pathogens. Front. Microbiol. 2021, 12, 664276. [Google Scholar] [CrossRef]
- Li, X.; Xu, S.; Zhang, J.; Li, M. Assembly and annotation of whole-genome sequence of Fusarium equiseti. Genomics 2021, 113, 2870–2876. [Google Scholar] [CrossRef]
- Xie, S.-Y.; Ma, T.; Zhao, N.; Zhang, X.; Fang, B.; Huang, L. Whole-genome-sequencing and comparative genome analysis of Fusarium solani-melongenae causing Fusarium root and stem rot in sweetpotatoes. Microbiol. Spectr. 2022, 10, e00683-22. [Google Scholar] [CrossRef]
- Hoogendoorn, K.; Barra, L.; Waalwijk, C.; Dickschat, J.S.; Van der Lee, T.A.; Medema, M.H. Evolution and diversity of biosynthetic gene clusters in Fusarium. Front. Microbiol. 2018, 9, 1158. [Google Scholar] [CrossRef]
- Fokkens, L.; Shahi, S.; Connolly, L.R.; Stam, R.; Schmidt, S.M.; Smith, K.M.; Freitag, M.; Rep, M. The multi-speed genome of Fusarium oxysporum reveals association of histone modifications with sequence divergence and footprints of past horizontal chromosome transfer events. BioRxiv 2018, 465070. [Google Scholar] [CrossRef]
- Bates, H.J.; Pike, J.; Price, R.J.; Jenkins, S.; Connell, J.; Legg, A.; Armitage, A.; Harrison, R.J.; Clarkson, J.P. Comparative genomics and transcriptomics reveal differences in effector complement and expression between races of Fusarium oxysporum f.sp. lactucae. Front. Plant Sci. 2024, 15, 1415534. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Carnegie, A.J.; Hardy, G.; Smith, D.; Summerell, B.A.; Cano-Lira, J.F.; Guarro, J.; Houbraken, J. Fungal Planet description sheets: 625–715. Persoonia Mol. Phylogeny Evol. Fungi 2017, 39, 270. [Google Scholar] [CrossRef]
- Navale, V.D.; Sawant, A.M.; Gowda, V.U.; Vamkudoth, K.R. Assembly, annotation, and comparative whole genome sequence of Fusarium verticillioides isolated from stored maize grains. Pathogens 2022, 11, 810. [Google Scholar] [CrossRef]
- Urbaniak, M.; Waśkiewicz, A.; Trzebny, A.; Koczyk, G.; Stępień, Ł. Cyclodepsipeptide biosynthesis in hypocreales fungi and sequence divergence of the non-ribosomal peptide synthase genes. Pathogens 2020, 9, 552. [Google Scholar] [CrossRef]
- Navasca, A.; Singh, J.; Rivera-Varas, V.; Gill, U.; Secor, G.; Baldwin, T. Dispensable genome and segmental duplications drive the genome plasticity in Fusarium solani. Front. Fungal Biol. 2025, 6, 1432339. [Google Scholar] [CrossRef]
- van Westerhoven, A.C.; Aguilera-Galvez, C.; Nakasato-Tagami, G.; Shi-Kunne, X.; Martinez de la Parte, E.; Chavarro-Carrero, E.; Meijer, H.J.; Feurtey, A.; Maryani, N.; Ordóñez, N. Segmental duplications drive the evolution of accessory regions in a major crop pathogen. New Phytol. 2024, 242, 610–625. [Google Scholar] [CrossRef]
- Ma, L.J. Horizontal chromosome transfer and rational strategies to manage Fusarium vascular wilt diseases. Mol. Plant Pathol. 2014, 15, 763. [Google Scholar] [CrossRef]
- Yang, H.; Yu, H.; Ma, L.-J. Accessory chromosomes in Fusarium oxysporum. Phytopathology 2020, 110, 1488–1496. [Google Scholar] [CrossRef]
- Wiemann, P.; Sieber, C.M.; Von Bargen, K.W.; Studt, L.; Niehaus, E.M.; Espino, J.J.; Huss, K.; Michielse, C.B.; Albermann, S.; Wagner, D.; et al. Deciphering the cryptic genome: Genome-wide analyses of the rice pathogen Fusarium fujikuroi reveal complex regulation of secondary metabolism and novel metabolites. PLoS Pathog. 2013, 9, e1003475. [Google Scholar] [CrossRef]
- Rampersad, S.N. Pathogenomics and Management of Fusarium Diseases in Plants. Pathogens 2020, 9, 340. [Google Scholar] [CrossRef]
- Thatcher, L.F.; Gardiner, D.M.; Kazan, K.; Manners, J.M. A highly conserved effector in Fusarium oxysporum is required for full virulence on Arabidopsis. Mol. Plant-Microbe Interact. MPMI 2012, 25, 180–190. [Google Scholar] [CrossRef]
- Henry, P.M.; Pincot, D.D.A.; Jenner, B.N.; Borrero, C.; Aviles, M.; Nam, M.-H.; Epstein, L.; Knapp, S.J.; Gordon, T.R. Horizontal chromosome transfer and independent evolution drive diversification in Fusarium oxysporum f. sp. fragariae. New Phytol. 2021, 230, 327–340. [Google Scholar] [CrossRef]
- Epstein, L.; Kaur, S.; Henry, P.M. The emergence of Fusarium oxysporum f. sp. apii race 4 and Fusarium oxysporum f. sp. coriandrii highlights major obstacles facing agricultural production in coastal California in a warming climate: A case study. Front. Plant Sci. 2022, 13, 921516. [Google Scholar] [CrossRef]
- Ma, H.; Liu, Y.; Zhao, X.; Zhang, S.; Ma, H. Exploring and applying genes to enhance the resistance to Fusarium head blight in wheat. Front. Plant Sci. 2022, 13, 1026611. [Google Scholar] [CrossRef]
- Niehaus, E.-M.; Münsterkötter, M.; Proctor, R.H.; Brown, D.W.; Sharon, A.; Idan, Y.; Oren-Young, L.; Sieber, C.M.; Novák, O.; Pěnčík, A.; et al. Comparative “omics” of the Fusarium fujikuroi species complex highlights differences in genetic potential and metabolite synthesis. Genome Biol. Evol. 2016, 8, 3574–3599. [Google Scholar] [CrossRef]
- Armitage, A.D.; Taylor, A.; Sobczyk, M.K.; Baxter, L.; Greenfield, B.P.J.; Bates, H.J.; Wilson, F.; Jackson, A.C.; Ott, S.; Harrison, R.J.; et al. Characterisation of pathogen-specific regions and novel effector candidates in Fusarium oxysporum f. sp. cepae. Sci. Rep. 2018, 8, 13530. [Google Scholar] [CrossRef]
- Lanubile, A.; Ferrarini, A.; Maschietto, V.; Delledonne, M.; Marocco, A.; Bellin, D. Functional genomic analysis of constitutive and inducible defense responses to Fusarium verticillioides infection in maize genotypes with contrasting ear rot resistance. BMC Genom. 2014, 15, 710. [Google Scholar] [CrossRef]
- Lightfoot, J.D.; Fuller, K.K. CRISPR/Cas9-Mediated Gene Replacement in the Fungal Keratitis Pathogen Fusarium solani var. petroliphilum. Microorganisms 2019, 7, 457. [Google Scholar] [CrossRef]
- Pokhrel, A.; Seo, S.; Wang, Q.; Coleman, J.J. Targeted Gene Disruption via CRISPR/Cas9 Ribonucleoprotein Complexes in Fusarium oxysporum. In Fusarium Wilt; Coleman, J.J., Ed.; Methods in Molecular Biology; Humana: New York, NY, USA, 2022; Volume 2391, pp. 75–87. [Google Scholar] [CrossRef]
- van Dijk, A.; Wilson, A.M.; Marx, B.; Hough, B.; Swalarsk-Parry, B.; De Vos, L.; Wingfield, M.J.; Wingfield, B.D.; Steenkamp, E.T. CRISPR-Cas9 genome editing reveals that the Pgs gene of Fusarium circinatum is involved in pathogenicity, growth and sporulation. Fungal Genet. Biol. 2025, 177, 103970. [Google Scholar] [CrossRef]
- de Vega-Bartol, J.J.; Martín-Dominguez, R.; Ramos, B.; García-Sánchez, M.A.; Díaz-Mínguez, J.M. New virulence groups in Fusarium oxysporum f. sp. phaseoli: The expression of the gene coding for the transcription factor ftf1 correlates with virulence. Phytopathology 2011, 101, 470–479. [Google Scholar] [CrossRef]
- Niño-Sánchez, J.; Casado-Del Castillo, V.; Tello, V.; De Vega-Bartol, J.J.; Ramos, B.; Sukno, S.A.; Díaz Mínguez, J.M. The FTF gene family regulates virulence and expression of SIX effectors in Fusarium oxysporum. Mol. Plant Pathol. 2016, 17, 1124–1139. [Google Scholar] [CrossRef]
- Hao, G.; Proctor, R.H.; Brown, D.W.; Rhoades, N.A.; Naumann, T.A.; Kim, H.; Gutiėrrez, S.; McCormick, S.P. TRI14 Is Critical for Fusarium graminearum Infection and Spread in Wheat. Appl. Microbiol. 2024, 4, 839–855. [Google Scholar] [CrossRef]
- Rep, M.; van der Does, H.C.; Meijer, M.; van Wijk, R.; Houterman, P.M.; Dekker, H.L.; de Koster, C.G.; Cornelissen, B.J. A small, cysteine-rich protein secreted by Fusarium oxysporum during colonization of xylem vessels is required for I-3-mediated resistance in tomato. Mol. Microbiol. 2004, 53, 1373–1383. [Google Scholar] [CrossRef]
- Jenner, B.N.; Henry, P.M. Pathotypes of Fusarium oxysporum f. sp. fragariae express discrete repertoires of accessory genes and induce distinct host transcriptional responses during root infection. Environ. Microbiol. 2022, 24, 4570–4586. [Google Scholar] [CrossRef]
- Brown, N.A.; Evans, J.; Mead, A.; Hammond-Kosack, K.E. A spatial temporal analysis of the Fusarium graminearum transcriptome during symptomless and symptomatic wheat infection. Mol. Plant Pathol. 2017, 18, 1295–1312. [Google Scholar] [CrossRef]
- Pimentel, M.F.; Rocha, L.F.; Subedi, A.; Bond, J.P.; Fakhoury, A.M. Dual RNA-seq reveals transcriptome changes during Fusarium virguliforme-Trichoderma afroharzianum interactions. PLoS ONE 2025, 20, e0310850. [Google Scholar] [CrossRef]
- Seong, K.-Y.; Pasquali, M.; Zhou, X.; Song, J.; Hilburn, K.; McCormick, S.; Dong, Y.; Xu, J.-R.; Kistler, H.C. Global gene regulation by Fusarium transcription factors Tri6 and Tri10 reveals adaptations for toxin biosynthesis. Mol. Microbiol. 2009, 72, 354–367. [Google Scholar] [CrossRef]
- Nasmith, C.G.; Walkowiak, S.; Wang, L.; Leung, W.W.Y.; Gong, Y.; Johnston, A.; Harris, L.J.; Guttman, D.S.; Subramaniam, R. Tri6 is a global transcription regulator in the phytopathogen Fusarium graminearum. PLoS Pathog. 2011, 7, e1002266. [Google Scholar] [CrossRef]
- Caracuel, Z.; Roncero, M.I.; Espeso, E.A.; González-Verdejo, C.I.; García-Maceira, F.I.; Di Pietro, A. The pH signaling transcription factor PacC controls virulence in the plant pathogen Fusarium oxysporum. Mol. Microbiol. 2003, 48, 765–779. [Google Scholar] [CrossRef]
- van der Does, H.C.; Fokkens, L.; Yang, A.; Schmidt, S.M.; Langereis, L.; Lukasiewicz, J.M.; Hughes, T.R.; Rep, M. Transcription factors encoded on core and accessory chromosomes of Fusarium oxysporum induce expression of effector genes. PLoS Genet. 2016, 12, e1006401. [Google Scholar] [CrossRef]
- Michielse, C.B.; van Wijk, R.; Reijnen, L.; Manders, E.M.M.; Boas, S.; Olivain, C.; Alabouvette, C.; Rep, M. The nuclear protein Sge1 of Fusarium oxysporum is required for parasitic growth. PLoS Pathog. 2009, 5, e1000637. [Google Scholar] [CrossRef]
- Kim, H.-K.; Lee, S.; Jo, S.-M.; McCormick, S.P.; Butchko, R.A.E.; Proctor, R.H.; Yun, S.-H. Functional roles of FgLaeA in controlling secondary metabolism, sexual development, and virulence in Fusarium graminearum. PLoS ONE 2013, 8, e68441. [Google Scholar] [CrossRef]
- Studt, L.; Rösler, S.M.; Burkhardt, I.; Arndt, B.; Freitag, M.; Humpf, H.-U.; Dickschat, J.S.; Tudzynski, B. Knock-down of the methyltransferase Kmt6 relieves H3K27me3 and results in induction of cryptic and otherwise silent secondary metabolite gene clusters in Fusarium fujikuroi. Environ. Microbiol. 2016, 18, 4037–4054. [Google Scholar] [CrossRef]
- Mihlan, M.; Homann, V.; Liu, T.-W.D.; Tudzynski, B. AREA directly mediates nitrogen regulation of gibberellin biosynthesis in Gibberella fujikuroi, but its activity is not affected by NMR. Mol. Microbiol. 2003, 47, 975–991. [Google Scholar] [CrossRef]
- Bae, H.; Kim, M.S.; Sicher, R.C.; Bae, H.-J.; Bailey, B.A. Necrosis- and ethylene-inducing peptide from Fusarium oxysporum induces a complex cascade of transcripts associated with signal transduction and cell death in Arabidopsis. Plant Physiol. 2006, 141, 1056–1067. [Google Scholar] [CrossRef]
- Ravalason, H.; Grisel, S.; Chevret, D.; Favel, A.; Berrin, J.-G.; Sigoillot, J.-C.; Herpoël-Gimbert, I. Fusarium verticillioides secretome as a source of auxiliary enzymes to enhance saccharification of wheat straw. Bioresour. Technol. 2012, 114, 589–596. [Google Scholar] [CrossRef]
- Gawehns, F.; Houterman, P.M.; Ait Ichou, F.; Michielse, C.B.; Hijdra, M.; Cornelissen, B.J.C.; Rep, M.; Takken, F.L.W. The Fusarium oxysporum effector Six6 contributes to virulence and suppresses I-2-mediated cell death. Mol. Plant-Microbe Interact. 2014, 27, 336–348. [Google Scholar] [CrossRef]
- Su, Z.; Bernardo, A.; Tian, B.; Chen, H.; Wang, S.; Ma, H.; Cai, S.; Liu, D.; Zhang, D.; Li, T.; et al. A deletion mutation in TaHRC confers Fhb1 resistance to Fusarium head blight in wheat. Nat. Genet. 2019, 51, 1099–1105. [Google Scholar] [CrossRef]
- Catanzariti, A.M.; Do, H.T.T.; Bru, P.; de Sain, M.; Thatcher, L.F.; Rep, M.; Jones, D.A. The tomato I gene for Fusarium wilt resistance encodes an atypical leucine-rich repeat receptor-like protein whose function is nevertheless dependent on SOBIR1 and SERK3/BAK1. Plant J. 2017, 89, 1195–1209. [Google Scholar] [CrossRef]
- Kumar, J.M.; Dikshit, H.K.; Aski, M.; Gupta, S.; Singh, A.; Tripathi, A.; Thribhuvan, R.; Chourasia, N.K. Molecular mapping of a gene conferring Fusarium wilt resistance in lentil (Lens culinaris Medikus subsp. culinaris) using bulked-segregant analysis. Legume Res. 2023, 46, 801–805. [Google Scholar] [CrossRef]
- Bugingo, C.; Brelsford, M.; McPhee, K.; Burrows, M. Pathotype characterization of Fusarium oxysporum f. sp. lentis (Fol) isolates in North America. Plant Health Progress. 2024, 25, 56–63. [Google Scholar] [CrossRef]
- Srour, A.; Afzal, A.J.; Blahut-Beatty, L.; Hemmati, N.; Simmonds, D.H.; Li, W.; Liu, M.; Town, C.D.; Sharma, H.; Lightfoot, D.A. The receptor-like kinase at *Rhg1-a/Rfs2* caused pleiotropic resistance to sudden death syndrome and soybean cyst nematode as a transgene by altering signaling responses. BMC Genom. 2012, 13, 368. [Google Scholar] [CrossRef]
- Ma, P.; Liu, E.; Zhang, Z.; Li, T.; Zhou, Z.; Yao, W.; Chen, J.; Wu, J.; Xu, Y.; Zhang, H. Genetic variation in ZmWAX2 confers maize resistance to Fusarium verticillioides. Plant Biotechnol. J. 2023, 21, 1812–1826. [Google Scholar] [CrossRef]
- Xu, Y.; Fan, Y.; Liu, L.; Cao, J.; Zhou, J.; Liu, E.; Li, R.; Ma, P.; Yao, W.; Wu, J.; et al. Enhancing maize resistance to Fusarium verticillioides through modulation of cell wall structure and components by ZmXYXT2. J. Adv. Res. 2025. Advance online publication. [Google Scholar] [CrossRef]
- Chen, A.; Sun, J.; Martin, G.; Gray, L.-A.; Hřibová, E.; Christelová, P.; Yahiaoui, N.; Rounsley, S.; Lyons, R.; Batley, J.; et al. Identification of a Major QTL-Controlling Resistance to the Subtropical Race 4 of Fusarium oxysporum f. sp. cubense in Musa acuminata ssp. malaccensis. Pathogens 2023, 12, 289. [Google Scholar] [CrossRef]
- Wang, P.; Zhou, L.; Jamieson, P.; Zhang, L.; Zhao, Z.; Babilonia, K.; Shao, W.; Wu, L.; Mustafa, R.; Amin, I.; et al. The cotton wall-associated kinase GhWAK7A mediates responses to fungal wilt pathogens by complexing with the chitin sensory receptors. Plant Cell 2020, 32, 3978–4001. [Google Scholar] [CrossRef]
- Dong, J.; Xu, J.; Xu, X.; Xu, Q.; Chen, X. Inheritance and quantitative trait locus mapping of Fusarium wilt resistance in cucumber. Front. Plant Sci. 2019, 10, 1425. [Google Scholar] [CrossRef]
- Bartholomew, E.S.; Xu, S.; Zhang, Y.; Yin, S.; Feng, Z.; Chen, S.; Sun, L.; Yang, S.; Wang, Y.; Liu, P.; et al. A chitinase CsChi23 promoter polymorphism underlies cucumber resistance against Fusarium oxysporum f. sp. cucumerinum. New Phytol. 2022, 236, 1471–1486. [Google Scholar] [CrossRef]
- Yang, D.; Zhang, X.; Ming, Y.; Liu, C.; Zhang, X.; Liu, S.; Zhu, L. Characterization of the High-Quality Genome Sequence and Virulence Factors of Fusarium oxysporum f. sp. vasinfectum Race 7. J. Fungi 2024, 10, 242. [Google Scholar] [CrossRef]
- Zhang, Q.; Shi, X.; Gao, T.; Xing, Y.; Jin, H.; Hao, J.; Liu, X.; Liu, X.; Liu, P. Precision management of Fusarium fujikuroi in rice through seed coating with an enhanced nanopesticide using a tannic acid-ZnII formulation. J. Nanobiotechnology 2024, 22, 717. [Google Scholar] [CrossRef]
- Beccari, G.; Hao, G.; Liu, H. Editorial: Fusarium pathogenesis: Infection mechanisms and disease progression in host plants. Front. Plant Sci. 2022, 13, 1020404. [Google Scholar] [CrossRef]
- Sperschneider, J.; Gardiner, D.M.; Thatcher, L.F.; Lyons, R.; Singh, K.B.; Manners, J.M.; Taylor, J.M. Genome-Wide Analysis in Three Fusarium Pathogens Identifies Rapidly Evolving Chromosomes and Genes Associated with Pathogenicity. Genome Biol. Evol. 2015, 7, 1613–1627. [Google Scholar] [CrossRef]
- Wang, H.; Yao, G.; Chen, W.; Ayhan, D.H.; Wang, X.; Sun, J.; Yi, S.; Meng, T.; Chen, S.; Geng, X.; et al. A gap-free genome assembly of Fusarium oxysporum f. sp. conglutinans, a vascular wilt pathogen. Sci Data 2024, 11, 925. [Google Scholar] [CrossRef]
- Huang, X.-Q.; Lu, X.-H.; Sun, M.-H.; Guo, R.-J.; van Diepeningen, A.D.; Li, S.-D. Transcriptome analysis of virulence-differentiated Fusarium oxysporum f. sp. cucumerinum isolates during cucumber colonisation reveals pathogenicity profiles. BMC Genom. 2019, 20, 570. [Google Scholar] [CrossRef]
- Armstrong, G.M.; Armstrong, J.K. Formae specialesand races of Fusarium oxysporum causing wilt disease. In Fusarium: Disease, Biology, and Taxonomy; Nelson, P.E., Toussoun, T.A., Cook, R.J., Eds.; Pennsylvania State University Press: University Park, PA, USA, 1981; pp. 391–399. Available online: https://www.scirp.org/reference/referencespapers?referenceid=399958 (accessed on 15 May 2025).
- Gordon, T.R.; Martyn, R.D. The evolutionary biology of Fusarium oxysporum. Annu. Rev. Phytopathol. 1997, 35, 111–128. [Google Scholar] [CrossRef]
- Lievens, B.; Rep, M.; Thomma, B.P. Recent developments in the molecular discrimination of formae specialesof Fusarium oxysporum. Pest Manag. Sci. 2008, 64, 781–788. [Google Scholar] [CrossRef]
- O’Donnell, K.; Sutton, D.A.; Rinaldi, M.G.; Magnon, K.C.; Cox, P.A.; Revankar, S.G.; Sanche, S.; Geiser, D.M.; Juba, J.H.; Van Burik, J.A.H.; et al. Genetic diversity of human pathogenic members of the Fusarium oxysporum complex inferred from multilocus DNA sequence data and amplified fragment length polymorphism analyses: Evidence for the recent dispersion of a geographically widespread clonal lineage and nosocomial origin. J. Clin. Microbiol. 2004, 42, 5109–5120. [Google Scholar] [CrossRef]
- Summerell, B.A.; Laurence, M.H.; Liew, E.C.Y.; Leslie, J.F. Biogeography and phylogeography of Fusarium: A review. Fungal Divers. 2010, 44, 3–13. [Google Scholar] [CrossRef]
- Babilonia, K.; Wang, P.; Liu, Z.; Jamieson, P.; Mormile, B.; Rodrigues, O.; Zhang, L.; Lin, W.; Clement, C.D.; Menezes de Moura, S.; et al. A nonproteinaceous Fusarium cell wall extract triggers receptor-like protein-dependent immune responses in Arabidopsis and cotton. New Phytol. 2021, 230, 275–289. [Google Scholar] [CrossRef]
- Dong, J.; Wang, Y.; Xian, Q.; Chen, X.; Xu, J. Transcriptome analysis reveals ethylene-mediated defense responses to Fusarium oxysporum f. sp. cucumerinum infection in Cucumis sativus L. BMC Plant Biol. 2020, 20, 334. [Google Scholar] [CrossRef]
- Mierziak, J.; Wojtasik, W. Epigenetic weapons of plants against fungal pathogens. BMC Plant Biol. 2024, 24, 175. [Google Scholar] [CrossRef]
- Liu, S.; Wu, J.; Sun, Y.; Xu, Y.; Zhou, S.; Luo, P.; Wang, Z.; Chen, D.; Liang, X.; Kang, Z.; et al. A novel key virulence factor, FoSSP71, inhibits plant immunity and promotes pathogenesis in Fusarium oxysporum f. sp. cubense. Microbiol. Spectr. 2025, 13, e02940-24. [Google Scholar] [CrossRef]
- Wang, C.; Huang, Z.; Duan, Z.; Zhu, L.; Di, R.; Bao, Y.; Powell, C.A.; Hu, Q.; Chen, B.; Zhang, M.; et al. Pectate lyase from Fusarium sacchari induces plant immune responses and contributes to virulence. Microbiol. Spectr. 2023, 11, e00165-23. [Google Scholar] [CrossRef]
- Munkvold, G.P. Fusarium species and their associated mycotoxins. In Mycotoxigenic Fungi: Methods and Protocols; Moretti, A., Susca, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2021; pp. 123–139. [Google Scholar] [CrossRef]
- Bräse, S.; Encinas, A.; Keck, J.; Nising, C.F. Chemistry and biology of mycotoxins and related fungal metabolites. Chem. Rev. 2009, 109, 3903–3990. [Google Scholar] [CrossRef]
- Steyn, P.S. Mycotoxins, general view, chemistry and structure. Toxicol. Lett. 1995, 82–83, 843–851. [Google Scholar] [CrossRef]
- International Agency for Research on Cancer. Some Naturally Occurring Substances: Food Items and Constituents, Heterocyclic Aromatic Amines and Mycotoxins. In IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; World Health Organization: Geneva, Switzerland, 1993; Volume 56, Available online: https://publications.iarc.fr/56 (accessed on 15 May 2025).
- Pietragallo, Z.; Kovalsky, P.; Nagl, V.; Schwartz-Zimmermann, H.E.; Moll, W.-D.; Schatzmayr, G.; Krska, R. Mycotoxin exposure and human health risk in rural sub-Saharan Africa. Toxins 2021, 13, 618. [Google Scholar] [CrossRef]
- Alexander, N.J.; Hohn, T.M.; McCormick, S.P. The TRI11 gene of Fusarium sporotrichioides encodes a cytochrome P450 monooxygenase required for C-15 hydroxylation in trichothecene biosynthesis. Appl. Environ. Microbiol. 1998, 65, 5621–5625. [Google Scholar] [CrossRef]
- Brown, D.W.; Butchko, R.A.E.; Busman, M.; Proctor, R.H. The Fusarium verticillioides FUM gene cluster encodes a Zn(II)2Cys6 protein that affects FUM gene expression and fumonisin production. Eukaryot. Cell 2007, 6, 1210–1218. [Google Scholar] [CrossRef]
- Chen, Y.; Kistler, H.C.; Ma, Z. Fusarium graminearum trichothecene mycotoxins: Biosynthesis, regulation, and management. Annu. Rev. Phytopathol. 2019, 57, 15–39. [Google Scholar] [CrossRef]
- Wang, J.; Lin, S.; Zhu, G.; Zhao, Z.; Wang, H.; Zhou, M.; Zhao, X.; Wu, A. Identification of Candidate Genes Associated with Trichothecene Biosynthesis in Fusarium graminearum Species Complex Combined with Transcriptomic and Proteomic Analysis. Microorganisms 2022, 10, 1479. [Google Scholar] [CrossRef]
- Schaarschmidt, S.; and Fauhl-Hassek, C. Mycotoxins during the processes of nixtamalization and tortilla production. Toxins 2019, 11, 227. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. FAOSTAT Statistical Database. 2023. Available online: https://www.fao.org/faostat/ (accessed on 15 May 2025).
- Food and Agriculture Organization of the United Nations. World Food and Agriculture—Statistical Yearbook; Food & Agriculture Org: Rome, Italy, 2022; Available online: https://openknowledge.fao.org/server/api/core/bitstreams/0c372c04-8b29-4093-bba6-8674b1d237c7/content (accessed on 30 July 2025).
- Logrieco, A.; Battilani, P.; Leggieri, M.C.; Jiang, D.; Haque, M.M.; Mahmud, N.U. Mycotoxins in maize: A global concern. Toxins 2021, 13, 494. [Google Scholar] [CrossRef]
- Bottalico, A.; Perrone, G. Toxigenic Fusarium species and mycotoxins associated with head blight in small-grain cereals in Europe. Eur. J. Plant Pathol. 2002, 108, 611–624. [Google Scholar] [CrossRef]
- Lee, H.J.; Ryu, D. Worldwide occurrence of mycotoxins in cereals and cereal-derived food products: Public health perspectives of their co-occurrence. J. Agric. Food Chem. 2017, 65, 7034–7051. [Google Scholar] [CrossRef]
- Tanaka, K.; Sago, Y.; Zheng, Y.; Nakagawa, H.; Kushiro, M. Mycotoxins in rice. Int. J. Food Microbiol. 2007, 118, 59–65. [Google Scholar] [CrossRef]
- Desjardins, A.E.; Manandhar, H.K.; Plattner, R.D.; Manandhar, G.; Poling, S.M.; Maragos, C.M. Fusarium species from Nepalese rice and production of mycotoxins and gibberellic acid by selected species. Appl. Environ. Microbiol. 2000, 66, 1020–1025. [Google Scholar] [CrossRef]
- Eskola, M.; Kos, G.; Elliott, C.T.; Hajšlová, J.; Mayar, S.; Krska, R. Worldwide contamination of food-crops with mycotoxins: Validity of the widely cited ‘FAO estimate’ of 25%. Crit. Rev. Food Sci. Nutr. 2020, 60, 2773–2789. [Google Scholar] [CrossRef]
- Magan, N.; Medina, A.; Aldred, D. Possible climate-change effects on mycotoxin contamination of food crops pre- and postharvest. Plant Pathol. 2011, 60, 150–163. [Google Scholar] [CrossRef]
- Battilani, P.; Toscano, P.; Van der Fels-Klerx, H.J.; Moretti, A.; Camardo Leggieri, M.; Brera, C.; Rortais, A.; Goumperis, T.; Robinson, T. Aflatoxin B1 contamination in maize in Europe increases due to climate change. Sci. Rep. 2016, 6, 24328. [Google Scholar] [CrossRef]
- Wagacha, J.M.; Muthomi, J.W. Mycotoxin problem in Africa: Current status, implications to food safety and health and possible management strategies. Int. J. Food Microbiol. 2008, 124, 1–12. [Google Scholar] [CrossRef]
- Torres, O.; Matute, J.; Gelineau-van Waes, J.; Maddox, J.R.; Gregory, S.G.; Ashley-Koch, A.E.; Showker, J.L.; Voss, K.A.; Riley, R.T. Human dietary exposure to fumonisin B1 in a high-risk maize area of Guatemala. Food Addit. Contam. Part A 2007, 24, 429–437. [Google Scholar] [CrossRef]
- Liu, Y.; Wu, F. Global burden of aflatoxin-induced hepatocellular carcinoma: A risk assessment. Environ. Health Perspect. 2010, 118, 818–824. [Google Scholar] [CrossRef]
- Yazar, S.; Omurtag, G.Z. Fumonisins, trichothecenes and zearalenone in cereals. Int. J. Mol. Sci. 2008, 9, 2062–2090. [Google Scholar] [CrossRef]
- Medina, Á.; Rodríguez, A.; Magan, N. Effect of climate change on Aspergillus flavus and aflatoxin B1 production. Front. Microbiol. 2014, 5, 348. [Google Scholar] [CrossRef]
- Bhat, R.V.; Rai, R.N.; Karimullakhan, M.; Rao, V.S. Food Safety in Food Security and Food Trade: Mycotoxin Food Safety Risk in Developing Countries; International Food Policy Research Institute: Washington, DC, USA, 2003; Available online: https://www.ifpri.org/publication/mycotoxin-food-safety-risk-developing-countries (accessed on 15 May 2025).
- Udomkun, P.; Wiredu, A.N.; Nagle, M.; Müller, J.; Vanlauwe, B.; Bandyopadhyay, R. Mycotoxins in sub-Saharan Africa: Present situation, socio-economic impact, awareness, and outlook. Food Control 2017, 72, 110–122. [Google Scholar] [CrossRef]
- Gómez-Osorio, L.-M. Addressing the gaps in mycotoxin management: Challenges and strategies for South America’s poultry industry. MycotoxinSite 2023. Available online: https://mycotoxinsite.com/addressing-gaps-mycotoxin-management-challenges-strategies-south-americas-poultry-industry/?lang=en (accessed on 15 May 2025).
- Juan-García, A.; Jakšić, D. Novel Approaches in Mycotoxins Research: Detection, Prevention and Mode of Action. Toxins 2025, 17, 161. [Google Scholar] [CrossRef]
- Li, R.; Wen, Y.; Wang, F.; He, P. Recent advances in immunoassays and biosensors for mycotoxins detection in feedstuffs and foods. J. Anim. Sci. Biotechnol. 2021, 12, 108. [Google Scholar] [CrossRef]
- Beres, B.L.; Brûlé-Babel, A.L.; Ye, Z.; Graf, R.J.; Turkington, T.K.; Harding, M.W.; Kutcher, H.R.; Hooker, D.C. Exploring genotype × environment × management synergies to manage Fusarium head blight in wheat. Can. J. Plant Pathol. 2018, 40, 179–188. [Google Scholar] [CrossRef]
- Marburger, D.A.; Venkateshwaran, M.; Conley, S.P.; Esker, P.D.; Lauer, J.G.; Ané, J.M. Crop rotation and management effect on Fusarium spp. populations. Crop Sci. 2015, 55, 365–376. [Google Scholar] [CrossRef]
- Rojas, E.C.; Jørgensen, H.J.; Jensen, B.; Collinge, D.B. Fusarium diseases: Biology and management perspectives. In Integrated Disease Management of Wheat and Barley; Burleigh Dodds Science Publishing: Cambridge, UK, 2018; pp. 43–66. [Google Scholar]
- University of Wisconsin-Madison. What do we know about crop rotations? Wis. Corn Agron. 2002, 9, 84–85. Available online: https://corn.agronomy.wisc.edu/WCM/W110.aspx (accessed on 6 May 2025).
- Leslie, J.F.; Pearson, C.A.; Nelson, P.E.; Toussoun, T. Fusarium spp. from corn, sorghum, and soybean fields in the central and eastern United States. Ecol. Stud. 1990, 44, 66. [Google Scholar]
- Nelson, P.E.; Toussoun, T.A.; Marasas, W.F.O. Fusarium Species: An Illustrated Manual for Identification; Pennsylvania State University Press: University Park, PA, USA, 1983. [Google Scholar]
- Islam, M.N.; Banik, M.; Sura, S.; Tucker, J.R.; Wang, X. Implications of crop rotation and fungicide on Fusarium and mycotoxin spectra in Manitoba barley, 2017–2019. Toxins 2022, 14, 463. [Google Scholar] [CrossRef]
- Islam, T. An integrated pest management program for managing Fusarium head blight disease in cereals. J. Integr. Agric. 2022, 21, 3434–3444. [Google Scholar] [CrossRef]
- Krupinsky, J.M.; Bailey, K.L.; McMullen, M.P.; Gossen, B.D.; Turkington, T.K. Managing plant disease risk in diversified cropping systems. Agron. J. 2002, 94, 198–209. [Google Scholar] [CrossRef]
- Arias, M.D.; Munkvold, G.P.; Ellis, M.L.; Leandro, L.F. Distribution and frequency of Fusarium species associated with soybean roots in Iowa. Plant Dis. 2013, 97, 1557–1562. [Google Scholar] [CrossRef]
- Broders, K.D.; Lipps, P.E.; Paul, P.A.; Dorrance, A.E. Evaluation of Fusarium graminearum associated with corn and soybean seed and seedling disease in Ohio. Plant Dis. 2007, 91, 1155–1160. [Google Scholar] [CrossRef]
- Okello, P.N.; Petrović, K.; Kontz, B.; Ali, S.; Marek, L.F.; Mathew, F.M. Root rot caused by species of Fusarium on Brassica carinata in South Dakota. Plant Health Prog. 2018, 19, 188–192. [Google Scholar] [CrossRef]
- Okello, P.N.; Petrovic, K.; Singh, A.K.; Kontz, B.; Mathew, F.M. Characterization of species of Fusarium causing root rot of soybean (Glycine max L.) in South Dakota, USA. Can. J. Plant Pathol. 2020, 42, 560–571. [Google Scholar] [CrossRef]
- Pioli, R.N.; Mozzoni, L.; Morandi, E.N. First report of pathogenic association between Fusarium graminearum and soybean. Plant Dis. 2004, 88, 220. [Google Scholar] [CrossRef]
- Xue, A.G.; Cober, E.; Voldeng, H.D.; Babcock, C.; Clear, R.M. Evaluation of the pathogenicity of Fusarium graminearum and Fusarium pseudograminearum on soybean seedlings under controlled conditions. Can. J. Plant Pathol. 2007, 29, 35–40. [Google Scholar] [CrossRef]
- Xu, F.; Liu, W.; Song, Y.; Zhou, Y.; Xu, X.; Yang, G.; Wang, J.; Zhang, J.; Liu, L. The distribution of Fusarium graminearum and Fusarium asiaticum causing Fusarium head blight of wheat in relation to climate and cropping system. Plant Dis. 2021, 105, 2830–2835. [Google Scholar] [CrossRef]
- Cotten, T.K.; Munkvold, G.P. Survival of Fusarium moniliforme, F. proliferatum, and F. subglutinans in maize stalk residue. Phytopathology 1998, 88, 550–555. [Google Scholar] [CrossRef]
- Inch, S.A.; Gilbert, J. Survival of Gibberella zeae in Fusarium-damaged wheat kernels. Plant Dis. 2003, 87, 282–287. [Google Scholar] [CrossRef]
- Paulitz, T.C.; Smiley, R.W.; Cook, R.J. Insights into the prevalence and management of soilborne cereal pathogens under direct seeding in the Pacific Northwest, USA. Can. J. Plant Pathol. 2002, 24, 416–428. [Google Scholar] [CrossRef]
- Bernhoft, A.; Torp, M.; Clasen, P.E.; Løes, A.K.; Kristoffersen, A.B. Influence of agronomic and climatic factors on Fusarium infestation and mycotoxin contamination of cereals in Norway. Food Addit. Contam. Part A 2012, 29, 1129–1140. [Google Scholar] [CrossRef]
- Panth, M.; Hassler, S.C.; Baysal-Gurel, F. Methods for management of soilborne diseases in crop production. Agriculture 2020, 10, 16. [Google Scholar] [CrossRef]
- Bugingo, C.; Ali, S.; Yabwalo, D.; Byamukama, E. Optimizing fungicide seed treatments for early foliar disease management in wheat under Northern Great Plains conditions. Agronomy 2025, 15, 291. [Google Scholar] [CrossRef]
- Lamichhane, J.R.; You, M.P.; Laudinot, V.; Barbetti, M.J.; Aubertot, J.N. Revisiting sustainability of fungicide seed treatments for field crops. Plant Dis. 2020, 104, 610–623. [Google Scholar] [CrossRef]
- Munkvold, G.P.; O’Mara, J.K. Laboratory and growth chamber evaluation of fungicidal seed treatments for maize seedling blight caused by Fusarium species. Plant Dis. 2002, 86, 143–150. [Google Scholar] [CrossRef]
- Cromey, M.G.; Lauren, D.R.; Parkes, R.A.; Sinclair, K.I.; Shorter, S.C.; Wallace, A.R. Control of Fusarium head blight of wheat with fungicides. Australas. Plant Pathol. 2001, 30, 301–308. [Google Scholar] [CrossRef]
- Hollingsworth, C.R.; Motteberg, C.D.; Assistant, R.; Thompson, W.G. Assessing fungicide efficacies for the management of Fusarium head blight on spring wheat and barley. Plant Health Prog. 2006, 7, 14. [Google Scholar] [CrossRef]
- Ioos, R.; Belhadj, A.; Menez, M.; Faure, A. The effects of fungicides on Fusarium spp. and Microdochium nivale and their associated trichothecene mycotoxins in French naturally-infected cereal grains. Crop Prot. 2005, 24, 894–902. [Google Scholar] [CrossRef]
- Jones, R.K. Assessments of Fusarium head blight of wheat and barley in response to fungicide treatment. Plant Dis. 2000, 84, 1021–1030. [Google Scholar] [CrossRef]
- Mesterhazy, A.; Bartok, T.; Lamper, C. Influence of wheat cultivar, species of Fusarium, and isolate aggressiveness on the efficacy of fungicides for control of Fusarium head blight. Plant Dis. 2003, 87, 1107–1115. [Google Scholar] [CrossRef]
- Wegulo, S.N.; Bockus, W.W.; Nopsa, J.H.; De Wolf, E.D.; Eskridge, K.M.; Peiris, K.H.; Dowell, F.E. Effects of integrating cultivar resistance and fungicide application on Fusarium head blight and deoxynivalenol in winter wheat. Plant Dis. 2011, 95, 554–560. [Google Scholar] [CrossRef]
- Willyerd, K.T.; Li, C.; Madden, L.V.; Bradley, C.A.; Bergstrom, G.C.; Sweets, L.E.; McMullen, M.; Ransom, J.K.; Grybauskas, A.; Osborne, L.; et al. Efficacy and stability of integrating fungicide and cultivar resistance to manage Fusarium head blight and deoxynivalenol in wheat. Plant Dis. 2012, 96, 957–967. [Google Scholar] [CrossRef]
- Kandel, Y.R.; Bradley, C.A.; Chilvers, M.I.; Mathew, F.M.; Tenuta, A.U.; Smith, D.L.; Wise, K.A.; Mueller, D.S. Effect of seed treatment and foliar crop protection products on sudden death syndrome and yield of soybean. Plant Dis. 2019, 103, 1712–1720. [Google Scholar] [CrossRef]
- Bugingo, C.; Brelsford, M.; Burrows, M. Fungicide sensitivity of Fusarium oxysporum f. sp. lentis and Fusarium acuminatum affecting lentil in the Northern Great Plains. Plant Dis. 2024, 108, 286–290. [Google Scholar]
- Ellis, M.L.; Broders, K.D.; Paul, P.A.; Dorrance, A.E. Infection of soybean seed by Fusarium graminearum and effect of seed treatments on disease under controlled conditions. Plant Dis. 2011, 95, 401–407. [Google Scholar] [CrossRef]
- Capo, L.; Zappino, A.; Reyneri, A.; Blandino, M. Role of the fungicide seed dressing in controlling seed-borne Fusarium spp. infection and in enhancing the early development and grain yield of maize. Agronomy 2020, 10, 784. [Google Scholar] [CrossRef]
- Edwards, S.G. Pydiflumetofen co-formulated with prothioconazole: A novel fungicide for Fusarium head blight and deoxynivalenol control. Toxins 2022, 14, 34. [Google Scholar] [CrossRef]
- Hou, Y.; Guo, Y.; Wang, L.; He, S.; Zheng, W.; Liu, S.; Xu, J. Impact of phenamacril on the growth and development of Fusarium pseudograminearum and control of crown rot of wheat. Plant Dis. 2023, 107, 3843–3850. [Google Scholar] [CrossRef]
- Jayawardana, M.A.; Fernando, W.G. The mechanisms of developing fungicide resistance in Fusarium graminearum causing Fusarium head blight and fungicide resistance management. Pathogens 2024, 13, 1012. [Google Scholar] [CrossRef]
- Ghosal, D.; Tarafder, M.; Datta, B. Fusarium oxysporum isolates showed varied benzimidazole fungicides sensitivity under low temperature and heavy metal stress. Rhizosphere 2020, 15, 100227. [Google Scholar] [CrossRef]
- Miao, J.; Li, Y.; Hu, S.; Li, G.; Gao, X.; Dai, T.; Liu, X. Resistance risk, resistance mechanism and the effect on DON production of a new SDHI fungicide cyclobutrifluram in Fusarium graminearum. Pestic. Biochem. Physiol. 2024, 199, 105795. [Google Scholar] [CrossRef]
- Ceresini, P.C.; Silva, T.C.; Vicentini, S.N.; Leite Júnior, R.P.; Moreira, S.I.; Castro-Ríos, K.; Garcés-Fiallos, F.R.; Krug, L.D.; Moura, S.S.; Silva, A.G.; et al. Strategies for managing fungicide resistance in the Brazilian tropical agroecosystem: Safeguarding food safety, health, and the environmental quality. Trop. Plant Pathol. 2024, 49, 36–70. [Google Scholar] [CrossRef]
- Lucas, J.A.; Hawkins, N.J.; Fraaije, B.A. The evolution of fungicide resistance. Adv. Appl. Microbiol. 2015, 90, 29–92. [Google Scholar] [CrossRef]
- Naqvi, S.A.; Farhan, M.; Ahmad, M.; Kiran, R.; Shahbaz, M.; Abbas, A.; Hakim, F.; Shabbir, M.; Tan, Y.S.; Sathiya Seelan, J.S. Fungicide resistance in Fusarium species: Exploring environmental impacts and sustainable management strategies. Arch. Microbiol. 2025, 207, 31. [Google Scholar] [CrossRef]
- Shang, N.; Yang, Y.; Xiao, Y.; Wu, Y.; Li, K.; Jiang, X.; Sanganyado, E.; Zhang, Q.; Xia, X. Exposure levels and health implications of fungicides, neonicotinoid insecticides, triazine herbicides and their associated metabolites in pregnant women and men. Environ. Pollut. 2024, 342, 123069. [Google Scholar] [CrossRef]
- Bartlett, D.W.; Clough, J.M.; Godwin, J.R.; Hall, A.A.; Hamer, M.; Parr-Dobrzanski, B. The strobilurin fungicides. Pest Manag. Sci. 2002, 58, 649–662. [Google Scholar] [CrossRef]
- Degani, O.; Kalman, B. Assessment of commercial fungicides against onion (Allium cepa) basal rot disease caused by Fusarium oxysporum f. sp. cepae and Fusarium acutatum. J. Fungi 2021, 7, 235. [Google Scholar] [CrossRef]
- Ramirez, M.L.; Chulze, S.; Magan, N. Impact of environmental factors and fungicides on growth and deoxynivalenol production by Fusarium graminearum isolates from Argentinian wheat. Crop Prot. 2004, 23, 117–125. [Google Scholar] [CrossRef]
- Dahal, N.; Shrestha, R.K. Evaluation of efficacy of fungicides against Fusarium oxysporum f. sp. lentis in vitro at Lamjung, Nepal. J. Inst. Agric. Anim. Sci. 2018, 35, 105–112. [Google Scholar] [CrossRef]
- Lal, D.K.; Wagan, K.H.; Mushatque, S.; Ehsan, S.; Chaudhary, N. Investigations on seed-borne fungi associated with maize varieties and their control. J. Microbiol. Sci. 2025, 4, 1–2. [Google Scholar] [CrossRef]
- Rajput, A.Q.; Arain, M.H.; Pathan, M.A.; Jiskani, M.M.; Lodhi, A.M. Efficacy of different fungicides against Fusarium wilt of cotton caused by Fusarium oxysporum f. sp. vasinfectum. Pak. J. Bot. 2006, 38, 875. [Google Scholar]
- Xu, C.; Guo, M.; Han, X.; Ren, C.; Liu, C.; Fu, W.; Qi, J.; Ge, Z.; Ma, Z.; Chen, Y. Fungal pathogen diversity and fungicide resistance assessment in Fusarium crown rot of wheat in the Huanghuai region of China. J. Agric. Food Chem. 2025, 73, 2299–2311. [Google Scholar] [CrossRef]
- Allen, T.W.; Enebak, S.A.; Carey, W.A. Evaluation of fungicides for control of species of Fusarium on longleaf pine seed. Crop Prot. 2004, 23, 979–982. [Google Scholar] [CrossRef]
- Hudec, K. Influence of seed treatment, temperature and origin of inocula on pathogenicity of Fusarium species to wheat and barley seedlings. Cereal Res. Commun. 2006, 34, 1059–1066. [Google Scholar] [CrossRef]
- Müllenborn, C.; Steiner, U.; Ludwig, M.; Oerke, E.C. Effect of fungicides on the complex of Fusarium species and saprophytic fungi colonizing wheat kernels. Eur. J. Plant Pathol. 2008, 120, 157–166. [Google Scholar] [CrossRef]
- Shin, J.H.; Han, J.H.; Lee, J.K.; Kim, K.S. Characterization of the maize stalk rot pathogens Fusarium subglutinans and F. temperatum and the effect of fungicides on their mycelial growth and colony formation. Plant Pathol. J. 2014, 30, 397–406. [Google Scholar] [CrossRef]
- Giedrojć, W.; Pluskota, W.E.; Wachowska, U. Fusarium graminearum in wheat—Management strategies in Central Europe. Pathogens 2025, 14, 265. [Google Scholar] [CrossRef]
- Paul, P.A.; Lipps, P.E.; Hershman, D.E.; McMullen, M.P.; Draper, M.A.; Madden, L.V. Efficacy of triazole-based fungicides for Fusarium head blight and deoxynivalenol control in wheat: A multivariate meta-analysis. Phytopathology 2008, 98, 999–1011. [Google Scholar] [CrossRef]
- Buttar, H.S.; Singh, A.; Sirari, A.; Anupam; Kaur, K.; Kumar, A.; Lal, M.K.; Tiwari, R.K.; Kumar, R. Investigating the impact of fungicides and mungbean genotypes on the management of pod rot disease caused by Fusarium equiseti and Fusarium chlamydosporum. Front. Plant Sci. 2023, 14, 1164245. [Google Scholar] [CrossRef]
- Vatankhah, M.; Saberi-Riseh, R.; Moradzadeh Eskandari, M.; Afzali, H. Evaluation of some fungicides for the control of Fusarium dry rot of potato. J. Crop Prot. 2019, 8, 275–285. [Google Scholar]
- Chala, A.; Weinert, J.; Wolf, G.A. An integrated approach to the evaluation of the efficacy of fungicides against Fusarium culmorum, the cause of head blight of wheat. J. Phytopathol. 2003, 151, 673–678. [Google Scholar] [CrossRef]
- Weems, J.D.; Haudenshield, J.S.; Bond, J.P.; Hartman, G.L.; Ames, K.A.; Bradley, C.A. Effect of fungicide seed treatments on Fusarium virguliforme infection of soybean and development of sudden death syndrome. Can. J. Plant Pathol. 2015, 37, 435–447. [Google Scholar] [CrossRef]
- Boulahouat, S.; Cherif-Silini, H.; Silini, A.; Bouket, A.C.; Luptakova, L.; Alenezi, F.N.; Belbahri, L. Biocontrol efficiency of rhizospheric Bacillus against the plant pathogen Fusarium oxysporum: A promising approach for sustainable agriculture. Microbiol. Res. 2023, 14, 892–908. [Google Scholar] [CrossRef]
- Mon, Y.Y.; Bidabadi, S.S.; Oo, K.S.; Zheng, S.J. The antagonistic mechanism of rhizosphere microbes and endophytes on the interaction between banana and Fusarium oxysporum f. sp. cubense. Physiol. Mol. Plant Pathol. 2021, 116, 101733. [Google Scholar] [CrossRef]
- Alukumbura, A.S.; Bigi, A.; Sarrocco, S.; Fernando, W.D.; Vannacci, G.; Mazzoncini, M.; Bakker, M.G. Minimal impacts on the wheat microbiome when Trichoderma gamsii T6085 is applied as a biocontrol agent to manage Fusarium head blight disease. Front. Microbiol. 2022, 13, 972016. [Google Scholar] [CrossRef]
- Liang, N.; Charron, J.B.; Jabaji, S. Comparative transcriptome analysis reveals the biocontrol mechanism of Bacillus velezensis E68 against Fusarium graminearum DAOMC 180378, the causal agent of Fusarium head blight. PLoS ONE 2023, 18, e0277983. [Google Scholar] [CrossRef]
- Yeo, Y.J.; Park, A.R.; Vuong, B.S.; Kim, J.C. Biocontrol of Fusarium head blight in rice using Bacillus velezensis JCK-7158. Front. Microbiol. 2024, 15, 1358689. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; Leeman, M.; Van Oorschot, M.M.; Van der Sluis, I.; Schippers, B.; Bakker, P.A. Dose-response relationships in biological control of Fusarium wilt of radish by Pseudomonas spp. Phytopathology 1995, 85, 1075–1080. [Google Scholar]
- Kiesewalter, H.T.; Lozano-Andrade, C.N.; Wibowo, M.; Strube, M.L.; Maróti, G.; Snyder, D.; Jørgensen, T.S.; Larsen, T.O.; Cooper, V.S.; Weber, T.; et al. Genomic and chemical diversity of Bacillus subtilis secondary metabolites against plant pathogenic fungi. mSystems 2021, 6, e00770-20. [Google Scholar] [CrossRef]
- Sivasithamparam, K.; Ghisalberti, E. Secondary metabolism in Trichoderma. In Trichoderma and Gliocladium; Taylor Francis: Oxfordshire, UK, 2002; Volume 1, pp. 139–191. [Google Scholar]
- Compant, S.; Duffy, B.; Nowak, J.; Clément, C.; Barka, E.A. Use of plant growth-promoting bacteria for biocontrol of plant diseases: Principles, mechanisms of action, and future prospects. Appl. Environ. Microbiol. 2005, 71, 4951–4959. [Google Scholar] [CrossRef]
- Petrucci, A.; Khairullina, A.; Sarrocco, S.; Jensen, D.F.; Jensen, B.; Jørgensen, H.J.; Collinge, D.B. Understanding the mechanisms underlying biological control of Fusarium diseases in cereals. Eur. J. Plant Pathol. 2023, 167, 453–476. [Google Scholar] [CrossRef]
- Gimeno, A.; Kägi, A.; Drakopoulos, D.; Bänziger, I.; Lehmann, E.; Forrer, H.R.; Keller, B.; Vogelgsang, S. From laboratory to the field: Biological control of Fusarium graminearum on infected maize crop residues. J. Appl. Microbiol. 2020, 129, 680–694. [Google Scholar] [CrossRef]
- Xue, A.G.; Chen, Y.; Voldeng, H.D.; Fedak, G.; Savard, M.E.; Längle, T.; Zhang, J.; Harman, G.E. Concentration and cultivar effects on efficacy of CLO-1 biofungicide in controlling Fusarium head blight of wheat. Biol. Control 2014, 73, 2–7. [Google Scholar] [CrossRef]
- Abaya, A.; Serajazari, M.; Hsiang, T. Control of Fusarium head blight using the endophytic fungus, Simplicillium lamellicola, and its effect on the growth of Triticum aestivum. Biol. Control 2021, 160, 104684. [Google Scholar] [CrossRef]
- Bhardwaj, M.; Kailoo, S.; Khan, R.T.; Khan, S.S.; Rasool, S. Harnessing fungal endophytes for natural management: A biocontrol perspective. Front. Microbiol. 2023, 14, 1280258. [Google Scholar] [CrossRef]
- Noel, Z.A.; Roze, L.V.; Breunig, M.; Trail, F. Endophytic fungi as a promising biocontrol agent to protect wheat from Fusarium graminearum head blight. Plant Dis. 2022, 106, 595–602. [Google Scholar] [CrossRef]
- Thangavelu, R.; Gopi, M. Field suppression of Fusarium wilt disease in banana by the combined application of native endophytic and rhizospheric bacterial isolates possessing multiple functions. Phytopathol. Mediterr. 2015, 54, 241–252. [Google Scholar] [CrossRef]
- De Lamo, F.J.; Takken, F.L. Biocontrol by Fusarium oxysporum using endophyte-mediated resistance. Front. Plant Sci. 2020, 11, 37. [Google Scholar] [CrossRef]
- Rojas, E.C.; Jensen, B.; Jørgensen, H.J.; Latz, M.A.; Esteban, P.; Ding, Y.; Collinge, D.B. Selection of fungal endophytes with biocontrol potential against Fusarium head blight in wheat. Biol. Control 2020, 144, 104222. [Google Scholar] [CrossRef]
- Bosco, S.; Prencipe, S.; Mezzalama, M.; Spadaro, D. Screening and characterization of bacterial and fungal endophytes as potential biocontrol agents for rice seed dressing against Fusarium fujikuroi. Biol. Control 2024, 196, 105580. [Google Scholar] [CrossRef]
- Nawaz, M.E.; Malik, K.; Hassan, M.N. Rice-associated antagonistic bacteria suppress the Fusarium fujikuroi causing rice bakanae disease. BioControl 2022, 67, 101–109. [Google Scholar] [CrossRef]
- O’Sullivan, C.A.; Roper, M.M.; Myers, C.A.; Thatcher, L.F. Developing actinobacterial endophytes as biocontrol products for Fusarium pseudograminearum in wheat. Front. Bioeng. Biotechnol. 2021, 9, 691770. [Google Scholar] [CrossRef]
- Abd-Elgawad, M.M.; Askary, T.H. Factors affecting success of biological agents used in controlling the plant-parasitic nematodes. Egypt. J. Biol. Pest Control 2020, 30, 17. [Google Scholar] [CrossRef]
- He, D.C.; He, M.H.; Amalin, D.M.; Liu, W.; Alvindia, D.G.; Zhan, J. Biological control of plant diseases: An evolutionary and eco-economic consideration. Pathogens 2021, 10, 1311. [Google Scholar] [CrossRef]
- Teixidó, N.; Usall, J.; Torres, R. Insight into a successful development of biocontrol agents: Production, formulation, packaging, and shelf life as key aspects. Horticulturae 2022, 8, 305. [Google Scholar] [CrossRef]
- Alisaac, E.; Mahlein, A.K. Fusarium head blight on wheat: Biology, modern detection and diagnosis and integrated disease management. Toxins 2023, 15, 192. [Google Scholar] [CrossRef]
- Figlan, S.; Mwadzingeni, L. Breeding tools for assessing and improving resistance and limiting mycotoxin production by Fusarium graminearum in wheat. Plants 2022, 11, 1933. [Google Scholar] [CrossRef]
- Poudel, B. CRISPR/Cas9 Based Gene Editing in Wheat and Potato to Enhance Multiple Disease Resistance. Ph.D. Thesis, McGill University, Montreal, QC, Canada, 2024. [Google Scholar]
- Hameed, A.; Poznanski, P.; Noman, M.; Ahmed, T.; Iqbal, A.; Nadolska-Orczyk, A.; Orczyk, W. Barley resistance to Fusarium graminearum infections: From transcriptomics to field with food safety concerns. J. Agric. Food Chem. 2022, 70, 14571–14587. [Google Scholar] [CrossRef]
- Jha, U.C.; Bohra, A.; Pandey, S.; Parida, S.K. Breeding, genetics, and genomics approaches for improving Fusarium wilt resistance in major grain legumes. Front. Genet. 2020, 11, 1001. [Google Scholar] [CrossRef]
- Yin, K.; Qiu, J.L. Genome editing for plant disease resistance: Applications and perspectives. Philos. Trans. R. Soc. B 2019, 374, 20180322. [Google Scholar] [CrossRef]
- Ollier, M.; Talle, V.; Brisset, A.L.; Le Bihan, Z.; Duerr, S.; Lemmens, M.; Goudemand, E.; Robert, O.; Hilbert, J.L.; Buerstmayr, H. QTL mapping and successful introgression of the spring wheat-derived QTL Fhb1 for Fusarium head blight resistance in three European triticale populations. Theor. Appl. Genet. 2020, 133, 457–477. [Google Scholar] [CrossRef]
- Waites, J.; Achary, V.M.; Syombua, E.D.; Hearne, S.J.; Bandyopadhyay, A. CRISPR-mediated genome editing of wheat for enhancing disease resistance. Front. Genome Ed. 2025, 7, 1542487. [Google Scholar] [CrossRef]
- Zhang, F.; Shi, Y.; Ali, J.; Xu, J.; Li, Z. Breeding by selective introgression: Theory, practices, and lessons learned from rice. Crop J. 2021, 9, 646–657. [Google Scholar] [CrossRef]
- Ma, H.; Zhang, X.; Yao, J.; Cheng, S. Breeding for the resistance to Fusarium head blight of wheat in China. Front. Agric. Sci. Eng. 2019, 6, 251–264. [Google Scholar] [CrossRef]
- Mapari, A.R.; Mehandi, S. Enhancing crop resilience: Advances and challenges in marker-assisted selection for disease resistance. J. Adv. Biol. Biotechnol. 2024, 27, 569–580. [Google Scholar] [CrossRef]
- Gaire, R.; de Arruda, M.P.; Mohammadi, M.; Brown-Guedira, G.; Kolb, F.L.; Rutkoski, J. Multi-trait genomic selection can increase selection accuracy for deoxynivalenol accumulation resulting from Fusarium head blight in wheat. Plant Genome 2022, 15, e20188. [Google Scholar] [CrossRef]
- Steiner, B.; Buerstmayr, M.; Michel, S.; Schweiger, W.; Lemmens, M.; Buerstmayr, H. Breeding strategies and advances in line selection for Fusarium head blight resistance in wheat. Trop. Plant Pathol. 2017, 42, 165–174. [Google Scholar] [CrossRef]
- Guo-Liang, J. Molecular markers and marker-assisted breeding in plants. In Plant Breeding from Laboratories to Fields; Bode, A.S., Ed.; IntechOpen: London, UK, 2013; p. Ch.3. [Google Scholar] [CrossRef]
- Luo, K.; Guo, J.; He, D.; Li, G.; Ouellet, T. Deoxynivalenol accumulation and detoxification in cereals and its potential role in wheat–Fusarium graminearum interactions. Abiotech 2023, 4, 155–171. [Google Scholar] [CrossRef]
- Jiang, F.; Doudna, J.A. CRISPR–Cas9 structures and mechanisms. Annu. Rev. Biophys. 2017, 46, 505–529. [Google Scholar] [CrossRef]
- Langner, T.; Kamoun, S.; Belhaj, K. CRISPR crops: Plant genome editing toward disease resistance. Annu. Rev. Phytopathol. 2018, 56, 479–512. [Google Scholar] [CrossRef]
- Wolter, F.; Schindele, P.; Puchta, H. Plant breeding at the speed of light: The power of CRISPR/Cas to generate directed genetic diversity at multiple sites. BMC Plant Biol. 2019, 19, 176. [Google Scholar] [CrossRef]
- Pixley, K.V.; Falck-Zepeda, J.B.; Paarlberg, R.L.; Phillips, P.W.; Slamet-Loedin, I.H.; Dhugga, K.S.; Campos, H.; Gutterson, N. Genome-edited crops for improved food security of smallholder farmers. Nat. Genet. 2022, 54, 364–367. [Google Scholar] [CrossRef]
- Borrelli, V.M.; Brambilla, V.; Rogowsky, P.; Marocco, A.; Lanubile, A. The enhancement of plant disease resistance using CRISPR/Cas9 technology. Front. Plant Sci. 2018, 9, 1245. [Google Scholar] [CrossRef]
- Nekrasov, V.; Wang, C.; Win, J.; Lanz, C.; Weigel, D.; Kamoun, S. Rapid generation of a transgene-free powdery mildew resistant tomato by genome deletion. Sci. Rep. 2017, 7, 482. [Google Scholar] [CrossRef]
- Paul, N.C.; Park, S.W.; Liu, H.; Choi, S.; Ma, J.; MacCready, J.S.; Chilvers, M.I.; Sang, H. Plant and fungal genome editing to enhance plant disease resistance using the CRISPR/Cas9 system. Front. Plant Sci. 2021, 12, 700925. [Google Scholar] [CrossRef]
- Tyagi, S.; Kumar, R.; Kumar, V.; Won, S.Y.; Shukla, P. Engineering disease resistant plants through CRISPR-Cas9 technology. GM Crops Food 2021, 12, 125–144. [Google Scholar] [CrossRef]
- Zhang, M.; Liu, Q.; Yang, X.; Xu, J.; Liu, G.; Yao, X.; Ren, R.; Xu, J.; Lou, L. CRISPR/Cas9-mediated mutagenesis of Clpsk1 in watermelon to confer resistance to Fusarium oxysporum f. sp. niveum. Plant Cell Rep. 2020, 39, 589–595. [Google Scholar] [CrossRef]
- Brauer, E.K.; Balcerzak, M.; Rocheleau, H.; Leung, W.; Schernthaner, J.; Subramaniam, R.; Ouellet, T. Genome editing of a deoxynivalenol-induced transcription factor confers resistance to Fusarium graminearum in wheat. Mol. Plant-Microbe Interact. 2020, 33, 553–560. [Google Scholar] [CrossRef]
- Zaidi, S.S.; Mahas, A.; Vanderschuren, H.; Mahfouz, M.M. Engineering crops of the future: CRISPR approaches to develop climate-resilient and disease-resistant plants. Genome Biol. 2020, 21, 289. [Google Scholar] [CrossRef]
- Ahmad, I.; Mazhar, K.; Atiq, M.; Khalaf, A.K.; Rashid, M.H.; Asif, M.; Ahmed, S.; Adil, Z.; Fayyaz, A.; Al-Sadoon, M.K.; et al. Epidemiology and management of Fusarium wilt of Eucalyptus camaldulensis through systemic acquired resistance. PeerJ 2024, 12, e17022. [Google Scholar] [CrossRef]
- Ejaz, M.R.; Jaoua, S.; Ahmadi, M.; Shabani, F. An examination of how climate change could affect the future spread of Fusarium spp. around the world, using correlative models to model the changes. Environ. Technol. Innov. 2023, 31, 103177. [Google Scholar] [CrossRef]
- Liu, C.; Kogel, K.H.; Ladera-Carmona, M. Harnessing RNA interference for the control of Fusarium species: A critical review. Mol. Plant Pathol. 2024, 25, e70011. [Google Scholar] [CrossRef]
- Araújo, M.B.; New, M. Ensemble forecasting of species distributions. Trends Ecol. Evol. 2007, 22, 42–47. [Google Scholar] [CrossRef]
- Alagendran, S.; Prasanna, G.S. Climate-adapted pathogen control through biological and chemical agents. In Plant Pathology in the Age of Climate Change, 1st ed.; Golden Leaf Publishers: Uttar Pradesh, India, 2024; pp. 244–263. ISBN 978-93-48240-31-6. [Google Scholar]
- De Wolf, E.D.; Madden, L.V.; Lipps, P.E. Risk assessment models for wheat Fusarium head blight epidemics based on within-season weather data. Phytopathology 2003, 93, 428–435. [Google Scholar] [CrossRef]
- Beernink, B.M.; Amanat, N.; Li, V.H.; Manchur, C.L.; Whyard, S.; Belmonte, M.F. SIGS vs. HIGS: Opportunities and challenges of RNAi pest and pathogen control strategies. Can. J. Plant Pathol. 2024, 46, 675–689. [Google Scholar] [CrossRef]
- Koch, A.; Biedenkopf, D.; Furch, A.; Weber, L.; Rossbach, O.; Abdellatef, E.; Linicus, L.; Johannsmeier, J.; Jelonek, L.; Goesmann, A.; et al. An RNAi-based control of Fusarium graminearum infections through spraying of long dsRNAs involves a plant passage and is controlled by the fungal silencing machinery. PLoS Pathog. 2016, 12, e1005901. [Google Scholar] [CrossRef]
- Padilla-Roji, I.; Ruiz-Jiménez, L.; Bakhat, N.; Vielba-Fernández, A.; Pérez-García, A.; Fernández-Ortuño, D. RNAi technology: A new path for the research and management of obligate biotrophic phytopathogenic fungi. Int. J. Mol. Sci. 2023, 24, 9082. [Google Scholar] [CrossRef]
- Machado, A.K.; Brown, N.A.; Urban, M.; Kanyuka, K.; Hammond-Kosack, K.E. RNAi as an emerging approach to control Fusarium head blight disease and mycotoxin contamination in cereals. Pest Manag. Sci. 2018, 74, 790–799. [Google Scholar] [CrossRef]
- Huang, X.N.; Wang, Y.; Li, Y.T.; Xiao, Y.; Ouyang, S.Q. Trans-kingdom sRNA silencing in the prevention and control of crop Fusarium wilt disease. Phytopathol. Res. 2025, 7, 18. [Google Scholar] [CrossRef]
- Ouyang, S.Q.; Ji, H.M.; Feng, T.; Luo, S.J.; Cheng, L.; Wang, N. Artificial trans-kingdom RNAi of FolRDR1 is a potential strategy to control tomato wilt disease. PLoS Pathog. 2023, 19, e1011463. [Google Scholar] [CrossRef]
- Qiao, L.; Lan, C.; Capriotti, L.; Ah-Fong, A.; Nino Sanchez, J.; Hamby, R.; Heller, J.; Zhao, H.; Glass, N.L.; Judelson, H.S.; et al. Spray-induced gene silencing for disease control is dependent on the efficiency of pathogen RNA uptake. Plant Biotechnol. J. 2021, 19, 1756–1768. [Google Scholar] [CrossRef]
- Werner, B.T.; Gaffar, F.Y.; Schuemann, J.; Biedenkopf, D.; Koch, A.M. RNA-spray-mediated silencing of Fusarium graminearum AGO and DCL genes improve barley disease resistance. Front. Plant Sci. 2020, 11, 476. [Google Scholar] [CrossRef]
- Yang, W.; Wang, B.; Lei, G.; Chen, G.; Liu, D. Advances in nanocarriers to improve the stability of dsRNA in the environment. Front. Bioeng. Biotechnol. 2022, 10, 974646. [Google Scholar] [CrossRef]
- Hanamasagar, Y.; Ramesha, N.M.; Mahapatra, S.; Panigrahi, C.K.; Vidhya, C.S.; Agnihotri, N.; Satapathy, S.N. Advancing RNAi for sustainable insect management: Targeted solutions for eco-friendly pest control. J. Exp. Agric. Int. 2024, 46, 740–745. [Google Scholar] [CrossRef]
- Lamichhane, S.; Neupane, S.; Timsina, S.; Chapagain, B.; Paudel, P.P.; Rimal, A. Potato late blight caused by Phytophthora infestans: An overview on pathology, integrated disease management approaches, and forecasting models. Soc. Sci. Res. Netw. 2024, 4, 105–118. [Google Scholar] [CrossRef]
- Gerlach, W.; Nirenberg, H. The Genus Fusarium: A Pictorial Atlas; Inst. Mikrobiol.: Berlin-Dahlem, Germany, 1982. [Google Scholar]
- Burgess, L.W.; Liddell, C.M.; Summerell, B.A. Laboratory manual for Fusarium Research, 2nd ed.; Fusarium Research Laboratory: University Park, PA, USA, 1988. [Google Scholar]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual; Blackwell Publishing: Oxford, UK, 2006. [Google Scholar] [CrossRef]
- Mulè, G.; González-Jaén, M.T.; Hornok, L.; Nicholson, P.; Waalwijk, C. Advances in molecular diagnosis of toxigenic Fusarium species: A review. Food Addit. Contam. 2005, 22, 316–323. [Google Scholar] [CrossRef]
- Jurado, M.; Vázquez, C.; Marín, S.; Sanchis, V.; González-Jaén, M.T. PCR-based strategy to detect contamination with mycotoxigenic Fusarium species in maize. Syst. Appl. Microbiol. 2006, 29, 681–689. [Google Scholar] [CrossRef]
- Gutiérrez-Aguirre, I.; Rački, N.; Dreo, T.; Ravnikar, M. Droplet digital PCR for absolute quantification of pathogens. In Plant Pathology: Methods in Molecular Biology; Lacomme, C., Ed.; Humana Press: Totowa, NJ, USA, 2015; Volume 1302, pp. 431–442. [Google Scholar] [CrossRef]
- Morcia, C.; Tumino, G.; Gasparo, G.; Ceresoli, C.; Fattorini, C.; Ghizzoni, R.; Carnevali, P.; Terzi, V. Moving from qPCR to Chip Digital PCR Assays for tracking of some Fusarium species causing Fusarium head blight in cereals. Microorganisms 2020, 8, 1307. [Google Scholar] [CrossRef]
- Natarajan, S.; Bucur, D.; Kildea, S.; Doohan, F. Digital PCR assays for quantifying trichothecene-producing Fusarium species, including Fusarium langsethiae, F. poae, and F. sporotrichioides, in oats. Anal. Bioanal. Chem. 2025, 417, 2957–2969. [Google Scholar] [CrossRef]
- Li, B.; Du, J.; Lan, C.; Liu, P.; Weng, Q.; Chen, Q. Development of a loop-mediated isothermal amplification assay for rapid and sensitive detection of Fusarium oxysporum f. sp. cubense race 4. Eur. J. Plant Pathol. 2013, 135, 903–911. [Google Scholar] [CrossRef]
- Lovera, A.; Rodríguez, E.; Simbaqueba, J.; Burbano-David, D.; Carmona, S.L.; Izquierdo-García, L.F.; Gómez-Correa, J.C.; Leon-Pacheco, R.I.; Parra-Fuentes, M.; Rodríguez-Yzquierdo, G.; et al. Development and application of a droplet digital PCR assay for the detection of Fusarium oxysporum f. sp. cubense tropical race 4 from different biological samples. Plant Pathology. 2023, 73, 342–354. [Google Scholar] [CrossRef]
- Notomi, T.; Okayama, H.; Masubuchi, H.; Yonekawa, T.; Watanabe, K.; Amino, N.; Hase, T. Loop-mediated isothermal amplification of DNA. Nucleic Acids Res. 2000, 28, e63. [Google Scholar] [CrossRef]
- Li, Y.; Fan, P.; Zhou, S.; Zhang, L. Loop-mediated isothermal amplification (LAMP): A novel rapid detection platform for pathogens. Microb. Pathog. 2017, 107, 54–61. [Google Scholar] [CrossRef]
- Ghosh, R.; Nagavardhini, A.; Sengupta, A.; Sharma, M. Development of loop-mediated isothermal amplification (LAMP) assay for rapid detection of Fusarium oxysporum f. sp. ciceris—Wilt pathogen of chickpea. BMC Res. Notes 2015, 8, 40. [Google Scholar] [CrossRef]
- Jiang, H.; Wu, N.; Jin, S.; Ahmed, T.; Wang, H.; Li, B.; Wu, X.; Bao, Y.; Liu, F.; Zhang, J.-Z. Identification of rice seed-derived Fusarium spp. and development of LAMP assay against Fusarium fujikuroi. Pathogens 2021, 10, 1. [Google Scholar] [CrossRef]
- Ghosh, D.K.; Kokane, S.B.; Kokane, A.D.; Warghane, A.J.; Motghare, M.R.; Bhose, S.; Sharma, A.K.; Reddy, M.K. Development of a recombinase polymerase based isothermal amplification combined with lateral flow assay (HLB-RPA-LFA) for rapid detection of “Candidatus Liberibacter asiaticus”. PLoS ONE 2018, 13, e0208530. [Google Scholar] [CrossRef]
- Hu, S.; Yan, C.; Yu, H.; Zhang, Y.; Zhang, C.Q. Establishment of the Recombinase Polymerase Amplification-Lateral Flow Dipstick Detection Technique for Fusarium oxysporum. Plant Dis. 2023, 107, 2665–2672. [Google Scholar] [CrossRef]
- Liang, X.; Zhang, X.; Haseeb, H.A.; Tang, T.; Shan, J.; Yin, B.; Guo, W. Development and evaluation of a novel visual and rapid detection assay for toxigenic Fusarium graminearum in maize based on recombinase polymerase amplification and lateral flow analysis. Int. J. Food Microbiol. 2022, 372, 109682. [Google Scholar] [CrossRef]
- Cobo-Díaz, J.F.; Baroncelli, R.; Le Floch, G.; Picot, A. A novel metabarcoding approach to investigate Fusarium species composition in soil and plant samples. FEMS Microbiol. Ecol. 2019, 95, fiz084. [Google Scholar] [CrossRef]
- Walder, F.; Schlaeppi, K.; Wittwer, R.; van der Heijden, M.G.A.; Vogelgsang, S. Community profiling of Fusarium and other plant-associated fungi in crops using SMRT sequencing. Front. Plant Sci. 2017, 8, 2019. [Google Scholar] [CrossRef]
- Karlsson, I.; Persson, P.; Friberg, H. Fusarium head blight from a microbiome perspective. Front. Microbiol. 2021, 12, 628373. [Google Scholar] [CrossRef]
- Pieczul, K.; Świerczyńska, I.; Wójtowicz, A. Advanced rDNA-based detection of wheat pathogens in grain samples using NGS. Pathogens 2025, 14, 164. [Google Scholar] [CrossRef]
- Martinez, A.; Bennett, J.W. Fungal volatile organic compounds. In Encyclopedia of Mycology; Zaragoza, O., Ed.; Elsevier: Amsterdam, The Netherlands, 2021; Volume 1, pp. 239–245. [Google Scholar]
- Cui, S.; Ling, P.; Zhu, H.; Keener, H.M. Plant pest detection using an artificial nose system: A review. Sensors 2018, 18, 378. [Google Scholar] [CrossRef]
- Cellini, A.; Blasioli, S.; Biondi, E.; Bertaccini, A.; Braschi, I.; Spinelli, F. Potential applications and limitations of electronic nose devices for plant disease diagnosis. Sensors 2017, 17, 2596. [Google Scholar] [CrossRef]
- Infantino, A.; Costa, C.; Aragona, M.; Reverberi, M.; Taiti, C.; Mancuso, S. Identification of different Fusarium spp. through mVOCs profiling by means of proton-transfer-reaction time-of-flight (PTR-ToF-MS) analysis. J. Plant Pathol. 2017, 99, 663–669. [Google Scholar] [CrossRef]
- Camardo Leggieri, M.; Mazzoni, M.; Bertuzzi, T.; Moschini, M.; Prandini, A.; Battilani, P. Electronic nose for the rapid detection of deoxynivalenol in wheat using classification and regression trees. Toxins 2022, 14, 617. [Google Scholar] [CrossRef]
- Bu’sko, M.; Gracka, A.; Jele’n, H.; Szablewska, K.S.; Przybylska-Balcerek, A.; Szwajkowska-Michałek, L.; Góral, T. The effect of organic and conventional cultivation systems on the profile of Volatile Organic Compounds in winter wheat grain, including susceptibility to Fusarium Head Blight. Metabolites 2023, 13, 1045. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, B.; Dong, J.; Tian, Y.; Lin, Y.; Fang, G.; Wang, S. Identification of mouldy rice using an electronic nose combined with SPME-GC/MS. J. Stored Prod. Res. 2022, 95, 101921. [Google Scholar] [CrossRef]
- Wesoly, M.; Daulton, E.; Jenkins, S.; van Amsterdam, S.; Clarkson, J.; Covington, J.A. Early detection of Fusarium basal rot infection in onions and shallots based on VOC profiles analysis. J. Agric. Food Chem. 2024, 72, 3664–3672. [Google Scholar] [CrossRef]
- Rizzi, L.; Galuppi, R.; Simioli, M.; Canestrari, G.; Bonoli, C.; Tampieri, M.P. Grains colonised by moulds: Fungal identification and headspace analysis of produced volatile metabolites. Ital. J. Anim. Sci. 2009, 8 (Suppl. S2), 334–336. [Google Scholar] [CrossRef]
- Infantino, A.; Taiti, C.; Grottoli, A.; Mancuso, S.; Costa, C.; Garzoli, S. Examination of volatile signatures of Fusarium bulb rot in garlic using PTR-ToF-MS and SPME-GC/MS. Separations 2023, 10, 556. [Google Scholar] [CrossRef]
- Kamle, M.; Mahato, D.K.; Devi, S.; Kumar, P.; Patil, R.K.; Kang, S.G.; Kumar, A. Fumonisins: Impact on agriculture, food, and human health and their management strategies. Toxins 2019, 11, 328. [Google Scholar] [CrossRef]
- International Agency for Research on Cancer (IARC). Fumonisin B1. In IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; IARC Press: Lyon, France, 2002; Volume 82. [Google Scholar]
- Mafe, A.N.; Büsselberg, D. Mycotoxins in food: Cancer risks and strategies for control. Foods 2024, 13, 3502. [Google Scholar] [CrossRef]
- Pestka, J.J.; Smolinski, A.T. Deoxynivalenol: Toxicology and potential effects on humans. J. Toxicol. Environ. Health Part B 2005, 8, 39–69. [Google Scholar] [CrossRef]
- Kamle, M.; Devi, S.; Mahato, D.K.; Mishra, S.; Kumar, A. Occurrence, toxicity, and risk assessment of deoxynivalenol and its modified forms in food. Toxins 2022, 14, 187. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA). Scientific opinion on the risks for animal and public health related to the presence of T-2 and HT-2 toxin in food and feed. EFSA J. 2017, 15, e04655. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA). Scientific opinion on the risks for public health related to the presence of zearalenone in food. EFSA J. 2011, 9, 2197. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA). Risks to human and animal health related to the presence of deoxynivalenol (DON) in food and feed. EFSA J. 2017, 15, e04718. [Google Scholar] [CrossRef]
- Streit, E.; Schatzmayr, G.; Tassis, P.; Tzika, E.; Marin, D.; Taranu, I.; Puel, O.; Oswald, I.P. Current situation of mycotoxin contamination and co-occurrence in animal feed—Focus on Europe. Toxins 2012, 4, 788–809. [Google Scholar] [CrossRef]
- Pestka, J.J. Deoxynivalenol: Mechanisms of action, human exposure, and toxicological relevance. Arch. Toxicol. 2010, 84, 663–679. [Google Scholar] [CrossRef]
- Qie, Y.; Liu, C.; Yang, C.; Zhang, H.; Wang, J.; Shen, C. Occurrence and toxicity of beauvericin: A review. Toxins 2024, 16, 45. [Google Scholar] [CrossRef]
- Jestoi, M. Emerging Fusarium mycotoxins fusaproliferin, beauvericin, enniatins, and moniliformin—A review. Crit. Rev. Food Sci. Nutr. 2008, 48, 21–49. [Google Scholar] [CrossRef]
- Zinedine, A.; Soriano, J.M.; Moltó, J.C.; Mañes, J. Review on the toxicity, occurrence, metabolism, detoxification, regulations and intake of zearalenone: An oestrogenic mycotoxin. Food Chem. Toxicol. 2007, 45, 1–18. [Google Scholar] [CrossRef]
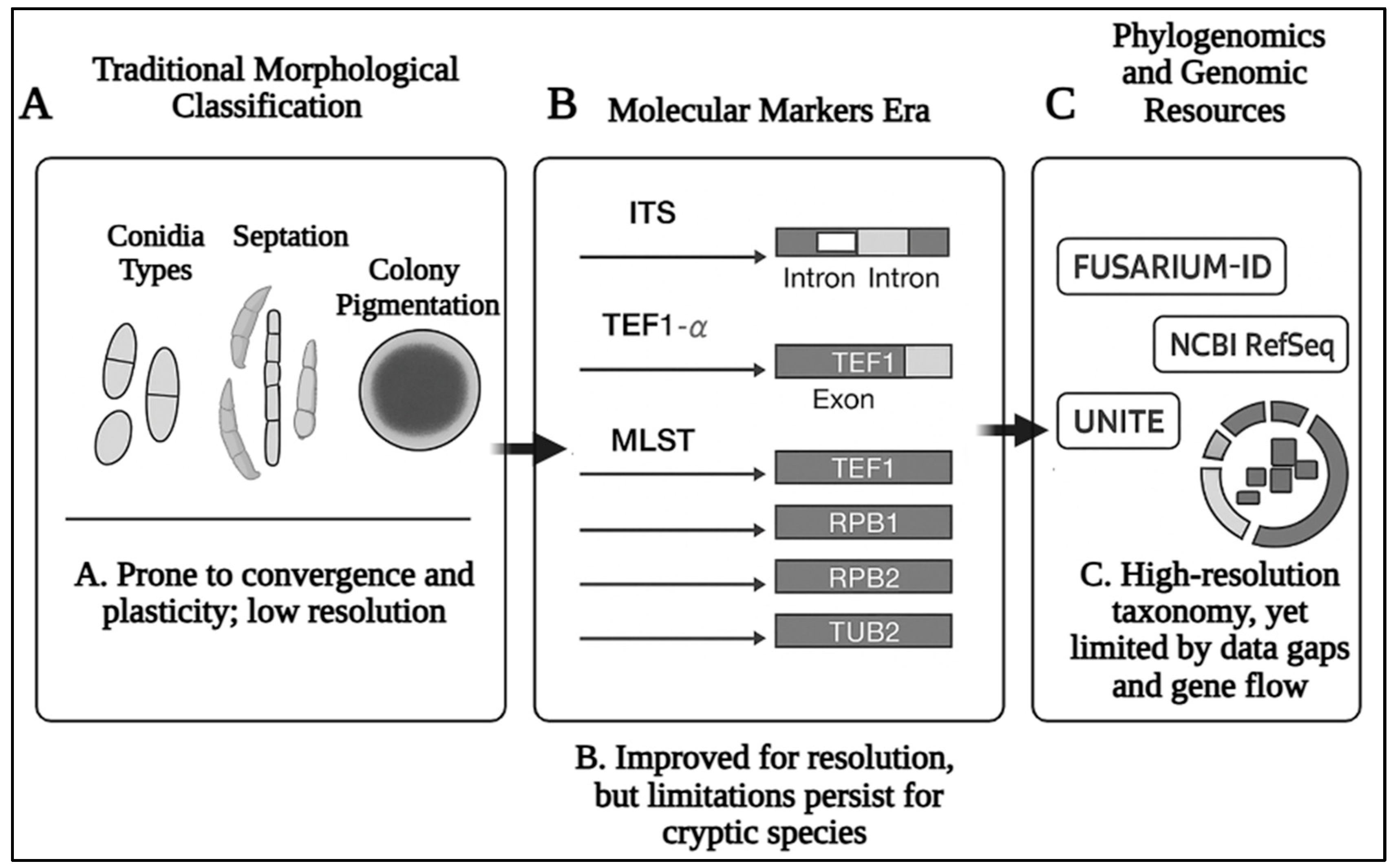
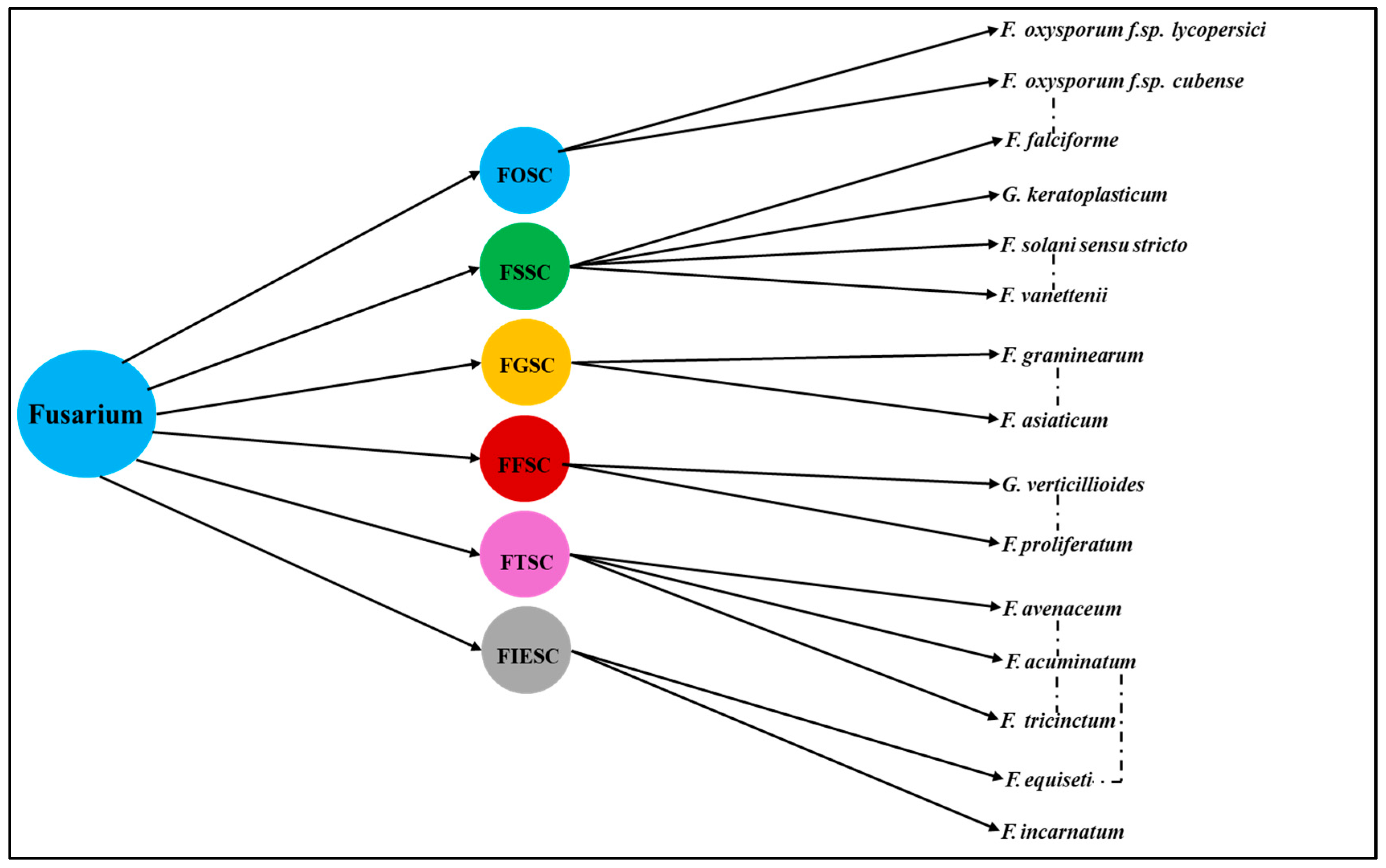
| Transcription Factor | Fusarium Species | Function | Regulates | References |
|---|---|---|---|---|
| Tri6 | F. graminearum | Zn(II)2Cys6 TF; pathway-specific regulator | Trichothecene biosynthesis gene cluster (TRI genes); also acts as global regulator | [122,123] |
| Tri10 | F. graminearum | Transcriptional activator | Activates TRI gene expression in coordination with Tri6 | [122] |
| PacC | F. oxysporum | pH-responsive TF | Virulence gene expression under neutral-alkaline pH; adaptation to host environment | [124] |
| Ftf1 | F. oxysporum | Zn(II)2Cys6 TF encoded on accessory chromosome | Effector gene expression and host-specific pathogenicity | [125] |
| Sge1 | F. oxysporum | Global TF involved in effector regulation | SIX gene expression (secreted in xylem proteins) | [126] |
| FgLaeA (part of VeA complex) | F. graminearum | Global regulator of secondary metabolism, sexual development, and virulence | Regulates the expression of trichothecene biosynthetic genes (TRI6, ZEB2), sexual development genes, and virulence factors via interaction with FgVeA | [127] |
| Kmt6 (H3K27 methyltransferase) | F. fujikuroi | Epigenetic regulator of secondary metabolism via histone modification | Controls the activation of silent secondary metabolite gene clusters by modifying chromatin accessibility through H3K27me3 marks | [128] |
| AreA | F. fujikuroi | Nitrogen metabolism TF | Gibberellin biosynthesis genes (in response to nitrogen availability) | [129] |
| Fusarium Species | Host Crop | Disease | Resistance Gene/Locus | References |
|---|---|---|---|---|
| F. graminearum | Wheat | Fusarium head blight |
| [133] |
| F. oxysporum f. sp. lycopersici | Tomato | Fusarium wilt |
| [134] |
| F. oxysporum f. sp. lentis | Lentil | Fusarium wilt/root rot | Fw Gene | [135,136] |
| F. virguliforme (syn. F. solani f. sp. Glycines) | Soybean | Sudden death syndrome |
| [137] |
| F. verticillioides | Maize | Ear rot, seedling blight |
| [138,139] |
| F. oxysporum f. sp. cubense tropical race 4 (Tr4) | Banana | Fusarium wilt (Panama disease) |
| [140] |
| F. oxysporum f. sp. vasinfectum | Cotton | Fusarium wilt |
| [141] |
| F. oxysporum f. sp. cucumerinum | Cucumber | Fusarium wilt |
| [142,143] |
| Fusarium Species | Known for Formae speciales? | Notable Formae speciales (f. sp.) |
|---|---|---|
| F. oxysporum | Yes—over 100 | f. sp. lycopersici (tomato), cubense (banana), vasinfectum (cotton), cepae (onion), melonis (melon), pisi (pea) |
| F. solani (FSSC) | Yes, but being revised | f. sp. pisi (pea), phaseoli (bean), cucurbitae (cucurbits), mori (mulberry) |
| F. graminearum | No | Not applicable—broad host range |
| F. verticillioides | No | Not applicable—infects maize, sorghum, and others broadly |
| F. proliferatum | No | Not applicable—opportunistic across many hosts |
| F. avenaceum | No | Not applicable—generalist necrotrophy |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bugingo, C.; Infantino, A.; Okello, P.; Perez-Hernandez, O.; Petrović, K.; Turatsinze, A.N.; Moparthi, S. From Morphology to Multi-Omics: A New Age of Fusarium Research. Pathogens 2025, 14, 762. https://doi.org/10.3390/pathogens14080762
Bugingo C, Infantino A, Okello P, Perez-Hernandez O, Petrović K, Turatsinze AN, Moparthi S. From Morphology to Multi-Omics: A New Age of Fusarium Research. Pathogens. 2025; 14(8):762. https://doi.org/10.3390/pathogens14080762
Chicago/Turabian StyleBugingo, Collins, Alessandro Infantino, Paul Okello, Oscar Perez-Hernandez, Kristina Petrović, Andéole Niyongabo Turatsinze, and Swarnalatha Moparthi. 2025. "From Morphology to Multi-Omics: A New Age of Fusarium Research" Pathogens 14, no. 8: 762. https://doi.org/10.3390/pathogens14080762
APA StyleBugingo, C., Infantino, A., Okello, P., Perez-Hernandez, O., Petrović, K., Turatsinze, A. N., & Moparthi, S. (2025). From Morphology to Multi-Omics: A New Age of Fusarium Research. Pathogens, 14(8), 762. https://doi.org/10.3390/pathogens14080762