
Evaluation of Fusarium Head Blight Resistance Through a Genome-Wide Association Study in CIMMYT and South Asian Wheat Germplasm

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methodology
2.1. Plant Materials
2.2. Field Trials and Disease Scoring
2.3. Statistical Analysis
2.4. Genotyping, Population Structure, and LD Analysis
2.5. Marker-Traits Association (MTA) for FHB
3. Results
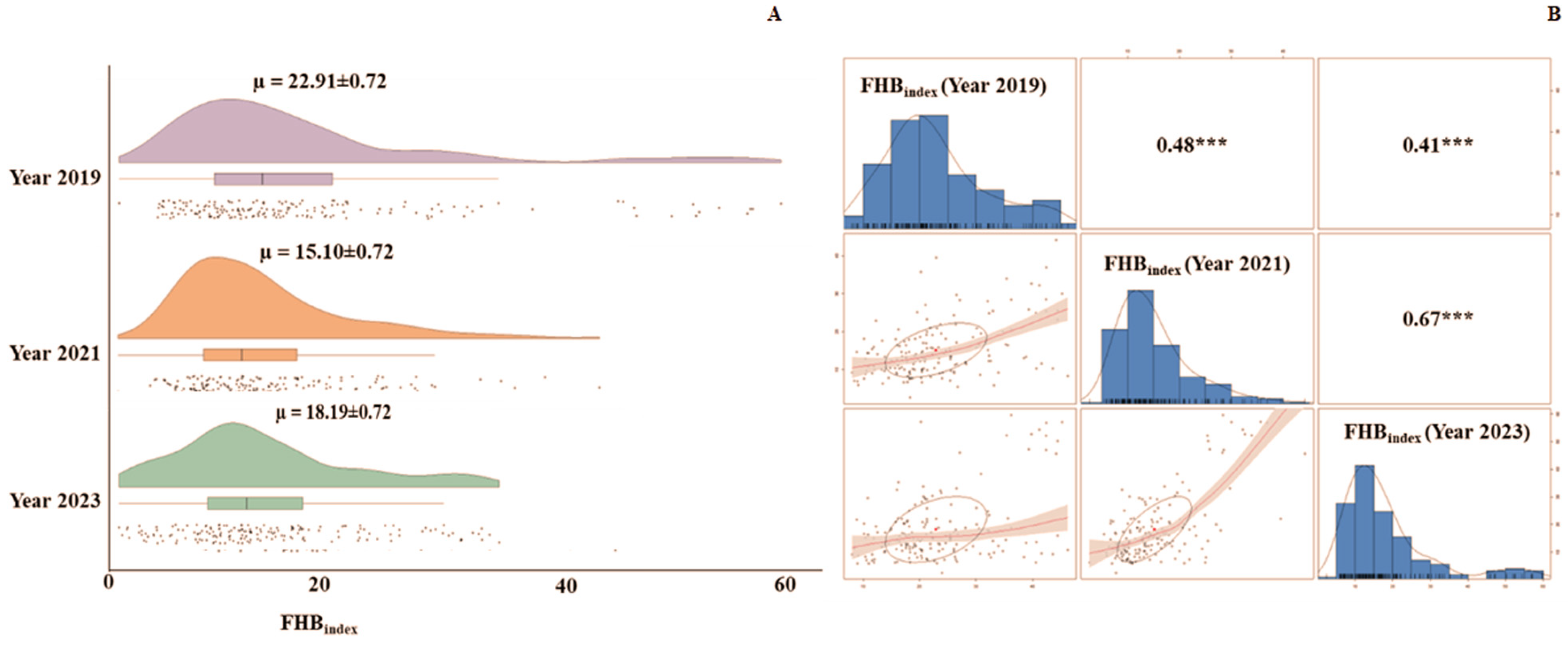
3.1. Reaction of Genotypes Against FHB and Identification of Resistant Lines
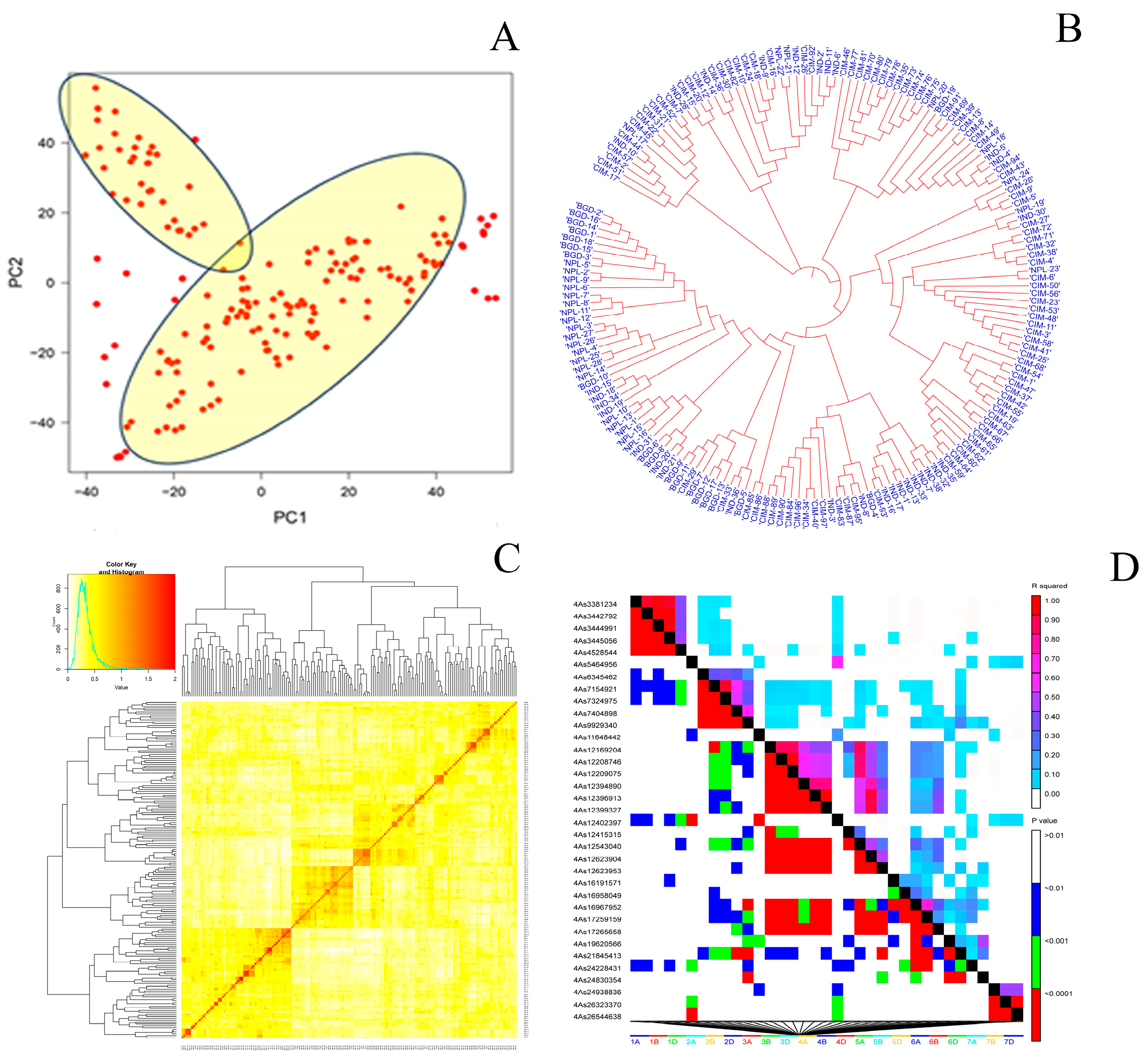
3.2. Genotyping, PCA, Kinship and Linkage Disequilibrium Analysis
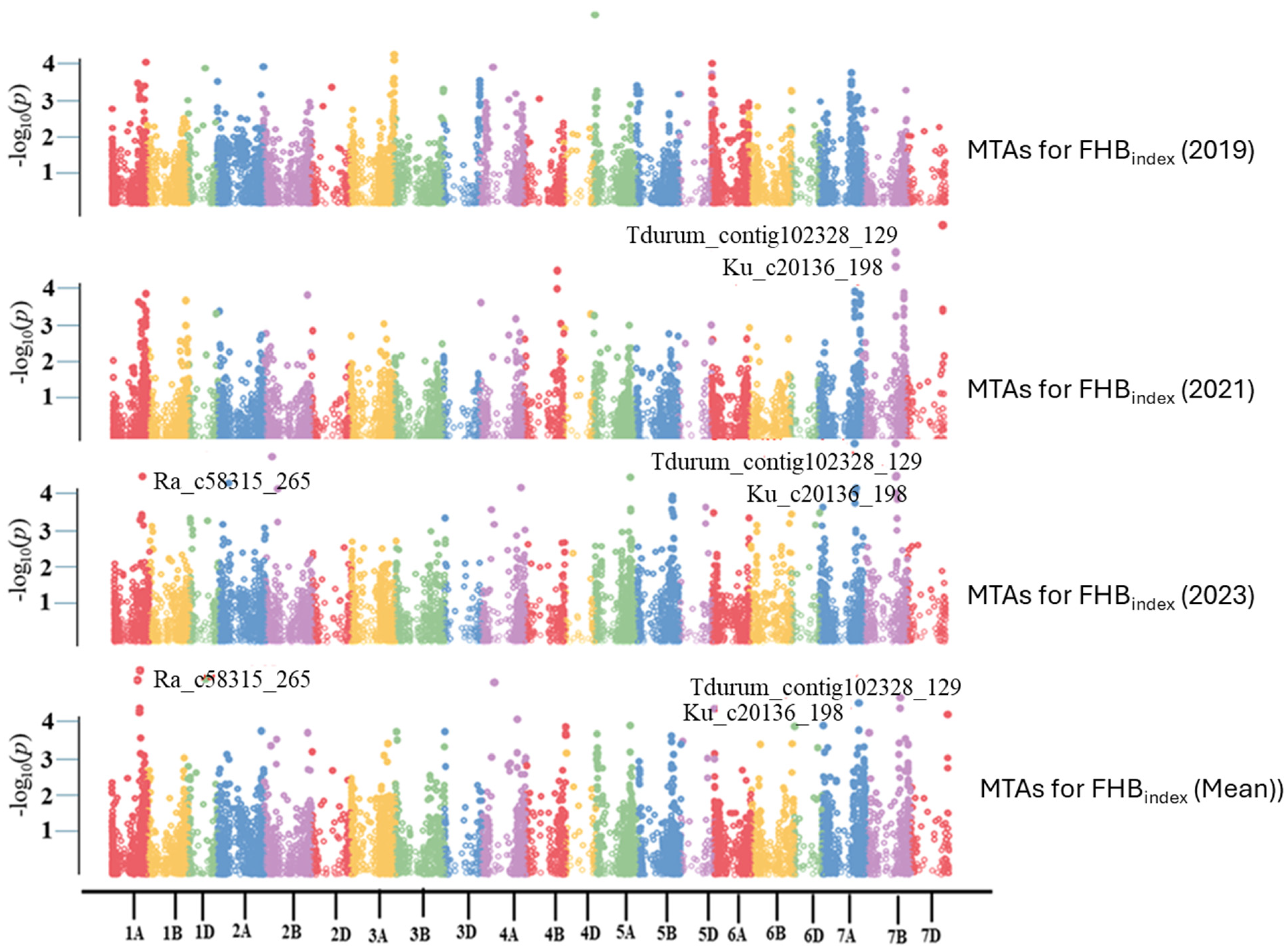
3.3. Genome-Wide Association Study
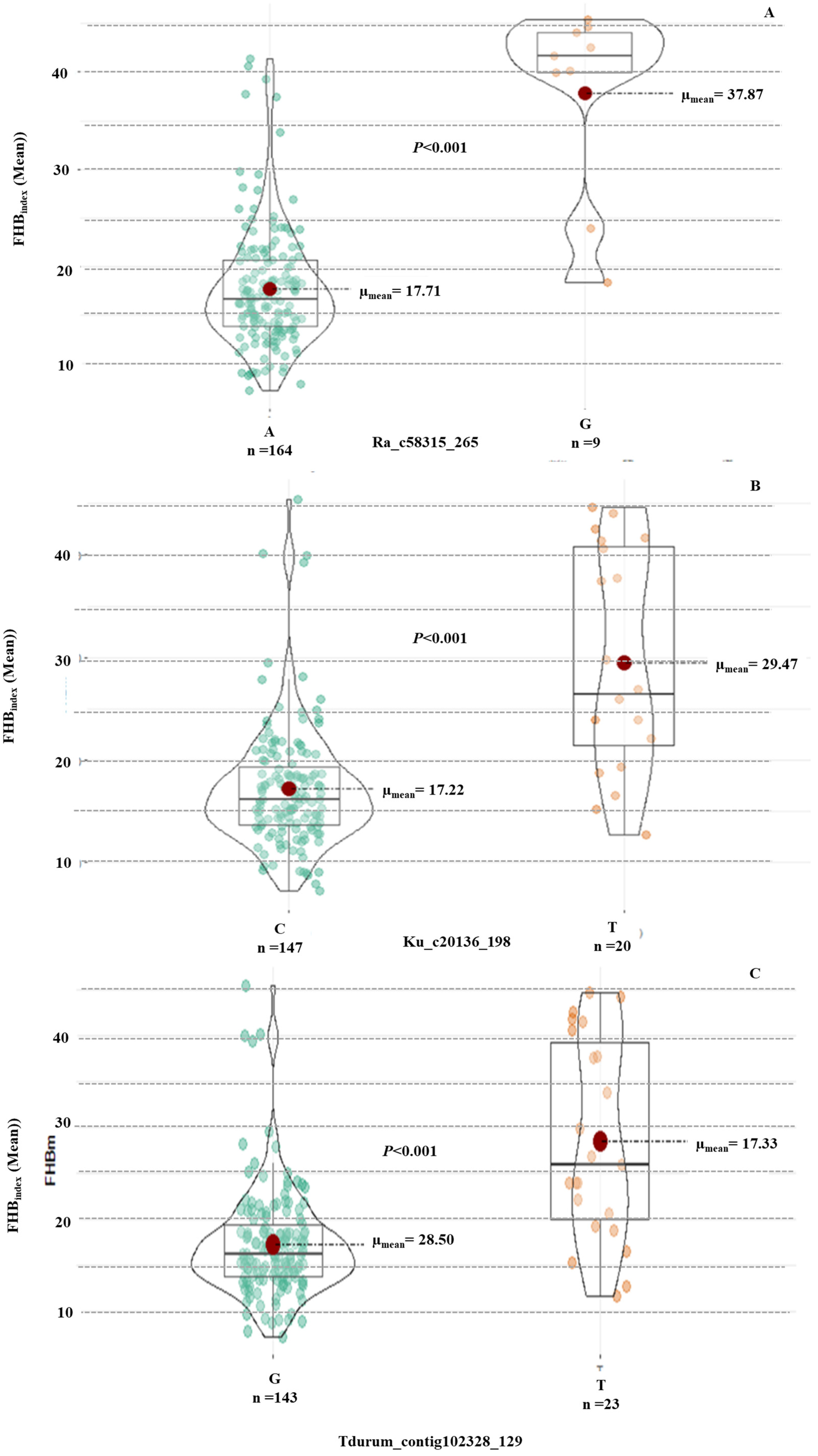
3.4. Candidate Genes for the Significant MTA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. 2024. Available online: https://www.fao.org/faostat/en/#data (accessed on 2 May 2025).
- Khan, M.K.; Pandey, A.; Athar, T.; Choudhary, S.; Deval, R.; Gezgin, S.; Hamurcu, M.; Topal, A.; Atmaca, E.; Santos, P.A.; et al. Fusarium head blight in wheat: Contemporary status and molecular approaches. 3 Biotech 2020, 10, 172. [Google Scholar] [CrossRef] [PubMed]
- Friesen, T.L.; Faris, J.D.; Solomon, P.S.; Oliver, R.P. Host-specific toxins: Effectors of necrotrophic pathogenicity. Cell Microbiol. 2008, 10, 1421–1428. [Google Scholar] [CrossRef]
- Gurung, S.; Hansen, J.M.; Bonman, J.M.; Gironella, A.I.N.; Adhikari, T.B. Multiple disease resistance to four leaf spot diseases in winter wheat accessions from the USDA National Small Grains Collection. Crop Sci. 2012, 52, 1640–1650. [Google Scholar] [CrossRef]
- Nganje, W.E.; Kaitibie, S.; Wilson, W.W.; Leistritz, F.L.; Bangsund, D.A. Economic Impacts of Fusarium Head Blight in Wheat and Barley: 1993–2001 (Agribusiness and Applied Economics Report No 538); Department of Agribusiness and Applied Economics, Agricultural Experiment Station, North Dakota State University: Fargo, ND, USA, 2004. [Google Scholar]
- McMullen, M.; Bergstrom, G.; De Wolf, E.; Dill-Macky, R.; Hershman, D.; Shaner, G.; Van Sanford, D. A unified effort to fight an enemy of wheat and barley: Fusarium head blight. Plant Dis. 2012, 96, 1712–1728. [Google Scholar] [CrossRef]
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The global burden of pathogens and pests on major food crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef]
- Shude, S.P.N.; Yobo, K.S.; Mbili, N.C. Progress in the management of Fusarium head blight of wheat: An overview. S. Afr. J. Sci. 2020, 116, 11–12. [Google Scholar] [CrossRef]
- Mesterházy, Á. Methodology of resistance testing and breeding against Fusarium head blight in wheat and results of the selection. Cereal Res. Commun. 1997, 25, 631–637. [Google Scholar] [CrossRef]
- Mesterházy, Á.; Bartók, T.; Mirocha, C.; Komoroczy, R. Nature of wheat resistance to Fusarium head blight and the role of deoxynivalenol for breeding. Plant Breed. 1999, 118, 97–110. [Google Scholar] [CrossRef]
- Gilbert, J.; Tekauz, A. Recent developments in research on Fusarium head blight of wheat in Canada. Can. J. Plant Pathol. 2000, 22, 1–8. [Google Scholar] [CrossRef]
- Mesterhazy, A.; Bartok, T.; Lamper, C. Influence of wheat cultivar, species of Fusarium, and isolate aggressiveness on the efficacy of fungicides for control of Fusarium head blight. Plant Dis. 2003, 87, 1107–1115. [Google Scholar] [CrossRef]
- Sirangelo, T.M. Molecular investigations to improve Fusarium head blight resistance in wheat: An update focusing on multi-omics approaches. Plants 2024, 13, 2179. [Google Scholar] [CrossRef] [PubMed]
- Kollers, S.; Rodemann, B.; Ling, J.; Korzun, V.; Ebmeyer, E.; Argillier, O.; Hinze, M.; Plieske, J.; Kulosa, D.; Ganal, M.W.; et al. Whole genome association mapping of Fusarium head blight resistance in European winter wheat (Triticum aestivum L.). PLoS ONE 2013, 8, e57500. [Google Scholar] [CrossRef]
- Arruda, M.P.; Brown, P.; Brown-Guedira, G.; Krill, A.M.; Thurber, C.; Merrill, K.R.; Foresman, B.J.; Kolb, F.L. Genome-wide association mapping of Fusarium head blight resistance in wheat using genotyping-by-sequencing. Plant Genome 2016, 9, 1–14. [Google Scholar] [CrossRef]
- Buerstmayr, H.; Ban, T.; Anderson, J.A. QTL mapping and marker-assisted selection for Fusarium head blight resistance in wheat: A review. Plant Breed. 2009, 128, 1–26. [Google Scholar] [CrossRef]
- Buerstmayr, M.; Steiner, B.; Buerstmayr, H.; Léon, J. Breeding for Fusarium head blight resistance in wheat—Progress and challenges. Plant Breed. 2019, 139, 429–454. [Google Scholar] [CrossRef]
- Cai, J.; Wang, S.; Su, Z.; Li, T.; Zhang, X.; Bai, G. Meta-analysis of QTL for Fusarium head blight resistance in Chinese wheat landraces. Crop J. 2019, 7, 784–798. [Google Scholar] [CrossRef]
- Liu, S.; Hall, M.D.; Griffey, C.A.; McKendry, A.L. Meta-analysis of QTL associated with Fusarium head blight resistance in wheat. Crop Sci. 2009, 49, 1955–1968. [Google Scholar] [CrossRef]
- Löffler, M.; Schön, C.C.; Miedaner, T. Revealing the genetic architecture of FHB resistance in hexaploid wheat (Triticum aestivum L.) by QTL Meta-analysis. Mol. Breed. 2009, 23, 473–488. [Google Scholar] [CrossRef]
- Venske, E.; Dos Santos, R.S.; Farias, D.D.R.; Rother, V.; Da Maia, L.C.; Pegoraro, C.; Costa de Oliveira, A. Meta-analysis of the QTLome of Fusarium head blight resistance in bread wheat: Refining the current puzzle. Front. Plant Sci. 2019, 10, 727. [Google Scholar] [CrossRef]
- Zheng, T.; Hua, C.; Li, L.; Sun, Z.X.; Yuan, M.M.; Bai, G.H.; Humphreys, G.; Li, T. Integration of meta-QTL discovery with omics: Towards a molecular breeding platform for improving wheat resistance to Fusarium head blight. Crop J. 2021, 9, 739–749. [Google Scholar] [CrossRef]
- Cuthbert, P.A.; Somers, D.J.; Thomas, J.; Cloutier, S.; Brulé-Babel, A. Fine mapping Fhb1, a major gene controlling Fusarium head blight resistance in bread wheat (Triticum aestivum L.). Theor. Appl. Genet. 2006, 112, 1465–1472. [Google Scholar] [CrossRef] [PubMed]
- Cuthbert, P.A.; Somers, D.J.; Brulé-Babel, A. Mapping of Fhb2 on chromosome 6BS: A gene controlling Fusarium head blight field resistance in bread wheat (Triticum aestivum L.). Theor. Appl. Genet. 2007, 114, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Steiner, B.; Buerstmayr, M.; Wagner, C.; Danler, A.; Eshonkulov, B.; Ehn, M.; Buerstmayr, H. Fine-mapping of the Fusarium head blight resistance QTL Qfhs.ifa-5A identifies two resistance QTL associated with anther extrusion. Theor. Appl. Genet. 2019, 132, 2039–2053. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Zhou, J.; Jia, H.; Gao, Z.; Fan, M.; Luo, Y.; Zhao, P.; Xue, S.; Li, N.; Yuan, Y.; et al. Mutation of a histidine-rich calcium-binding-protein gene in wheat confers resistance to Fusarium head blight. Nat. Genet. 2019, 51, 1106–1112. [Google Scholar] [CrossRef]
- Su, Z.; Bernardo, A.; Tian, B.; Chen, H.; Wang, S.; Ma, H.; Cai, S.; Liu, D.; Zhang, D.; Li, T.; et al. A deletion mutation in TaHRC confers Fhb1 resistance to Fusarium head blight in wheat. Nat. Genet. 2019, 51, 1099–1105. [Google Scholar] [CrossRef]
- Wang, H.; Sun, S.; Ge, W.; Zhao, L.; Hou, B.; Wang, K.; Lyu, Z.; Chen, L.; Xu, S.; Guo, J.; et al. Horizontal gene transfer of Fhb7 from fungus underlies Fusarium head blight resistance in wheat. Science 2020, 368, eaba5435. [Google Scholar] [CrossRef]
- Ghimire, B.; Sapkota, S.; Bahri, B.A.; Martinez-Espinoza, A.D.; Buck, J.W.; Mergoum, M. Fusarium head blight and rust diseases in soft red winter wheat in the southeast United States: State of the art, challenges and future perspective for breeding. Front. Plant Sci. 2020, 11, 1080. [Google Scholar] [CrossRef]
- Zhu, Z.; Hao, Y.; Mergoum, M.; Bai, G.; Humphreys, G.; Cloutier, S.; Xia, X.; He, Z. Breeding wheat for resistance to Fusarium head blight in the Global North: China, USA, and Canada. Crop J. 2019, 7, 730–738. [Google Scholar] [CrossRef]
- Cai, X.; Danilova, T.; Charif, A.; Wang, F.; Zhang, W.; Zhang, M.; Ren, S.; Zhu, X.; Zhong, S.; Dykes, L.; et al. Registration of WGC002 spring wheat containing wild grass-derived Fusarium head blight resistance gene Fhb7The2. J. Plant Regist. 2024, 18, 179–186. [Google Scholar] [CrossRef]
- Brar, G.S.; Pozniak, C.J.; Kutcher, H.R.; Hucl, P.J. Evaluation of Fusarium head blight resistance genes Fhb1, Fhb2, and Fhb5 introgressed into elite Canadian hard red spring wheats: Effect on agronomic and end-use quality traits and implications for breeding. Mol. Breed. 2019, 39, 44. [Google Scholar] [CrossRef]
- Schweiger, W.; Steiner, B.; Vautrin, S.; Nussbaumer, T.; Siegwart, G.; Zamini, M.; Jungreithmeier, F.; Gratl, V.; Lemmens, M.; Mayer, K.F.X.; et al. Suppressed recombination and unique candidate genes in the divergent haplotype encoding Fhb1, a major Fusarium head blight resistance locus in wheat. Theor. Appl. Genet. 2016, 129, 1607–1623. [Google Scholar] [CrossRef]
- Saharan, M.S.; Kumar, H.M.A.; Gurjar, M.S.; Aggarwal, R. Fusarium head blight of wheat in India-variability in pathogens associated and sources of resistance: An overview. Indian Phytopathol. 2021, 74, 345–353. [Google Scholar] [CrossRef]
- Kumar, A.; Aggarwal, H.M.; Rashmi, G.M.S.; Prasad, V.L.; Saharan, M.S. Identification of Fusarium head blight resistant sources in wheat under artificially inoculated condition. Indian J. Agric. Sci. 2021, 91, 895–899. [Google Scholar] [CrossRef]
- He, X.; Singh, P.K.; Dreisigacker, S.; Singh, S.; Lillemo, M.; Duveiller, E. Dwarfing genes Rht-B1b and Rht-D1b are associated with both type I FHB susceptibility and low anther extrusion in two bread wheat populations. PLoS ONE 2016, 11, e0162499. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Singh, P.K.; Duveiller, E.; Schlang, N.; Dreisigacker, S.; Singh, R.P. Identification and characterization of international Fusarium head blight screening nurseries of wheat at CIMMYT, Mexico. Eur. J. Plant Pathol. 2013, 136, 123–134. [Google Scholar] [CrossRef]
- He, X.; Singh, P.K.; Duveiller, E.; Dreisigacker, S.; Singh, R.P. Development and characterization of International Maize and Wheat Improvement Center (CIMMYT) germplasm for Fusarium head blight resistance. In Fusarium Head Blight in Latin America; Alconada, T.M., Chulze, S.N., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 241–262. [Google Scholar]
- Zhang, J.; Gill, H.S.; Halder, J.; Brar, N.K.; Ali, S.; Bernardo, A.; Amand, P.S.; Bai, G.; Turnipseed, B.; Sehgal, S.K. Multi-locus genome-wide association studies to characterize Fusarium head blight (FHB) resistance in hard winter wheat. Front. Plant Sci. 2022, 13, 946700. [Google Scholar] [CrossRef] [PubMed]
- Navathe, S.; Pandey, A.K.; Sharma, S.; Chand, R.; Mishra, V.K.; Kumar, D.; Jaiswal, S.; Iquebal, M.A.; Govindan, V.; Joshi, A.K.; et al. New genomic regions identified for resistance to spot blotch and terminal heat stress in an interspecific population of Triticum aestivum and T. spelta. Plants 2022, 11, 2987. [Google Scholar] [CrossRef]
- Osman, M.; He, X.; Singh, R.P.; Duveiller, E.; Lillemo, M.; Pereyra, S.A.; Westerdijk-Hoks, I.; Kurushima, M.; Yau, S.-K.; Benedettelli, S.; et al. Phenotypic and genotypic characterization of CIMMYT’s 15th international Fusarium head blight screening nursery of wheat. Euphytica 2015, 205, 521–537. [Google Scholar]
- He, X.; Brar, G.S.; Bonnett, D.; Dreisigacker, S.; Hyles, J.; Spielmeyer, W.; Bhavani, S.; Singh, R.P.; Singh, P.K. Disease resistance evaluation of elite CIMMYT wheat lines containing the coupled Fhb1 and Sr2 genes. Plant Dis. 2020, 104, 2369–2376. [Google Scholar] [CrossRef]
- Bhatta, M.; Morgounov, A.; Belamkar, V.; Poland, J.; Baenziger, S.P. Unlocking the novel genetic diversity and population structure of synthetic hexaploid wheat. BMC Genom. 2018, 19, 591. [Google Scholar] [CrossRef]
- Shi, C.; Chao, H.; Sun, X.; Suo, Y.; Chen, Z.; Li, Z.; Ma, L.; Li, J.; Ren, Y.; Hua, W.; et al. Genome-wide association study for Fusarium head blight resistance in common wheat from China. Agronomy 2023, 13, 1712. [Google Scholar] [CrossRef]
- Lu, S.; Edwards, M.C. Genome-wide analysis of small secreted cysteine-rich proteins identifies candidate effector proteins potentially involved in Fusarium graminearum−wheat interactions. Phytopathology 2016, 106, 166–176. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Shan, Z.; Yu, J.; Xu, G.; Zhang, Z. The cysteine-rich repeat protein TaCRR1 participates in defense against both Rhizoctonia cerealis and Bipolaris sorokiniana in wheat. Int. J. Mol. Sci. 2020, 21, 5698. [Google Scholar] [CrossRef] [PubMed]
- Paudel, B.; Zhuang, Y.; Galla, A.; Dahal, S.; Qiu, Y.; Ma, A.; Raihan, T.; Yen, Y. WFhb1-1 plays an important role in resistance against Fusarium head blight in wheat. Sci. Rep. 2020, 10, 7794. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Df | Mean Square | F Value | p-Value |
|---|---|---|---|---|
| Year | 2 | 5903.51 | 82.64 | p < 0.001 |
| Genotype | 173 | 327.37 | 4.58 | p < 0.001 |
| Genotype × Year | 198 | 76.12 | 1.07 | 0.28 |
| Rep | 1 | 365.05 | 5.11 | 0.02 |
| Error | 669 | 71.44 |
| Traits | SNP | Chr | Pos | p-Value | Effect |
|---|---|---|---|---|---|
| FHBm | wsnp_Ex_c11055_17927668 | 5D | 561706128 | 0.000794 | 3.06 |
| FHB21 | Tdurum_contig44948_1812 | 7B | 699838421 | 0.000648 | 2.91 |
| FHB23 | Tdurum_contig28176_55 | 7B | 544138607 | 0.000251 | 5.49 |
| FHB23 | *Tdurum_contig102328_129 | 7B | 565742062 | 0.000754 | 3.54 |
| FHB21 | *Tdurum_contig102328_129 | 7B | 565742062 | 0.000138 | 3.44 |
| FHBm | *Tdurum_contig102328_129 | 7B | 565742062 | 0.000491 | 2.83 |
| FHB21 | Tdurum_contig10036_474 | 1A | 556304788 | 0.000521 | 2.61 |
| FHB21 | RAC875_rep_c111159_57 | 7A | 700808659 | 0.000859 | 2.81 |
| FHBm | RAC875_c60218_63 | 4A | 217653852 | 0.000257 | 2.05 |
| FHB23 | RAC875_c32611_347 | 4A | 669816688 | 0.000427 | 4.46 |
| FHB21 | RAC875_c2456_849 | 4B | 562356529 | 0.000409 | 4.25 |
| FHB23 | *Ra_c58315_265 | 1A | 473637106 | 0.000242 | 6.05 |
| FHBm | *Ra_c58315_265 | 1A | 473637106 | 0.000152 | 4.99 |
| FHB21 | Kukri_c9683_723 | 7A | 700702592 | 0.000537 | 2.97 |
| FHB23 | *Ku_c20136_198 | 7B | 609385718 | 0.000434 | 3.92 |
| FHB21 | *Ku_c20136_198 | 7B | 609385718 | 0.000458 | 3.34 |
| FHBm | *Ku_c20136_198 | 7B | 609385718 | 0.000618 | 2.96 |
| FHB23 | Excalibur_rep_c111660_89 | 7A | 585065925 | 4.65 × 10−5 | 6.23 |
| FHBm | Excalibur_c59894_97 | 1A | 463813596 | 0.000928 | 3.71 |
| FHB23 | BS00059062_51 | 7B | 545991407 | 4.65 × 10−5 | 6.23 |
| FHB23 | BS00059061_51 | 7B | 545991505 | 0.000251 | 5.49 |
| FHB21 | BobWhite_c14736_188 | 7B | 699375055 | 0.000492 | 3.02 |
| FHBm | AX-94793903 | 1D | 291636258 | 0.00022 | 2.11 |
| FHB23 | AX-94639168 | 2B | 107117765 | 8.98 × 10−5 | 4.11 |
| Genotypes | Ra_c58315_265 | Ku_c20136_198 | Tdurum_contig102328 | FHBindex (Mean) |
|---|---|---|---|---|
| Allele (A/G) | Allele (C/T) | Allele (G/T) | ||
| CIM-29 | A | C | G | 7.93 |
| CIM-39 | A | C | G | 7.26 |
| CIM-9 | A | C | G | 8.80 |
| CIM-3 | A | C | G | 8.94 |
| SNP | Chr | Functional Annotation |
|---|---|---|
| Tdurum_contig10036_474 | 1A | PROTEASOME NON-ATPASE REGULATORY SUBUNIT 5-RELATED |
| Ra_c58315_265 | 1A | A/G-SPECIFIC ADENINE GLYCOSYLASE/ENDONUCLEASE III |
| Excalibur_c59894_97 | 1A | THYMIDINE KINASE 2 |
| AX-94793903 | 1D | Defensin and Defensin-like DEFL family |
| AX-94639168 | 2B | Unknown |
| RAC875_c60218_63 | 4A | F-box and tubby domain-containing protein, |
| RAC875_c32611_347 | 4A | C-terminal processing peptidase/Tsp protease |
| RAC875_c2456_849 | 4B | Exocyst complex subunit Sec15-like |
| wsnp_Ex_c11055_17927668 | 5D | COP1-INTERACTING PROTEIN-LIKE PROTEIN |
| RAC875_rep_c111159_57 | 7A | histone acetyltransferase of the TAFII250 family 2 |
| Kukri_c9683_723 | 7A | Transcription initiation factor TFIID subunit 1 |
| Ku_c20136_198 | 7B | Putative transmembrane protein cmp44E |
| Excalibur_rep_c111660_89 | 7A | AN1-TYPE ZINC FINGER PROTEIN |
| Tdurum_contig44948_1812 | 7B | transcription initiation factor TFIID subunit 1 |
| Tdurum_contig28176_55 | 7B | AN1-TYPE ZINC FINGER PROTEIN |
| Tdurum_contig102328_129 | 7B | Cysteine-Rich repeat protein |
| BS00059062_51 | 7B | Unknown |
| BS00059061_51 | 7B | Unknown |
| BobWhite_c14736_188 | 7B | Methyl-CpG binding domain (MBD) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mandal, R.; He, X.; Singh, G.P.; Kabir, M.R.; Joshi, A.K.; Singh, P.K. Evaluation of Fusarium Head Blight Resistance Through a Genome-Wide Association Study in CIMMYT and South Asian Wheat Germplasm. Pathogens 2025, 14, 490. https://doi.org/10.3390/pathogens14050490
Mandal R, He X, Singh GP, Kabir MR, Joshi AK, Singh PK. Evaluation of Fusarium Head Blight Resistance Through a Genome-Wide Association Study in CIMMYT and South Asian Wheat Germplasm. Pathogens. 2025; 14(5):490. https://doi.org/10.3390/pathogens14050490
Chicago/Turabian StyleMandal, Rupsanatan, Xinyao He, Gyanendra Pratap Singh, Muhammad Rezaul Kabir, Arun Kumar Joshi, and Pawan Kumar Singh. 2025. "Evaluation of Fusarium Head Blight Resistance Through a Genome-Wide Association Study in CIMMYT and South Asian Wheat Germplasm" Pathogens 14, no. 5: 490. https://doi.org/10.3390/pathogens14050490
APA StyleMandal, R., He, X., Singh, G. P., Kabir, M. R., Joshi, A. K., & Singh, P. K. (2025). Evaluation of Fusarium Head Blight Resistance Through a Genome-Wide Association Study in CIMMYT and South Asian Wheat Germplasm. Pathogens, 14(5), 490. https://doi.org/10.3390/pathogens14050490