
Genomic and Antimicrobial Resistance Analysis of an ST25 Streptococcus suis Strain Isolated from a Human in Zhejiang Province, China

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Strains
2.2. Case Information
2.3. Reagents, Instruments, and Strain Identification
2.4. Antimicrobial Susceptibility Testing (AST)
2.5. Genome Sequencing and Assembly
2.6. Bioinformatic Analysis
3. Results
3.1. Strain Identification
3.2. Antimicrobial Susceptibility Testing (AST)
3.3. Comparison of Genomic Sequences
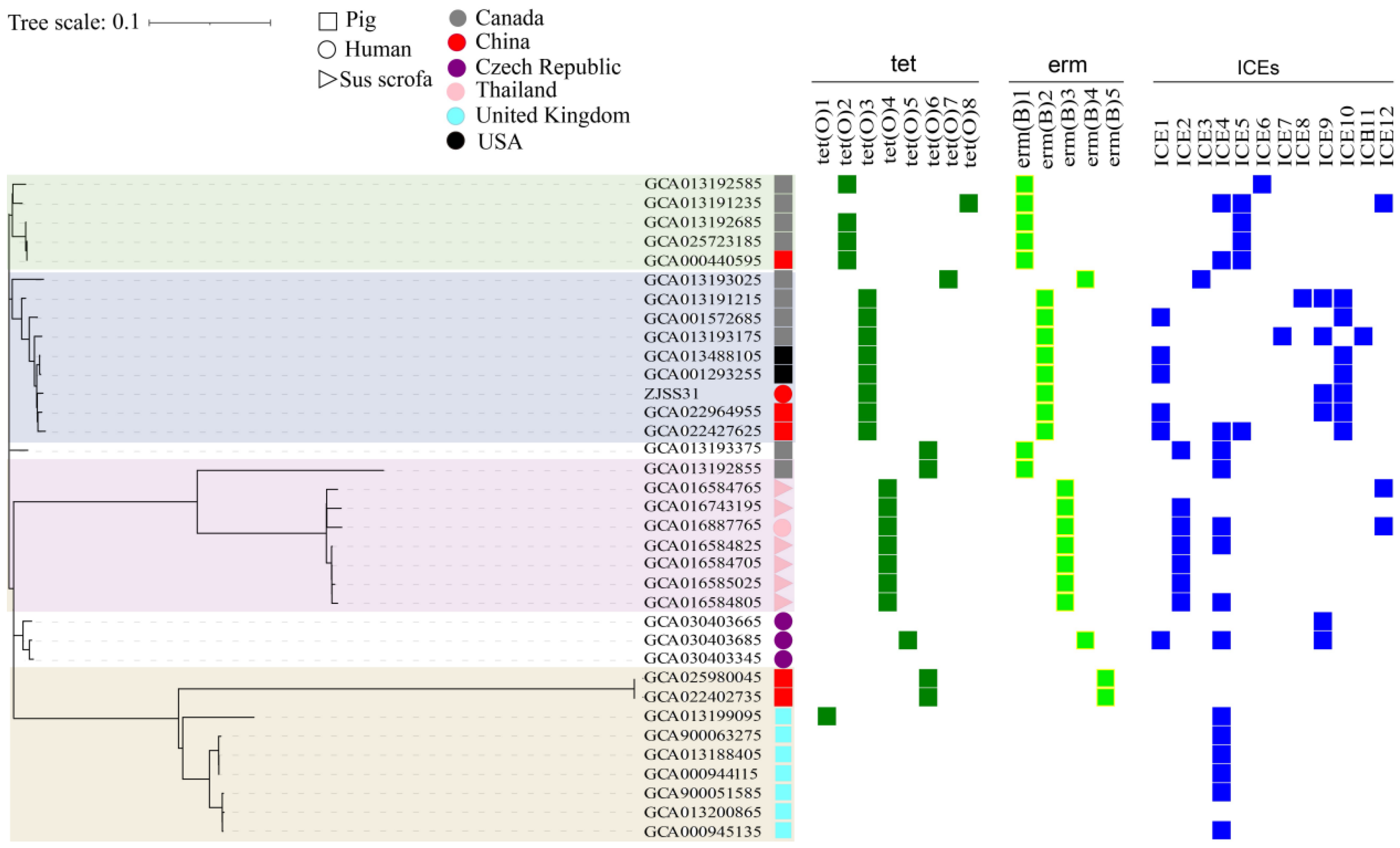
3.4. Antibiotic-Resistance Genes, ICEs, and Phylogeny
4. Discussion
5. Limitations of the Study
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tharavichitkul, P.; Wongsawan, K.; Takenami, N.; Pruksakorn, S.; Fongcom, A.; Gottschalk, M.; Khanthawa, B.; Supajatura, V.; Takai, S. Correlation between PFGE Groups and mrp/epf/sly Genotypes of Human Streptococcus suis Serotype 2 in Northern Thailand. J. Pathog. 2014, 2014, 350416. [Google Scholar] [CrossRef]
- Yongkiettrakul, S.; Maneerat, K.; Arechanajan, B.; Malila, Y.; Srimanote, P.; Gottschalk, M.; Visessanguan, W. Antimicrobial susceptibility of Streptococcus suis isolated from diseased pigs, asymptomatic pigs, and human patients in Thailand. BMC Vet. Res. 2019, 15, 5. [Google Scholar] [CrossRef]
- Zheng, H.; Ji, S.; Liu, Z.; Lan, R.; Huang, Y.; Bai, X.; Gottschalk, M.; Xu, J. Eight Novel Capsular Polysaccharide Synthesis Gene Loci Identified in Nontypeable Streptococcus suis Isolates. Appl. Environ. Microbiol. 2015, 81, 4111–4119. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Qiu, X.; Roy, D.; Segura, M.; Du, P.; Xu, J.; Gottschalk, M. Genotyping and investigating capsular polysaccharide synthesis gene loci of non-serotypeable Streptococcus suis isolated from diseased pigs in Canada. Vet. Res. 2017, 48, 10. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Zhang, H.; Wu, Z.; Wang, S.; Cao, M.; Hu, D.; Wang, C. Streptococcus suis infection: An emerging/reemerging challenge of bacterial infectious diseases? Virulence 2014, 5, 477–497. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.; Zhu, X.; Jing, H.; Du, H.; Segura, M.; Zheng, H.; Kan, B.; Wang, L.; Bai, X.; Zhou, Y.; et al. Streptococcus suis sequence type 7 outbreak, Sichuan, China. Emerg. Infect. Dis. 2006, 12, 1203–1208. [Google Scholar] [CrossRef]
- Tang, J.; Wang, C.; Feng, Y.; Yang, W.; Song, H.; Chen, Z.; Yu, H.; Pan, X.; Zhou, X.; Wang, H.; et al. Streptococcal toxic shock syndrome caused by Streptococcus suis serotype 2. PLoS Med. 2006, 3, e151. [Google Scholar] [CrossRef]
- Huang, W.; Wang, M.; Hao, H.; Yang, R.; Xie, J.; Su, J.; Lin, M.; Cui, Y.; Jiang, Y. Genomic epidemiological investigation of a Streptococcus suis outbreak in Guangxi, China, 2016. Infect. Genet. Evol. 2019, 68, 249–252. [Google Scholar] [CrossRef]
- van Leengoed, L.A.; Vecht, U.; Verheyen, E.R. Streptococcus suis type 2 infections in pigs in the Netherlands (Part two). Vet. Q. 1987, 9, 111–117. [Google Scholar] [CrossRef]
- Schultsz, C.; Jansen, E.; Keijzers, W.; Rothkamp, A.; Duim, B.; Wagenaar, J.A.; van der Ende, A. Differences in the population structure of invasive Streptococcus suis strains isolated from pigs and from humans in The Netherlands. PLoS ONE 2012, 7, e33854. [Google Scholar] [CrossRef]
- Wang, M.; Du, P.; Wang, J.; Lan, R.; Huang, J.; Luo, M.; Jiang, Y.; Zeng, J.; Quan, Y.; Shi, Z.; et al. Genomic Epidemiology of Streptococcus suis Sequence Type 7 Sporadic Infections in the Guangxi Zhuang Autonomous Region of China. Pathogens 2019, 8, 187. [Google Scholar] [CrossRef] [PubMed]
- Cucco, L.; Paniccià, M.; Massacci, F.R.; Morelli, A.; Ancora, M.; Mangone, I.; Di Pasquale, A.; Luppi, A.; Vio, D.; Cammà, C.; et al. New Sequence Types and Antimicrobial Drug-Resistant Strains of Streptococcus suis in Diseased Pigs, Italy, 2017–2019. Emerg. Infect. Dis. 2022, 28, 139–147. [Google Scholar] [CrossRef]
- Zhu, S.R.; Chen, J.C.; Wu, Z.Y.; Zhang, Y.L.; Fang, L.; Zhang, Y.J. Molecular characteristics of Streptococcus suis type 2 from humans in Zhejiang Province. Chin. J. Infect. Dis. 2022, 40, 483–489. [Google Scholar] [CrossRef]
- April, A.A.; Gottschalk, M.; Rossow, S.; Rendahl, A.; Gebhart, C.; Marthaler, D.G. Serotype and Genotype (Multilocus Sequence Type) of Streptococcus suis Isolates from the United States Serve as Predictors of Pathotype. J. Clin. Microbiol. 2019, 57, e00377-19. [Google Scholar] [CrossRef]
- Athey, T.B.; Teatero, S.; Takamatsu, D.; Wasserscheid, J.; Dewar, K.; Gottschalk, M.; Fittipaldi, N. Population Structure and Antimicrobial Resistance Profiles of Streptococcus suis Serotype 2 Sequence Type 25 Strains. PLoS ONE 2016, 11, e0150908. [Google Scholar] [CrossRef]
- Zhu, S.R.; Zhang, Z.; Yao, P.P.; Yang, Y.; Fang, L.; Zhang, Y.J. Identification and characterization of the virulence of Streptococcus suis from human patients in Zhejiang Province, China. Chin. J. Zoonoses. 2021, 37, 808–814. [Google Scholar]
- M100-S28E; Performance Standards for Antimicrobial Susceptibility Testing, 34th Informational Supplement. Clinical Laboratory Standards Institute: Wayne, PA, USA, 2024.
- Hu, Y.; Fu, S.; Zou, G.; Kerdsin, A.; Chen, X.; Dong, X.; Teng, L.; Li, J. Genome analysis provides insight into hyper-virulence of Streptococcus suis LSM178, a human strain with a novel sequence type 1005. Sci. Rep. 2021, 11, 23919. [Google Scholar] [CrossRef]
- Nedbalcova, K.; Kucharovicova, I.; Zouharova, M.; Matiaskova, K.; Kralova, N.; Brychta, M.; Simek, B.; Pecha, T.; Plodkova, H.; Matiasovic, J. Resistance of Streptococcus suis Isolates from the Czech Republic during 2018–2022. Antibiotics 2022, 11, 1214. [Google Scholar] [CrossRef]
- Bojarska, A.; Molska, E.; Janas, K.; Skoczyńska, A.; Stefaniuk, E.; Hryniewicz, W.; Sadowy, E. Streptococcus suis in invasive human infections in Poland: Clonality and determinants of virulence and antimicrobial resistance. Eur. J. Clin. Microbiol. Infect. Dis. 2016, 35, 917–925. [Google Scholar] [CrossRef] [PubMed]
- Bamphensin, N.; Chopjitt, P.; Hatrongjit, R.; Boueroy, P.; Fittipaldi, N.; Gottschalk, M.; Kerdsin, A. Non-Penicillin-Susceptible Streptococcus suis Isolated from Humans. Pathogens 2021, 10, 1178. [Google Scholar] [CrossRef] [PubMed]
- Gurung, M.; Tamang, M.D.; Moon, D.C.; Kim, S.R.; Jeong, J.H.; Jang, G.C.; Jung, S.C.; Park, Y.H.; Lim, S.K. Molecular basis of resistance to selected antimicrobial agents in the emerging zoonotic pathogen Streptococcus suis. J. Clin. Microbiol. 2015, 53, 2332–2336. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa, T.; Oshima, M.; Yamagishi, J.; Muramatsu, C.; Asai, T. Changes in antimicrobial resistance phenotypes and genotypes in Streptococcus suis strains isolated from pigs in the Tokai area of Japan. J. Vet. Med. Sci. 2020, 82, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Aradanas, M.; Poljak, Z.; Fittipaldi, N.; Ricker, N.; Farzan, A. Serotypes, virulence-associated factors, and antimicrobial resistance of Streptococcus suis isolates recovered from sick and healthy pigs determined by whole-genome sequencing. Front. Vet. Sci. 2021, 8, 742345. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Tsukagoshi, H.; Sekizuka, T.; Ishikawa, D.; Imai, M.; Fujita, M.; Kuroda, M.; Saruki, N. Next-generation DNA sequencing analysis of two Streptococcus suis ST28 isolates associated with human infective endocarditis and meningitis in Gunma, Japan: A case report. Infect. Dis. 2019, 51, 62–66. [Google Scholar] [CrossRef]
- Raberahona, M.; Rasoanandrasana, S.; Rahajamanana, V.L.; Ranaivo-Rabetokotany, F.; Andriananja, V.; Rakotomalala, F.A.; Randria, M.J.D.; Rakotovao, L.; Marois-Créhan, C.; Tocqueville, V.; et al. Novel Streptococcus suis Sequence Type 834 among Humans, Madagascar. Emerg. Infect. Dis. 2018, 24, 391–392. [Google Scholar] [CrossRef]
- Palmieri, C.; Magi, G.; Mingoia, M.; Bagnarelli, P.; Ripa, S.; Varaldo, P.E.; Facinelli, B. Characterization of a Streptococcus suis tet(O/W/32/O)-carrying element transferable to major streptococcal pathogens. Antimicrob. Agents Chemother. 2012, 56, 4697–4702. [Google Scholar] [CrossRef]
- Marini, E.; Palmieri, C.; Magi, G.; Facinelli, B. Recombination between Streptococcus suis ICESsu32457 and Streptococcus agalactiae ICESa2603 yields a hybrid ICE transferable to Streptococcus pyogenes. Vet. Microbiol. 2015, 178, 99–104. [Google Scholar] [CrossRef]
- Fittipaldi, N.; Takamatsu, D.; de la Cruz Domínguez-Punaro, M. Mutations in the gene encoding the ancillary pilin subunit of the Streptococcus suis srtF cluster result in pili formed by the major subunit only. PLoS ONE 2010, 5, e8426. [Google Scholar] [CrossRef]
- Okura, M.; Osaki, M.; Fittipaldi, N.; Gottschalk, M.; Sekizaki, T.; Takamatsu, D. The minor pilin subunit Sgp2 is necessary for assembly of the pilus encoded by the srtG cluster of Streptococcus suis. J. Bacteriol. 2011, 193, 822–831. [Google Scholar] [CrossRef]
- Guo, G.; Du, D.; Yu, Y.; Zhang, Y.; Qian, Y.; Zhang, W. Pan-genome analysis of Streptococcus suis serotype 2 revealed genomic diversity among strains of different virulence. Transbound. Emerg. Dis. 2021, 68, 637–647. [Google Scholar] [CrossRef]
- Uruén, C.; Fernandez, A.; Arnal, J.L.; Del Pozo, M.; Amoribieta, M.C.; de Blas, I.; Jurado, P.; Calvo, J.H.; Gottschalk, M.; González-Vázquez, L.D.; et al. Genomic and phenotypic analysis of invasive Streptococcus suis isolated in Spain reveals genetic diversification and associated virulence traits. Vet. Res. 2024, 55, 11. [Google Scholar] [CrossRef]
- Fittipaldi, N.; Xu, J.; Lacouture, S.; Tharavichitkul, P.; Osaki, M.; Sekizaki, T.; Takamatsu, D.; Gottschalk, M. Lineage and virulence of Streptococcus suis serotype 2 isolates from North America. Emerg. Infect. Dis. 2011, 17, 2239–2244. [Google Scholar] [CrossRef]
- Chu, Y.W.; Cheung, T.K.; Chu, M.Y.; Tsang, V.Y.; Fung, J.T.; Kam, K.M.; Lo, J.Y. Resistance to tetracycline, erythromycin and clindamycin in Streptococcus suis serotype 2 in Hong Kong. Int. J. Antimicrob. Agents. 2009, 34, 181–182. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.; Chen, Z.; Li, F.; Hu, Q.; Xu, L.; Duan, X.; Wu, H.; Xu, S.; Chen, Q.; Wu, S.; et al. Epidemiological and genomic analyses of human isolates of Streptococcus suis between 2005 and 2021 in Shenzhen, China. Front. Microbiol. 2023, 14, 1118056. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Accession No. | Strain Name | Submission Time | Region | Host | Sequence Type | Serotype |
|---|---|---|---|---|---|---|
| GCA_022964955.1 | TJS75 | 2015 | Tianjin, China | Pig | ST25 | 2 |
| GCA_025980045.1 | WUSS030 | 2017 | Jiangsu, China | Pig | ST25 | 8 |
| GCA_022427625.1 | LSSP102 | 2017 | Hubei, China | Wild pig | ST25 | 2 |
| GCA_022402735.1 | WUSS030 | 2022 | Nanjing, China | Pig | ST25 | 8 |
| GCA_000440595.1 | 89-1591 | 2012 | Beijing, China | Pig | ST25 | 2 |
| GCA_001572685.1 | NSUI060 | 2008 | Canada | Pig | ST25 | 2 |
| GCA_013192685.1 | 89-1591 | 1989 | Canada | Pig | ST25 | 2 |
| GCA_013192855.1 | 1636820 | 2014 | Canada | Pig | ST25 | 2 |
| GCA_013193025.1 | 1602951 | 2014 | Canada | Pig | ST25 | 2 |
| GCA_013191235.1 | 89-6891-2 | 1989 | Canada | Pig | ST25 | 2 |
| GCA_013192585.1 | 89-5046 | 1989 | Canada | Pig | ST25 | 2 |
| GCA_013191215.1 | 90-2741-7 | 1990 | Canada | Pig | ST25 | 2 |
| GCA_013193175.1 | 1667796 | 2014 | Canada | Pig | ST25 | 2 |
| GCA_013193375.1 | 1666725 | 2014 | Canada | Pig | ST25 | 2 |
| GCA_025723185.1 | 89-1591 | 1989 | Canada | Pig | ST25 | 2 |
| GCA_001293255.1 | ISU2514 | 2014 | USA | Wild pig | ST25 | 2 |
| GCA_013488105.1 | ISU2514 | 2014 | USA | Wild pig | ST25 | 2 |
| GCA_030403345.1 | 150002 | 2004 | Czech | Human | ST25 | 2 |
| GCA_030403665.1 | 148897 | 2003 | Czech | Human | ST25 | 2 |
| GCA_030403685.1 | 143741 | 2001 | Czech | Human | ST25 | 2 |
| GCA_900063275.1 | SS1053 | 2012 | UK | Pig | ST25 | 2 |
| GCA_000944115.1 | S15T | 2010 | UK | Pig | ST25 | 2 |
| GCA_900051585.1 | SS1064 | 2012 | UK | Pig | ST25 | 2 |
| GCA_000945135.1 | S16C | 2010 | UK | Pig | ST25 | 2 |
| GCA_013199095.1 | TMW_SS009 | 2013 | UK | Pig | ST25 | 2 |
| GCA_013188405.1 | SS1053 | 2010 | UK | Pig | ST25 | 2 |
| GCA_013200865.1 | SS1064 | 2010 | UK | Pig | ST25 | 2 |
| GCA_016743195.1 | DP_SS29 | 2012 | Thailand | Wild pig | ST25 | 2 |
| GCA_016584705.1 | MNCM02 | 2000 | Thailand | Wild pig | ST25 | 2 |
| GCA_016585025.1 | MNCM34 | 2002 | Thailand | Wild pig | ST25 | 2 |
| GCA_016584825.1 | MNCM13 | 2001 | Thailand | Wild pig | ST25 | 2 |
| GCA_016584765.1 | MNCM03 | 2000 | Thailand | Wild pig | ST25 | 2 |
| GCA_016887765.1 | ID30190 | 2009 | Thailand | Human | ST25 | 2 |
| GCA_016584805.1 | MNCM11 | 2001 | Thailand | Wild pig | ST25 | 2 |
| Antibiotic Type | Antimicrobial Agent | MIC (mg/L)/(R/I/S) | |
|---|---|---|---|
| β-lactams | Penicillins | Penicillin | ≤0.03 (S) |
| Cephalosporins | Cefotaxime | ≤0.12 (S) | |
| Ceftriaxone | ≤0.12 (S) | ||
| Cefepime | ≤0.5 (S) | ||
| Carbapenems | Meropenem | ≤0.25 (S) | |
| Ertapenem | ≤0.5(S) | ||
| Tetracycline | Tetracycline | >8 (R) | |
| Chloramphenicol | Chloramphenicol | =4 (S) | |
| Macrolides | Azithromycin | >2 (R) | |
| Erythromycin | >2 (R) | ||
| Lincosamides | Clindamycin | >1 (R) | |
| Cyclic lipopeptide | Daptomycin | =0.12 (S) | |
| Quinolones | Levofloxacin | ≤0.5 (S) | |
| Oxazolidone | Linezolid | =1 (S) | |
| Glycopeptides | Vancomycin | ≤0.5 (S) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, S.; Wu, X.; Yao, W.; Wu, Z.; Wang, L.; Yang, Z.; Wu, B.; Zhang, Y. Genomic and Antimicrobial Resistance Analysis of an ST25 Streptococcus suis Strain Isolated from a Human in Zhejiang Province, China. Pathogens 2025, 14, 742. https://doi.org/10.3390/pathogens14080742
Zhu S, Wu X, Yao W, Wu Z, Wang L, Yang Z, Wu B, Zhang Y. Genomic and Antimicrobial Resistance Analysis of an ST25 Streptococcus suis Strain Isolated from a Human in Zhejiang Province, China. Pathogens. 2025; 14(8):742. https://doi.org/10.3390/pathogens14080742
Chicago/Turabian StyleZhu, Shuirong, Xiaofang Wu, Wenwu Yao, Zhuoying Wu, Lingbo Wang, Zhangnv Yang, Beibei Wu, and Yanjun Zhang. 2025. "Genomic and Antimicrobial Resistance Analysis of an ST25 Streptococcus suis Strain Isolated from a Human in Zhejiang Province, China" Pathogens 14, no. 8: 742. https://doi.org/10.3390/pathogens14080742
APA StyleZhu, S., Wu, X., Yao, W., Wu, Z., Wang, L., Yang, Z., Wu, B., & Zhang, Y. (2025). Genomic and Antimicrobial Resistance Analysis of an ST25 Streptococcus suis Strain Isolated from a Human in Zhejiang Province, China. Pathogens, 14(8), 742. https://doi.org/10.3390/pathogens14080742