

Expanding the Antiviral Spectrum of Scorpion-Derived Peptides Against Toscana Virus and Schmallenberg Virus

 , , , , , ,
, , , , , ,  , , ,
, , ,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Peptide Synthesis and Characterization
2.3. Cell and Virus Growth
2.4. Cytotoxicity
2.5. Antiviral Activity
2.5.1. Plaque Assay
2.5.2. Tissue Culture Infective Dose Assay
2.6. Real-Time Quantitative PCR (qPCR)
2.7. Calculation of Selectivity Index
2.8. Statistics
3. Results
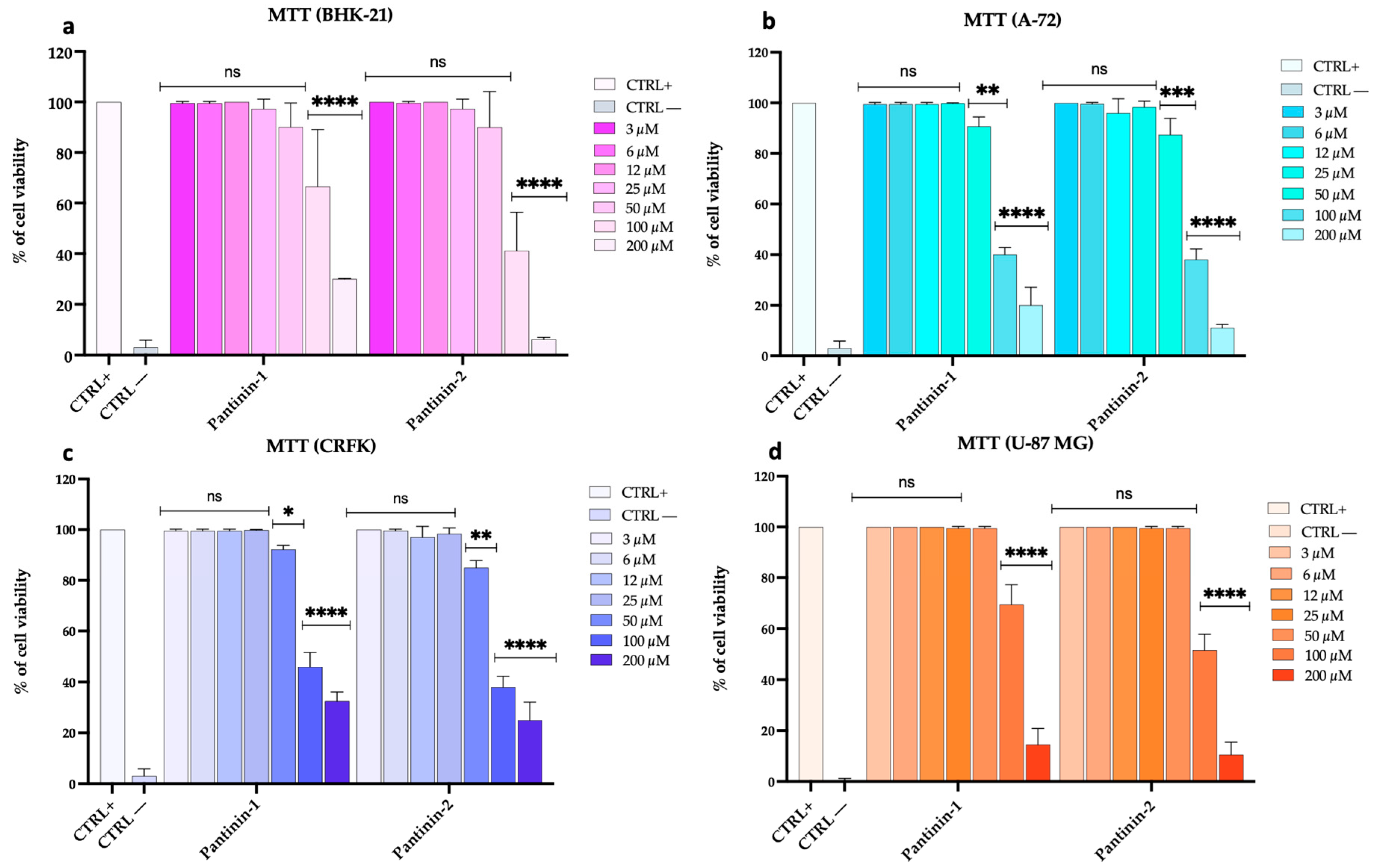
3.1. Pantinins’ Cytotoxicity on Different Cellular Models
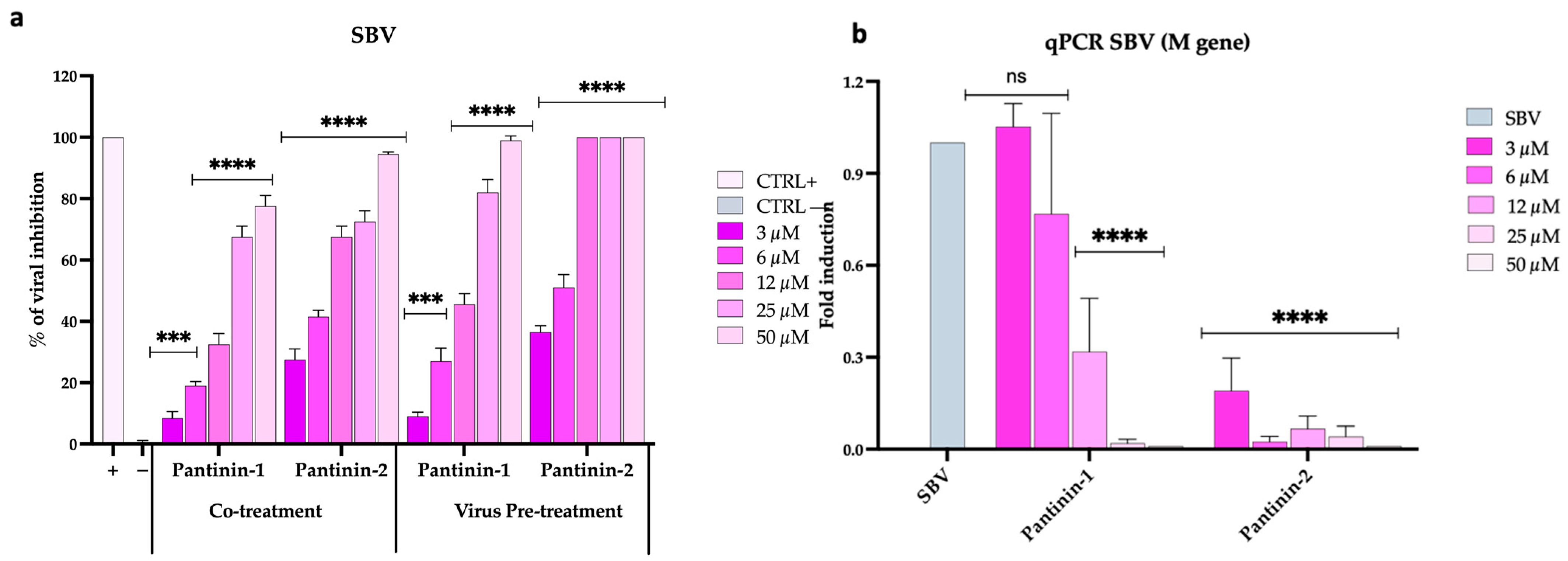
3.2. Inhibitory Effects of Pantinins on SBV Replication
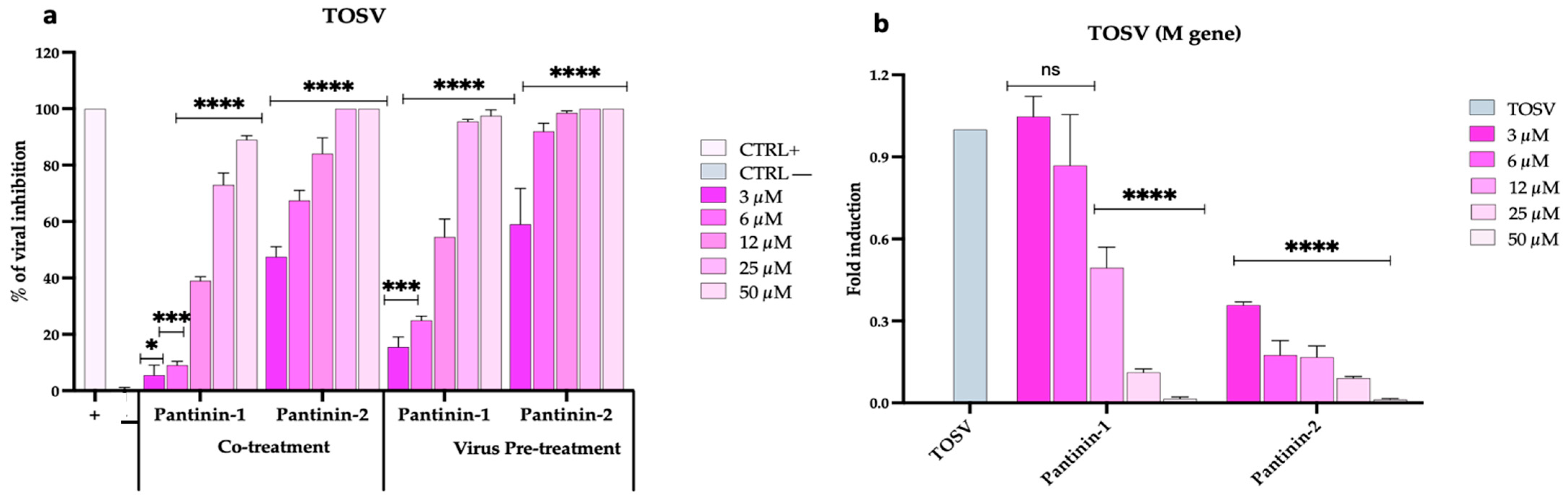
3.3. Inhibitory Effects of Pantinins on TOSV Replication
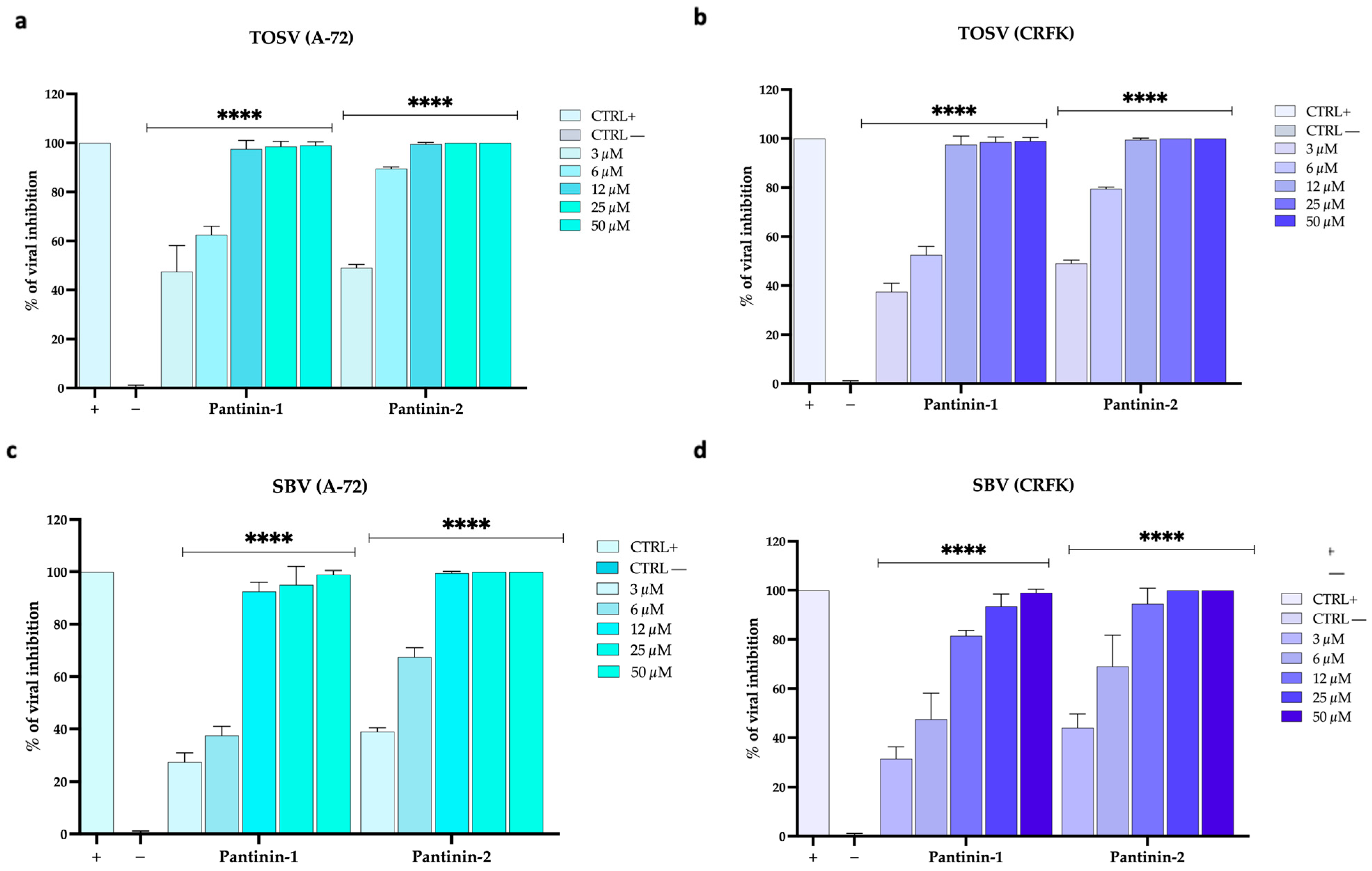
3.4. Inhibitory Effects of Pantinins Against SBV and TOSV Infection in Animal Cell Models
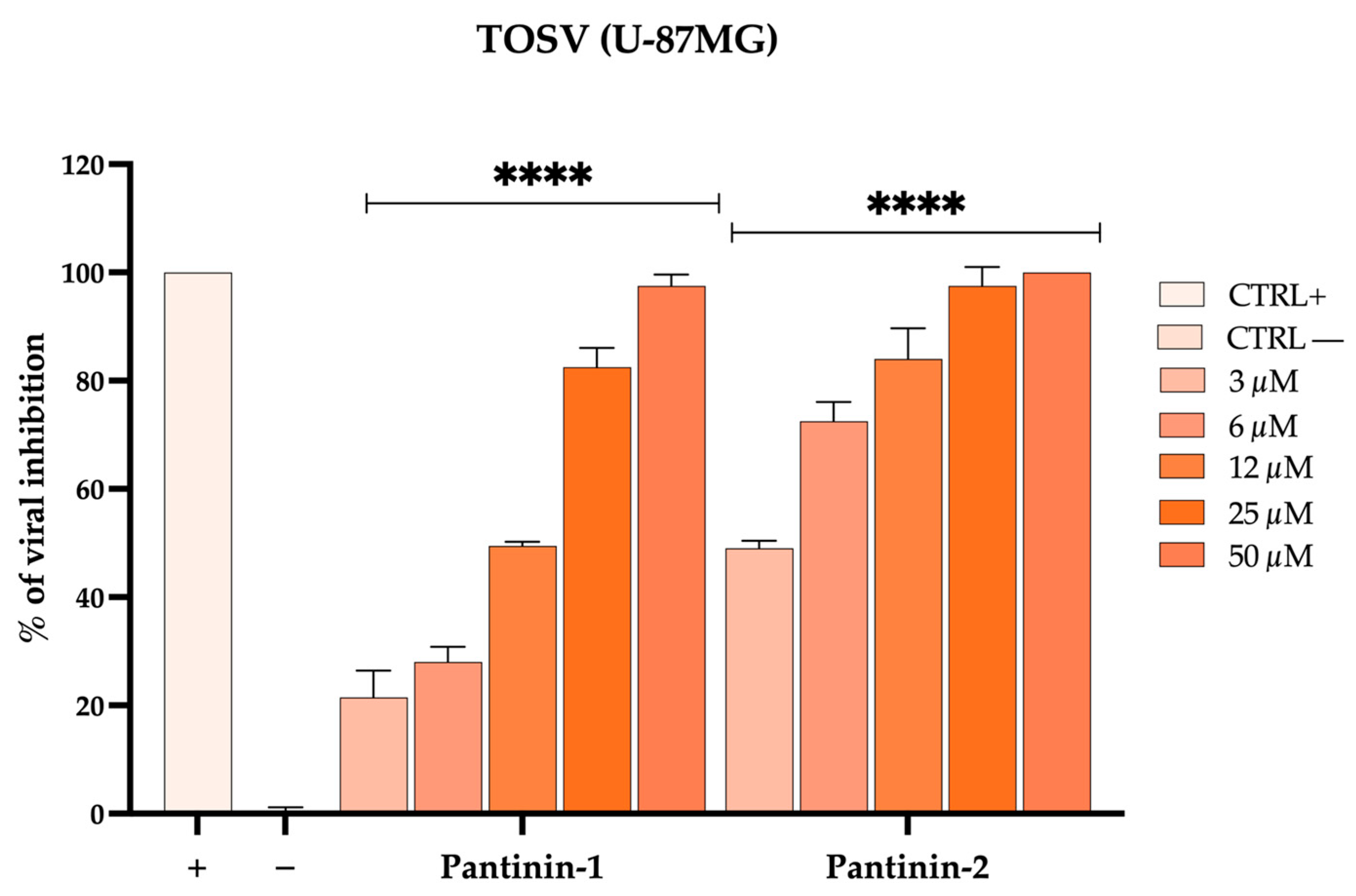
3.5. Pantinins Inhibit TOSV Infection in U-87 MG Neuronal Cells
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| TOSV | Toscana virus |
| SBV | Schmallenberg virus |
| HDPs | Host defense peptides |
| IC50 | Half-maximal inhibitory concentration |
| MTT | Methylthiazolyldiphenyl-tetrazolium bromide |
| DMSO | Dimethyl sulfoxide |
| MOI | Multiplicity of infection |
| DMEM | Dulbecco’s Modified Eagle Medium |
| FBS | Fetal bovine serum |
| CMC | Carboxymethylcellulose |
| CV | Crystal violet |
| TCID50 | Tissue culture infective dose |
| CPE | Cytopathic effect |
| qPCR | Real-time quantitative PCR |
| Ct | Cycle threshold |
| GAPDH | Glyceraldehyde 3-phosphate dehydrogenase |
| SI | Selectivity index |
References
- Baghdadi, G.A.; Aakef, I.R.; Mahdi, S.G.; Khaleel, R.I. Crimean-Congo haemorrhagic fever in Iraq. East. Mediterr. Health J. 2024, 30, 570–576. [Google Scholar] [CrossRef] [PubMed]
- Ze-Ze, L.; Nunes, C.; Sousa, M.; de Sousa, R.; Gomes, C.; Santos, A.S.; Alexandre, R.T.; Amaro, F.; Loza, T.; Blanco, M.; et al. Fatal Case of Crimean-Congo Hemorrhagic Fever, Portugal, 2024. Emerg. Infect. Dis. 2025, 31, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Hamdard, E.; Zahir, A.; Karwand, B.; Nazari, Z.U.; Shi, F. Silent public threat: Crimean-Congo hemorrhagic fever outbreak spikes during Eid-al-Adha in Afghanistan (Reported cases to National Surveillance System, 2015–2024). J. Infect. Public Health 2025, 18, 102591. [Google Scholar] [CrossRef] [PubMed]
- Youssouf, H.; Subiros, M.; Dennetiere, G.; Collet, L.; Dommergues, L.; Pauvert, A.; Rabarison, P.; Vauloup-Fellous, C.; Le Godais, G.; Jaffar-Bandjee, M.C.; et al. Rift Valley Fever Outbreak, Mayotte, France, 2018-2019. Emerg. Infect. Dis. 2020, 26, 769–772. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.; Ali, Y.; Elduma, A.; Eldigail, M.H.; Mhmoud, R.A.; Mohamed, N.S.; Ksiazek, T.G.; Dietrich, I.; Weaver, S.C. Unique Outbreak of Rift Valley Fever in Sudan, 2019. Emerg. Infect. Dis. 2020, 26, 3030–3033. [Google Scholar] [CrossRef] [PubMed]
- Sah, R.; Srivastava, S.; Mehta, R.; Khan, S.R.; Kumar, S.; Satpathy, P.; Mohanty, A.; Ferraz, C.; Feehan, J.; Apostolopoulos, V.; et al. Oropouche fever fatalities and vertical transmission in South America: Implications of a potential new mode of transmission. Lancet Reg. Health Am. 2024, 38, 100896. [Google Scholar] [CrossRef] [PubMed]
- Okesanya, O.J.; Amisu, B.O.; Adigun, O.A.; Ahmed, M.M.; Agboola, A.O.; Kab, T.; Eshun, G.; Ukoaka, B.M.; Oso, T.A.; Ogaya, J.B.; et al. Addressing the emerging threat of Oropouche virus: Implications and public health responses for healthcare systems. Trop. Dis. Travel. Med. Vaccines 2025, 11, 1. [Google Scholar] [CrossRef] [PubMed]
- Castilletti, C.; Mori, A.; Matucci, A.; Ronzoni, N.; Van Duffel, L.; Rossini, G.; Sponga, P.; D’Errico, M.L.; Rodari, P.; Cristini, F.; et al. Oropouche fever cases diagnosed in Italy in two epidemiologically non-related travellers from Cuba, late May to early June 2024. Eurosurveillance 2024, 29, 2400362. [Google Scholar] [CrossRef] [PubMed]
- Scachetti, G.C.; Forato, J.; Claro, I.M.; Hua, X.; Salgado, B.B.; Vieira, A.; Simeoni, C.L.; Barbosa, A.R.C.; Rosa, I.L.; de Souza, G.F.; et al. Re-emergence of Oropouche virus between 2023 and 2024 in Brazil: An observational epidemiological study. Lancet Infect. Dis. 2025, 25, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Ozkale, Y.; Ozkale, M.; Kiper, P.; Cetinkaya, B.; Erol, I. Sadfly fever: Two case reports. Turk. Pediatri. Ars. 2016, 51, 110–113. [Google Scholar] [CrossRef] [PubMed]
- Tchouassi, D.P.; Marklewitz, M.; Chepkorir, E.; Zirkel, F.; Agha, S.B.; Tigoi, C.C.; Koskei, E.; Drosten, C.; Borgemeister, C.; Torto, B.; et al. Sand Fly-Associated Phlebovirus with Evidence of Neutralizing Antibodies in Humans, Kenya. Emerg. Infect. Dis. 2019, 25, 681–690. [Google Scholar] [CrossRef] [PubMed]
- Hong, X.G.; Zhang, M.Q.; Tang, F.; Song, S.H.; Wang, J.Y.; Hu, Z.Y.; Liu, L.M.; Zhang, X.A.; Sun, Y.; Fang, L.Q.; et al. Epidemiology and Ecology of Toscana Virus Infection and Its Global Risk Distribution. Viruses 2024, 17, 15. [Google Scholar] [CrossRef] [PubMed]
- Ayhan, N.; Eldin, C.; Charrel, R. Toscana virus: A comprehensive review of 1381 cases showing an emerging threat in the Mediterranean regions. J. Infect. 2025, 90, 106415. [Google Scholar] [CrossRef] [PubMed]
- Fotakis, E.A.; Di Maggio, E.; Del Manso, M.; Mateo-Urdiales, A.; Petrone, D.; Fabiani, M.; Perego, G.; Bella, A.; Bongiorno, G.; Bernardini, I.; et al. Human neuroinvasive Toscana virus infections in Italy from 2016 to 2023: Increased incidence in 2022 and 2023. Eurosurveillance 2025, 30, 2400203. [Google Scholar] [CrossRef] [PubMed]
- Tomljenovic, M.; Lakoseljac, D.; Knezevic, L.; Batista, M.; Vilibic-Cavlek, T.; Kaic, B.; Hansen, L.; Rode, O.D. Spread of Puumala Hantavirus to New Areas in a Large Croatian Outbreak of Hemorrhagic Fever with Renal Syndrome, 2021. Vector Borne Zoonotic Dis. 2024, 24, 773–783. [Google Scholar] [CrossRef] [PubMed]
- Mir, S. Hantavirus Induced Kidney Disease. Front. Med. 2021, 8, 795340. [Google Scholar] [CrossRef] [PubMed]
- Wernike, K.; Beer, M. Schmallenberg Virus: A Novel Virus of Veterinary Importance. Adv. Virus Res. 2017, 99, 39–60. [Google Scholar] [CrossRef] [PubMed]
- Varela, M.; Schnettler, E.; Caporale, M.; Murgia, C.; Barry, G.; McFarlane, M.; McGregor, E.; Piras, I.M.; Shaw, A.; Lamm, C.; et al. Schmallenberg virus pathogenesis, tropism and interaction with the innate immune system of the host. PLoS Pathog. 2013, 9, e1003133. [Google Scholar] [CrossRef] [PubMed]
- Dagnaw, M.; Solomon, A.; Dagnew, B. Serological prevalence of the Schmallenberg virus in domestic and wild hosts worldwide: A systematic review and meta-analysis. Front. Vet. Sci. 2024, 11, 1371495. [Google Scholar] [CrossRef] [PubMed]
- Pinto, G.O.A.; Ferreira, M.N.S.; Oliveira, P.R.F.; Samico-Fernandes, E.F.T.; Melo, R.P.B.; Junior, J.W.P.; Peixoto, R.M.; Mota, R.A. Investigation of Schmallenberg virus and update on the serological status of Toxoplasma gondii in goat herds in the semiarid region of Pernambuco state, Brazil. Vet. Parasitol. Reg. Stud. Rep. 2025, 62, 101284. [Google Scholar] [CrossRef] [PubMed]
- Waret-Szkuta, A.; Alarcon, P.; Hasler, B.; Rushton, J.; Corbiere, F.; Raboisson, D. Economic assessment of an emerging disease: The case of Schmallenberg virus in France. Rev. Sci. Tech. 2017, 36, 265–277. [Google Scholar] [CrossRef] [PubMed]
- Dincer, E.; Gargari, S.; Ozkul, A.; Ergunay, K. Potential animal reservoirs of Toscana virus and coinfections with Leishmania infantum in Turkey. Am. J. Trop. Med. Hyg. 2015, 92, 690–697. [Google Scholar] [CrossRef] [PubMed]
- Munoz, C.; Ayhan, N.; Ortuno, M.; Ortiz, J.; Gould, E.A.; Maia, C.; Berriatua, E.; Charrel, R.N. Experimental Infection of Dogs with Toscana Virus and Sandfly Fever Sicilian Virus to Determine Their Potential as Possible Vertebrate Hosts. Microorganisms 2020, 8, 596. [Google Scholar] [CrossRef] [PubMed]
- Hubálek, Z.; Rudolf, I. Vertebrates as Hosts and Reservoirs of Zoonotic Microbial Agents. Microb. Zoonoses Sapronoses 2010, 10, 83–128. [Google Scholar] [CrossRef]
- Adnyana, I.M.; Utomo, B.; Eljatin, D.S.; Sudaryati, N.L. One Health approach and zoonotic diseases in Indonesia: Urgency of implementation and challenges. Narra J. 2023, 3, e257. [Google Scholar] [CrossRef] [PubMed]
- Heitmann, A.; Gusmag, F.; Rathjens, M.G.; Maurer, M.; Frankze, K.; Schicht, S.; Jansen, S.; Schmidt-Chanasit, J.; Jung, K.; Becker, S.C. Mammals Preferred: Reassortment of Batai and Bunyamwera orthobunyavirus Occurs in Mammalian but Not Insect Cells. Viruses 2021, 13, 1702. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Hu, S.; Liu, X.; Yang, J.; Liu, D.; Wu, L.; Wang, H.; Hu, Z.; Deng, F.; Shen, S. Migration, recombination, and reassortment are involved in the evolution of severe fever with thrombocytopenia syndrome bunyavirus. Infect. Genet. Evol. 2017, 47, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Klempa, B. Reassortment events in the evolution of hantaviruses. Virus Genes 2018, 54, 638–646. [Google Scholar] [CrossRef] [PubMed]
- Freire, C.C.; Iamarino, A.; Soumare, P.O.; Faye, O.; Sall, A.A.; Zanotto, P.M. Reassortment and distinct evolutionary dynamics of Rift Valley Fever virus genomic segments. Sci. Rep. 2015, 5, 11353. [Google Scholar] [CrossRef] [PubMed]
- Drayton, M.; Deisinger, J.P.; Ludwig, K.C.; Raheem, N.; Muller, A.; Schneider, T.; Straus, S.K. Host Defense Peptides: Dual Antimicrobial and Immunomodulatory Action. Int. J. Mol. Sci. 2021, 22, 11172. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.C.; Zhou, L.; Shi, W.; Luo, X.; Zhang, L.; Nie, Y.; Wang, J.; Wu, S.; Cao, B.; Cao, H. Three new antimicrobial peptides from the scorpion Pandinus imperator. Peptides 2013, 45, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Keikha, M.; Kamali, H.; Ghazvini, K.; Karbalaei, M. Antimicrobial peptides: Natural or synthetic defense peptides against HBV and HCV infections. Virusdisease 2022, 33, 445–455. [Google Scholar] [CrossRef] [PubMed]
- Xia, Z.; Xie, L.; Li, B.; Lv, X.; Zhang, H.; Cao, Z. Antimicrobial Potential of Scorpion-Venom-Derived Peptides. Molecules 2024, 29, 5080. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Lang, Y.; Ji, Z.; Xia, Z.; Han, Y.; Cheng, Y.; Liu, G.; Sun, F.; Zhao, Y.; Gao, M.; et al. A scorpion venom peptide Ev37 restricts viral late entry by alkalizing acidic organelles. J. Biol. Chem. 2019, 294, 182–194. [Google Scholar] [CrossRef] [PubMed]
- Ji, Z.; Li, F.; Xia, Z.; Guo, X.; Gao, M.; Sun, F.; Cheng, Y.; Wu, Y.; Li, W.; Ali, S.A.; et al. The Scorpion Venom Peptide Smp76 Inhibits Viral Infection by Regulating Type-I Interferon Response. Virol. Sin. 2018, 33, 545–556. [Google Scholar] [CrossRef] [PubMed]
- Lima, W.G.; Maia, C.Q.; de Carvalho, T.S.; Leite, G.O.; Brito, J.C.M.; Godoi, I.P.D.; de Lima, M.E.; Ferreira, J.M.S. Animal venoms as a source of antiviral peptides active against arboviruses: A systematic review. Arch. Virol. 2022, 167, 1763–1772. [Google Scholar] [CrossRef] [PubMed]
- Julander, J.G.; Demarest, J.F.; Taylor, R.; Gowen, B.B.; Walling, D.M.; Mathis, A.; Babu, Y.S. An update on the progress of galidesivir (BCX4430), a broad-spectrum antiviral. Antiviral Res. 2021, 195, 105180. [Google Scholar] [CrossRef] [PubMed]
- Caporale, A.; Doti, N.; Monti, A.; Sandomenico, A.; Ruvo, M. Automatic procedures for the synthesis of difficult peptides using oxyma as activating reagent: A comparative study on the use of bases and on different deprotection and agitation conditions. Peptides 2018, 102, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Giugliano, R.; Ferraro, V.; Chianese, A.; Della Marca, R.; Zannella, C.; Galdiero, F.; Fasciana, T.M.A.; Giammanco, A.; Salerno, A.; Cannillo, J.; et al. Antiviral Properties of Moringa oleifera Leaf Extracts against Respiratory Viruses. Viruses 2024, 16, 1199. [Google Scholar] [CrossRef] [PubMed]
- Zannella, C.; Chianese, A.; Palomba, L.; Marcocci, M.E.; Bellavita, R.; Merlino, F.; Grieco, P.; Folliero, V.; De Filippis, A.; Mangoni, M.; et al. Broad-Spectrum Antiviral Activity of the Amphibian Antimicrobial Peptide Temporin L and Its Analogs. Int. J. Mol. Sci. 2022, 23, 2060. [Google Scholar] [CrossRef] [PubMed]
- Franci, G.; Falanga, A.; Zannella, C.; Folliero, V.; Martora, F.; Galdiero, M.; Galdiero, S.; Morelli, G.; Galdiero, M. Infectivity inhibition by overlapping synthetic peptides derived from the gH/gL heterodimer of herpes simplex virus type 1. J. Pept. Sci. 2017, 23, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Lei, C.; Yang, J.; Hu, J.; Sun, X. On the Calculation of TCID(50) for Quantitation of Virus Infectivity. Virol. Sin. 2021, 36, 141–144. [Google Scholar] [CrossRef] [PubMed]
- Ambrosino, A.; Chianese, A.; Zannella, C.; Piccolella, S.; Pacifico, S.; Giugliano, R.; Franci, G.; De Natale, A.; Pollio, A.; Pinto, G.; et al. Galdieria sulphuraria: An Extremophilic Alga as a Source of Antiviral Bioactive Compounds. Mar. Drugs 2023, 21, 383. [Google Scholar] [CrossRef] [PubMed]
- Brisbarre, N.M.; Plumet, S.; de Micco, P.; Leparc-Goffart, I.; Emonet, S.F. Toscana virus inhibits the interferon beta response in cell cultures. Virology 2013, 442, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Woelfl, F.; Leger, P.; Oreshkova, N.; Pahmeier, F.; Windhaber, S.; Koch, J.; Stanifer, M.; Roman Sosa, G.; Uckeley, Z.M.; Rey, F.A.; et al. Novel Toscana Virus Reverse Genetics System Establishes NSs as an Antagonist of Type I Interferon Responses. Viruses 2020, 12, 400. [Google Scholar] [CrossRef] [PubMed]
- Keskek Turk, Y.; Ergunay, K.; Kohl, A.; Hughes, J.; McKimmie, C.S. Toscana virus-an emerging Mediterranean arbovirus transmitted by sand flies. J. Gen. Virol. 2024, 105, 2045. [Google Scholar] [CrossRef] [PubMed]
- Di Nicuolo, G.; Pagliano, P.; Battisti, S.; Starace, M.; Mininni, V.; Attanasio, V.; Faella, F.S. Toscana virus central nervous system infections in southern Italy. J. Clin. Microbiol. 2005, 43, 6186–6188. [Google Scholar] [CrossRef] [PubMed]
- Cusi, M.G.; Gandolfo, C.; Terrosi, C.; Gori Savellini, G.; Belmonte, G.; Miracco, C. Toscana virus infects dendritic and endothelial cells opening the way for the central nervous system. J. Neurovirol. 2016, 22, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Braito, A.; Corbisiero, R.; Corradini, S.; Fiorentini, C.; Ciufolini, M.G. Toscana virus infections of the central nervous system in children: A report of 14 cases. J. Pediatr. 1998, 132, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Jaijakul, S.; Arias, C.A.; Hossain, M.; Arduino, R.C.; Wootton, S.H.; Hasbun, R. Toscana meningoencephalitis: A comparison to other viral central nervous system infections. J. Clin. Virol. 2012, 55, 204–208. [Google Scholar] [CrossRef] [PubMed]
- Naveca, F.G.; Almeida, T.A.P.; Souza, V.; Nascimento, V.; Silva, D.; Nascimento, F.; Mejia, M.; Oliveira, Y.S.; Rocha, L.; Xavier, N.; et al. Human outbreaks of a novel reassortant Oropouche virus in the Brazilian Amazon region. Nat. Med. 2024, 30, 3509–3521. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.T.; Tian, H.P.; Xu, J.L.; Liu, M.Q.; Cai, K.; Chen, S.L.; Ni, X.B.; Li, Y.R.; Hou, W.; Chen, L.J. Extensive genetic diversity of severe fever with thrombocytopenia syndrome virus circulating in Hubei Province, China, 2018–2022. PLoS Negl. Trop. Dis. 2023, 17, e0011654. [Google Scholar] [CrossRef] [PubMed]
- Alkan, C.; Bichaud, L.; de Lamballerie, X.; Alten, B.; Gould, E.A.; Charrel, R.N. Sandfly-borne phleboviruses of Eurasia and Africa: Epidemiology, genetic diversity, geographic range, control measures. Antivir. Res. 2013, 100, 54–74. [Google Scholar] [CrossRef] [PubMed]
- Ding, N.Z.; Luo, Z.F.; Niu, D.D.; Ji, W.; Kang, X.H.; Cai, S.S.; Xu, D.S.; Wang, Q.W.; He, C.Q. Identification of two severe fever with thrombocytopenia syndrome virus strains originating from reassortment. Virus Res. 2013, 178, 543–546. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.; Ayhan, N.; Cristovao, J.M.; Vilhena, H.; Martins, A.; Cachola, P.; Henriques, J.; Coimbra, M.; Catarino, A.; Lestinova, T.; et al. Antibody Response to Toscana Virus and Sandfly Fever Sicilian Virus in Cats Naturally Exposed to Phlebotomine Sand Fly Bites in Portugal. Microorganisms 2019, 7, 339. [Google Scholar] [CrossRef] [PubMed]
- Alwassouf, S.; Maia, C.; Ayhan, N.; Coimbra, M.; Cristovao, J.M.; Richet, H.; Bichaud, L.; Campino, L.; Charrel, R.N. Neutralization-based seroprevalence of Toscana virus and sandfly fever Sicilian virus in dogs and cats from Portugal. J. Gen. Virol. 2016, 97, 2816–2823. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Mari, J.M.; Palop-Borras, B.; Perez-Ruiz, M.; Sanbonmatsu-Gamez, S. Serosurvey study of Toscana virus in domestic animals, Granada, Spain. Vector Borne Zoonotic Dis. 2011, 11, 583–587. [Google Scholar] [CrossRef] [PubMed]
- Wensman, J.J.; Blomqvist, G.; Hjort, M.; Holst, B.S. Presence of antibodies to Schmallenberg virus in a dog in Sweden. J. Clin. Microbiol. 2013, 51, 2802–2803. [Google Scholar] [CrossRef] [PubMed]
- Giugliano, R.; Della Marca, R.; Chianese, A.; Monti, A.; Donadio, F.; Esposito, E.; Doti, N.; Zannella, C.; Galdiero, M.; De Filippis, A. The inhibitory potential of three scorpion venom peptides against multidrug-resistant Klebsiella pnemoniae. Front. Microbiol. 2025, 16, 1569719. [Google Scholar] [CrossRef] [PubMed]
- Xia, Z.; Wang, H.; Chen, W.; Wang, A.; Cao, Z. Scorpion Venom Antimicrobial Peptide Derivative BmKn2-T5 Inhibits Enterovirus 71 in the Early Stages of the Viral Life Cycle In Vitro. Biomolecules 2024, 14, 545. [Google Scholar] [CrossRef] [PubMed]
- Jlassi, A.; Mekni-Toujani, M.; Ferchichi, A.; Gharsallah, C.; Malosse, C.; Chamot-Rooke, J.; ElAyeb, M.; Ghram, A.; Srairi-Abid, N.; Daoud, S. BotCl, the First Chlorotoxin-like Peptide Inhibiting Newcastle Disease Virus: The Emergence of a New Scorpion Venom AMPs Family. Molecules 2023, 28, 4355. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pantinin-1 (µM) | Pantinin-2 (µM) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| SBV | TOSV | SBV | TOSV | |||||||
| CC50 | IC50 | SI | IC50 | SI | CC50 | IC50 | SI | IC50 | SI | |
| BHK-21 | 136.6 | 12.8 | 10.7 | 11 | 12.4 | 100.8 | 4.8 | 21 | 2.5 | 40.3 |
| A-72 | 93.6 | 6 | 15.6 | 3.8 | 24.6 | 90.7 | 4 | 22.7 | 3 | 30.2 |
| CRFK | 93.5 | 5.8 | 16.1 | 5 | 18.7 | 82.2 | 3.7 | 22.2 | 3 | 27.4 |
| U-87 | 114.3 | - | - | 12 | 9.5 | 82 | - | - | 3 | 27.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giugliano, R.; Zannella, C.; Della Marca, R.; Chianese, A.; Di Clemente, L.; Monti, A.; Doti, N.; Cacioppo, F.; Iovane, V.; Montagnaro, S.; et al. Expanding the Antiviral Spectrum of Scorpion-Derived Peptides Against Toscana Virus and Schmallenberg Virus. Pathogens 2025, 14, 713. https://doi.org/10.3390/pathogens14070713
Giugliano R, Zannella C, Della Marca R, Chianese A, Di Clemente L, Monti A, Doti N, Cacioppo F, Iovane V, Montagnaro S, et al. Expanding the Antiviral Spectrum of Scorpion-Derived Peptides Against Toscana Virus and Schmallenberg Virus. Pathogens. 2025; 14(7):713. https://doi.org/10.3390/pathogens14070713
Chicago/Turabian StyleGiugliano, Rosa, Carla Zannella, Roberta Della Marca, Annalisa Chianese, Laura Di Clemente, Alessandra Monti, Nunzianna Doti, Federica Cacioppo, Valentina Iovane, Serena Montagnaro, and et al. 2025. "Expanding the Antiviral Spectrum of Scorpion-Derived Peptides Against Toscana Virus and Schmallenberg Virus" Pathogens 14, no. 7: 713. https://doi.org/10.3390/pathogens14070713
APA StyleGiugliano, R., Zannella, C., Della Marca, R., Chianese, A., Di Clemente, L., Monti, A., Doti, N., Cacioppo, F., Iovane, V., Montagnaro, S., De Grazia, S., Galdiero, M., & De Filippis, A. (2025). Expanding the Antiviral Spectrum of Scorpion-Derived Peptides Against Toscana Virus and Schmallenberg Virus. Pathogens, 14(7), 713. https://doi.org/10.3390/pathogens14070713