Isolation and Molecular Characterization of Antimicrobial-Resistant Bacteria from Vegetable Foods

 ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Bacteriological Analyses
2.3. Molecular Analyses
2.4. Statistical Analyses
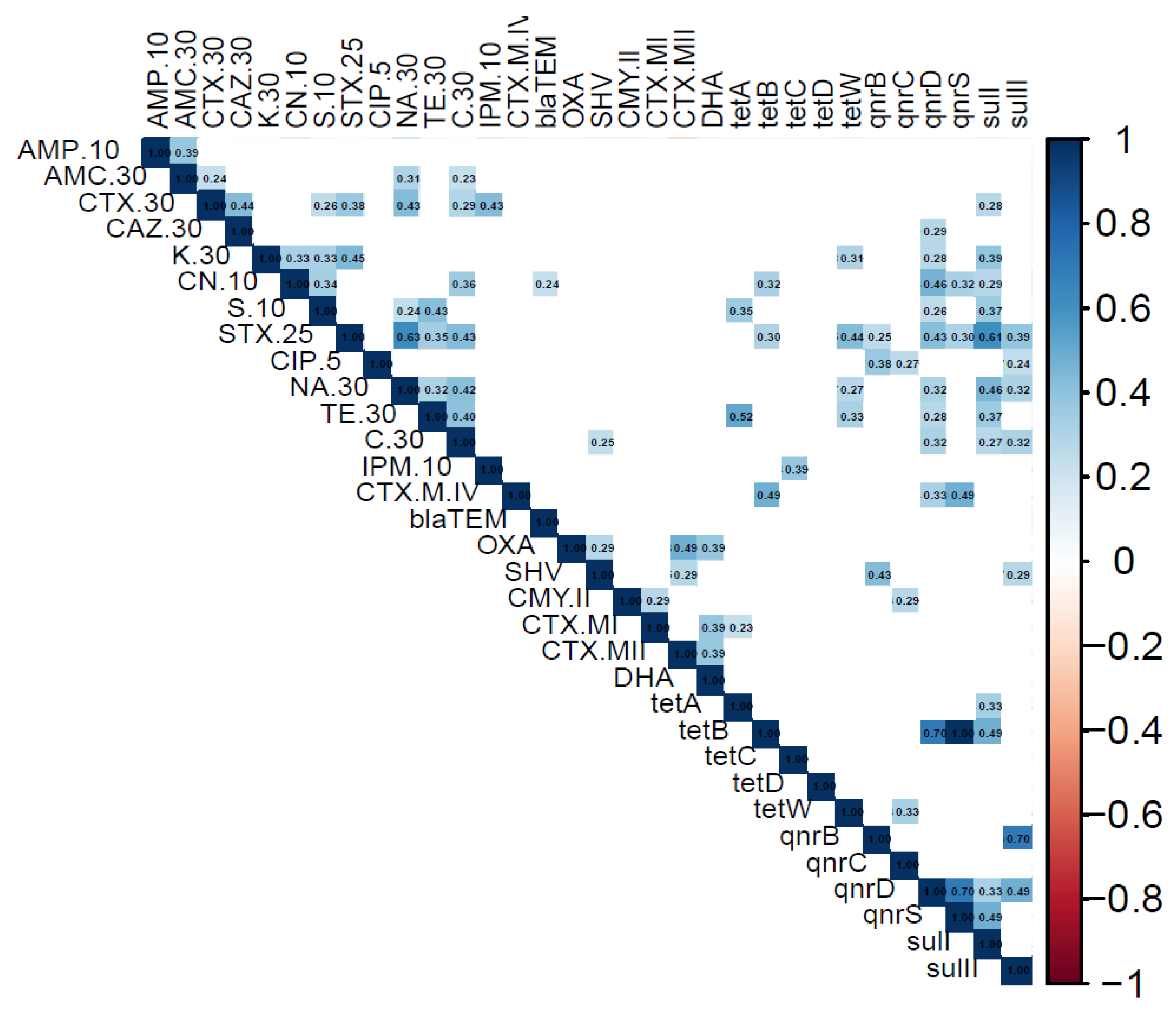
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fernández-Trapote, E.; Oliveira, M.; Cobo-Díaz, J.F.; Alvarez-Ordóñez, A. The Resistome of the Food Chain: A One Health Perspective. Microb. Biotechnol. 2024, 17, e14530. [Google Scholar] [CrossRef] [PubMed]
- Hutchings, M.I.; Truman, A.W.; Wilkinson, B. Antibiotics: Past, Present and Future. Curr. Opin. Microbiol. 2019, 51, 72–80. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control. Antimicrobial Consumption in the EU/EEA (ESAC-Net); Annual Epidemiological Report 2023; ECDC: Stockholm, Sweden, 2024. [Google Scholar]
- Council Recommendation on Stepping up EU Actions to Combat Antimicrobial Resistance in a One Health Approach 2023/C 220/01. Available online: https://eur-lex.europa.eu/legal-content/en/txt/?uri=celex:32023H0622(01) (accessed on 21 March 2025).
- Phan, D.; Bhattacharjee, A.S.; Hanan, D.; Park, S.; Herrera, D.; Ashworth, D.; Schmidt, M.; Men, Y.; Ferreira, J.F.S.; Ibekwe, A.M. Dissemination of Antimicrobial Resistance in Agricultural Ecosystems Following Irrigation with Treated Municipal Wastewater. Sci. Total Environ. 2024, 934, 173288. [Google Scholar] [CrossRef] [PubMed]
- Hijosa-Valsero, M.; Fink, G.; Schlüsener, M.P.; Sidrach-Cardona, R.; Martín-Villacorta, J.; Ternes, T.; Bécares, E. Removal of Antibiotics from Urban Wastewater by Constructed Wetland Optimization. Chemosphere 2011, 83, 713–719. [Google Scholar] [CrossRef]
- Rizzo, L.; Manaia, C.; Merlin, C.; Schwartz, T.; Dagot, C.; Ploy, M.C.; Michael, I.; Fatta-Kassinos, D. Urban Wastewater Treatment Plants as Hotspots for Antibiotic Resistant Bacteria and Genes Spread into the Environment: A Review. Sci. Total Environ. 2013, 447, 345–360. [Google Scholar] [CrossRef]
- Rowan, N.J. Defining Established and Emerging Microbial Risks in the Aquatic Environment: Current Knowledge, Implications, and Outlooks. Int. J. Microbiol. 2011, 2011, 462832. [Google Scholar] [CrossRef]
- Nguyen, A.Q.; Vu, H.P.; Nguyen, L.N.; Wang, Q.; Djordjevic, S.P.; Donner, E.; Yin, H.; Nghiem, L.D. Monitoring Antibiotic Resistance Genes in Wastewater Treatment: Current Strategies and Future Challenges. Sci. Total Environ. 2021, 783, 146964. [Google Scholar] [CrossRef]
- Taylor, P.; Reeder, R. Antibiotic Use on Crops in Low and Middle-Income Countries Based on Recommendations Made by Agricultural Advisors. CABI Agric. Biosci. 2020, 1, 1. [Google Scholar] [CrossRef]
- Mckenna, M. Antibiotics Set to Flood Florida’s Troubled Orange Orchards a Desperate Plan to Fight a Citrus Scourge Has Public-Health Advocates and Scientists Concerned. Nature 2019, 567, 302–303. [Google Scholar] [CrossRef]
- The Fruit and Vegetable Sector in the EU—A Statistical Overview—Statistics Explained—Eurostat. Available online: https://ec.europa.eu/eurostat/statistics-explained/index.php?title=The_fruit_and_vegetable_sector_in_the_EU_-_a_statistical_overview (accessed on 21 June 2025).
- European Union Sees Increase of Fruit and Vegetable Imports—Produce Business. Available online: https://producebusiness.com/european-union-sees-increase-of-fruit-and-vegetable-imports/#:~:text=Fresh%20fruit%20and%20vegetable%20imports%20into%20the%20EU,hub%2C%20handling%20about%20a%20third%20of%20total%20imports (accessed on 21 June 2025).
- European Commission. Agri-Food Trade Statistical Factsheet European Union—Extra EU27. 2024. Available online: https://www.eeas.europa.eu/sites/default/files/documents/2024/Agrifood%20Trade%20Statistical%20Factsheet-%20EU27.pdf (accessed on 21 June 2025).
- Kulik, K.; Lenart-Boroń, A.; Wyrzykowska, K. Impact of Antibiotic Pollution on the Bacterial Population within Surface Water with Special Focus on Mountain Rivers. Water 2023, 15, 975. [Google Scholar] [CrossRef]
- Tong, C.; Hu, H.; Chen, G.; Li, Z.; Li, A.; Zhang, J. Disinfectant Resistance in Bacteria: Mechanisms, Spread, and Resolution Strategies. Environ. Res. 2021, 195, 110897. [Google Scholar] [CrossRef]
- Ahmad, I.; Malak, H.A.; Abulreesh, H.H. Environmental Antimicrobial Resistance and Its Drivers: A Potential Threat to Public Health. J. Glob. Antimicrob. Resist. 2021, 27, 101–111. [Google Scholar]
- Regulation (EC) No 2073/2005 on Microbiological Criteria for Foodstuffs. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=celex%3A32005R2073 (accessed on 28 March 2025).
- Food and Agriculture Organization of the United Nations. Available online: https://Openknowledge.Fao.Org/Server/Api/Core/Bitstreams/9d071dca-6b05-4730-B9f2-2a53c48a3c6c/Content/Src/Html/Good-for-You.Html (accessed on 28 March 2025).
- World Health Organization. Available online: https://www.Who.Int/Tools/Elena/Interventions/Fruit-Vegetables-Ncds (accessed on 28 March 2025).
- Al-Kharousi, Z.S.; Guizani, N.; Al-Sadi, A.M.; Al-Bulushi, I.M. Antibiotic Resistance of Enterobacteriaceae Isolated from Fresh Fruits and Vegetables and Characterization of Their AmpC β-Lactamases. J. Food Prot. 2019, 82, 1857–1863. [Google Scholar] [CrossRef] [PubMed]
- Romyasamit, C.; Sornsenee, P.; Chimplee, S.; Yuwalaksanakun, S.; Wongprot, D.; Saengsuwan, P. Prevalence and Characterization of Extended-Spectrum β-Lactamase-Producing Escherichia coli and Klebsiella pneumoniae Isolated from Raw Vegetables Retailed in Southern Thailand. PeerJ 2021, 9, e11787. [Google Scholar] [CrossRef] [PubMed]
- ISO 6579-1:2017; Microbiology of the Food Chain—Horizontal Method for the Detection, Enumeration and Serotyping of Salmonella—Part 1: Detection of Salmonella spp. International Organization for Standardization: Geneva, Switzerland, 2017.
- ISO 11290-1:2017; Microbiology of the Food Chain—Horizontal Method for the Detection and Enumeration of Listeria monocytogenes and of Listeria spp.—Part 1: Detection Method. International Organization for Standardization: Geneva, Switzerland, 2017.
- ISO 16649-2:2010; Microbiology of Food and Animal Feeding Stuffs—Horizontal Method for the Enumeration of Beta-Glucuronidase-Positive Escherichia Coli—Part 2: Colony-Count Technique at 44 Degrees C Using 5-Bromo-4-Chloro-3-Indolyl Beta-D-Glucuronide. International Organization for Standardization: Geneva, Switzerland, 2010.
- ISO 21528-2:2017; Microbiology of the Food Chain—Horizontal Method for the Detection and Enumeration of Enterobacteriaceae—Part 2: Colony-Count Technique. International Organization for Standardization: Geneva, Switzerland, 2017.
- CLSI. Performance Standards for Antimicrobial Susceptibility: Supplement M100, 31st ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2021; ISBN 978-1-68440-105-5. [Google Scholar]
- Woodman, M.E.; Savage, C.R.; Arnold, W.K.; Stevenson, B. Direct PCR of Intact Bacteria (Colony PCR). CP Microbiol. 2016, 42, A-3D. [Google Scholar] [CrossRef]
- Coy, M.R.; Hoffmann, M.; Kingdom Gibbard, H.N.; Kuhns, E.H.; Pelz-Stelinski, K.S.; Stelinski, L.L. Nested-Quantitative PCR Approach with Improved Sensitivity for the Detection of Low Titer Levels of Candidatus Liberibacter Asiaticus in the Asian Citrus Psyllid, Diaphorina Citri Kuwayama. J. Microbiol. Methods 2014, 102, 15–22. [Google Scholar] [CrossRef]
- Kim, J.; Jeon, S.; Rhie, H.; Lee, B.; Park, M.; Lee, H.; Lee, J.; Kim, S. Rapid Detection of Extended Spectrum β-Lactamase (ESBL) for Enterobacteriaceae by Use of a Multiplex PCR-Based Method. Infect. Chemother. 2009, 41, 181. [Google Scholar] [CrossRef]
- Gargano, V.; Sciortino, S.; Gambino, D.; Costa, A.; Agozzino, V.; Reale, S.; Alduina, R.; Vicari, D. Antibiotic Susceptibility Profile and Tetracycline Resistance Genes Detection in Salmonella spp. Strains Isolated from Animals and Food. Antibiotics 2021, 10, 809. [Google Scholar] [CrossRef]
- Zhao, X.; Hu, M.; Zhang, Q.; Zhao, C.; Zhang, Y.; Li, L.; Qi, J.; Luo, Y.; Zhou, D.; Liu, Y. Characterization of Integrons and Antimicrobial Resistance in Salmonella from Broilers in Shandong, China. Poult. Sci. 2020, 99, 7046–7054. [Google Scholar] [CrossRef]
- Sucato, A.; Vecchioni, L.; Savoca, D.; Presentato, A.; Arculeo, M.; Alduina, R. A Comparative Analysis of Aquatic and Polyethylene-Associated Antibiotic-Resistant Microbiota in the Mediterranean Sea. Biology 2021, 10, 200. [Google Scholar] [CrossRef]
- Lynne, A.M.; Rhodes-Clark, B.S.; Bliven, K.; Zhao, S.; Foley, S.L. Antimicrobial Resistance Genes Associated with Salmonella enterica Serovar Newport Isolates from Food Animals. Antimicrob. Agents Chemother. 2008, 52, 353–356. [Google Scholar] [CrossRef] [PubMed]
- Hassuna, N.A.; Darwish, M.K.; Sayed, M.; Ibrahem, R.A. Molecular Epidemiology and Mechanisms of High-Level Resistance to Meropenem and Imipenem in Pseudomonas aeruginosa. IDR 2020, 13, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Randall, L.P. Antibiotic Resistance Genes, Integrons and Multiple Antibiotic Resistance in Thirty-Five Serotypes of Salmonella enterica Isolated from Humans and Animals in the UK. J. Antimicrob. Chemother. 2004, 53, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Kizheva, Y.; Georgiev, G.; Donchev, D.; Dimitrova, M.; Pandova, M.; Rasheva, I.; Hristova, P. Cross-Over Pathogenic Bacteria Detected in Infected Tomatoes (Solanum lycopersicum L.) and Peppers (Capsicum annuum L.) in Bulgaria. Pathogens 2022, 11, 1507. [Google Scholar] [CrossRef]
- European Food Safety Authority. Prolonged Multi-Country Outbreak of Salmonella Strathcona ST2559 Linked to Consumption of Tomatoes in the EU/EEA and the UK. EFS3 2024, 21, 9107E. [Google Scholar] [CrossRef]
- Freeland, G.; Hettiarachchy, N.; Atungulu, G.G.; Apple, J.; Mukherjee, S. Strategies to Combat Antimicrobial Resistance from Farm to Table. Food Rev. Int. 2023, 39, 27–40. [Google Scholar] [CrossRef]
- Poeys-Carvalho, R.M.P.; Gonzalez, A.G.M. Resistance to β-Lactams in Enterobacteriaceae Isolated from Vegetables: A Review. Crit. Rev. Food Sci. Nutr. 2025, 65, 936–946. [Google Scholar] [CrossRef]
- Mezzatesta, M.L.; Gona, F.; Stefani, S. Enterobacter cloacae Complex: Clinical Impact and Emerging Antibiotic Resistance. Future Microbiol. 2012, 7, 887–902. [Google Scholar] [CrossRef]
- Gomez-Simmonds, A.; Annavajhala, M.K.; Wang, Z.; Macesic, N.; Hu, Y.; Giddins, M.J.; O’Malley, A.; Toussaint, N.C.; Whittier, S.; Torres, V.J.; et al. Genomic and Geographic Context for the Evolution of High-Risk Carbapenem-Resistant Enterobacter cloacae Complex Clones ST171 and ST78. mBio 2018, 9, e00542-18. [Google Scholar] [CrossRef]
- Liu, L.-H.; Wang, N.-Y.; Wu, A.Y.-J.; Lin, C.-C.; Lee, C.-M.; Liu, C.-P. Citrobacter Freundii Bacteremia: Risk Factors of Mortality and Prevalence of Resistance Genes. J. Microbiol. Immunol. Infect. 2018, 51, 565–572. [Google Scholar] [CrossRef]
- Crone, S.; Vives-Flórez, M.; Kvich, L.; Saunders, A.M.; Malone, M.; Nicolaisen, M.H.; Martínez-García, E.; Rojas-Acosta, C.; Catalina Gomez-Puerto, M.; Calum, H.; et al. The Environmental Occurrence of Pseudomonas aeruginosa. APMIS 2020, 128, 220–231. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, R.; Rekha, N.D.; Gopal, S. Pseudomonas aeruginosa Biofilm: Treatment Strategies to Combat Infection. Arch. Microbiol. 2025, 207, 141. [Google Scholar] [CrossRef] [PubMed]
- Wareth, G.; Neubauer, H. The Animal-Foods-Environment Interface of Klebsiella Pneumoniae in Germany: An Observational Study on Pathogenicity, Resistance Development and the Current Situation. Vet. Res. 2021, 52, 16. [Google Scholar] [CrossRef]
- Zhang, C.-M.; Yuan, Q.-Q.; Li, Y.-Q.; Liu, A. Characteristics of Heterotrophic Endophytic Bacteria in Four Kinds of Edible Raw Vegetables: Species Distribution, Antibiotic Resistance, and Related Genes. Lett. Appl. Microbiol. 2024, 77, ovae120. [Google Scholar] [CrossRef] [PubMed]
- Oteo, J.; Delgado-Iribarren, A.; Vega, D.; Bautista, V.; Rodríguez, M.C.; Velasco, M.; Saavedra, J.M.; Pérez-Vázquez, M.; García-Cobos, S.; Martínez-Martínez, L.; et al. Emergence of Imipenem Resistance in Clinical Escherichia coli during Therapy. Int. J. Antimicrob. Agents 2008, 32, 534–537. [Google Scholar] [CrossRef]
- Agrawal, G.N. A Study on the AmpC Production Amongst the Urinary Enterobacteriaceae Isolates. J. Clin. Diagn. Res. 2013, 7, 1831–1832. [Google Scholar] [CrossRef]
- Van Hoek, A.H.A.M.; Veenman, C.; Van Overbeek, W.M.; Lynch, G.; De Roda Husman, A.M.; Blaak, H. Prevalence and Characterization of ESBL- and AmpC-Producing Enterobacteriaceae on Retail Vegetables. Int. J. Food Microbiol. 2015, 204, 1–8. [Google Scholar] [CrossRef]
- Qi, Z.; Le, Z.; Han, F.; Qi, Y.; Liu, R. β-Lactamase Genes Transmission Influenced by Tetracycline, Sulfonamide and β-Lactams Antibiotics Contamination in the on-Site Farm Soil. Ecotoxicol. Environ. Saf. 2022, 241, 113753. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, N.; Li, C.; Shao, L. Diversity of Antibiotic Resistance Genes in Soils with Four Different Fertilization Treatments. Front. Microbiol. 2023, 14, 1291599. [Google Scholar] [CrossRef]
- Pina-Sánchez, M.; Rua, M.; Del Pozo, J.L. Present and Future of Resistance in Pseudomonas aeruginosa: Implications for Treatment. Rev. Esp. Quim. 2023, 36, 54–58. [Google Scholar] [CrossRef]
- Garcia-Bustos, V.; Cabañero-Navalón, M.D.; Salavert Lletí, M. Resistance to Beta-Lactams in Gram-Negative Bacilli: Relevance and Potential Therapeutic Alternatives. Rev. Esp. Quim. 2022, 35, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Imperial, I.C.; Pabustan, P.M.; Valencia, K.A.; Nicdao, M.A.; Ibana, J. Emergence of Resistance Genes in Fecal Samples of Antibiotic-Treated Philippine Broilers Emphasizes the Need to Review Local Farming Practices. Trop. Biomed. 2022, 39, 150–159. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Dai, J.; Cao, C.; Su, L.; Cao, M.; He, Y.; Li, M.; Zhang, Z.; Chen, J.; Cui, S.; et al. Prevalence, Serotype, Antimicrobial Susceptibility, Contamination Factors, and Control Methods of Salmonella spp. in Retail Fresh Fruits and Vegetables: A Systematic Review and Meta-analysis. Comp. Rev. Food Sci. Food Saf. 2024, 23, e13407. [Google Scholar] [CrossRef]
- Jiménez-Belenguer, A.I.; Ferrús, M.A.; Hernández, M.; García-Hernández, J.; Moreno, Y.; Castillo, M.Á. Prevalence and Characterization of Beta-Lactam and Carbapenem-Resistant Bacteria Isolated from Organic Fresh Produce Retailed in Eastern Spain. Antibiotics 2023, 12, 387. [Google Scholar] [CrossRef] [PubMed]
- Colosi, I.A.; Baciu, A.M.; Opriș, R.V.; Peca, L.; Gudat, T.; Simon, L.M.; Colosi, H.A.; Costache, C. Prevalence of ESBL, AmpC and Carbapenemase-Producing Enterobacterales Isolated from Raw Vegetables Retailed in Romania. Foods 2020, 9, 1726. [Google Scholar] [CrossRef]
- Zhang, P.; Ji, L.; Wu, X.; Chen, L.; Yan, W.; Dong, F. Prevalence, Genotypic Characteristics, and Antibiotic Resistance of Listeria Monocytogenes from Retail Foods in Huzhou, China. J. Food Prot. 2024, 87, 100307. [Google Scholar] [CrossRef]
- Siri, Y.; Precha, N.; Sirikanchana, K.; Haramoto, E.; Makkaew, P. Antimicrobial Resistance in Southeast Asian Water Environments: A Systematic Review of Current Evidence and Future Research Directions. Sci. Total Environ. 2023, 896, 165229. [Google Scholar] [CrossRef]
- Federigi, I.; Bonetta, S.; Tesauro, M.; De Giglio, O.; Oliveri Conti, G.; Atomsa, N.T.; Bagordo, F.; Bonetta, S.; Consonni, M.; Diella, G.; et al. A Systematic Scoping Review of Antibiotic-Resistance in Drinking Tap Water. Environ. Res. 2024, 263, 120075. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, Q.; Chen, S.; Zhang, Z.; Song, J.; Long, Z.; Yu, Y.; Fang, H. Enterobacteriaceae Predominate in the Endophytic Microbiome and Contribute to the Resistome of Strawberry. Sci. Total Environ. 2020, 727, 138708. [Google Scholar] [CrossRef]

{kind=link}
| Target Gene | Primer Sequence (5′→3′) | Tm (°C) | Amplicon Size (bp) | Reference |
|---|---|---|---|---|
| 16S rDNA | F1: GAGTTTGATCCTGGCTCAG R12: ACGGCTACCTTGTTACGACT | 56 | 1402 | [29] |
| CTX-M IV | F: GACAAAGAGAGTGCAACGGATG R: TCAGTGCGATCCAGACGAAA | 61 | 501 | [30] |
| TEM | F: AGTGCTGCCATAACCATGAGTG R: CTGACTCCCC GTCGTGTAGATA | 61 | 431 | |
| OXA | F: ATTATCTACAGCAGCGCCAGTG R: TGCATCCACGTCTTTGGTG | 61 | 296 | |
| SHV | F: GATGAACGCTTTCCCATGATG R: CGCTGTTATCGCTCATGGTAA | 61 | 214 | |
| CMY II | F: AGCGATCCGGTCACGAAATA R: CCCGTTTTATG CACCCATGA | 61 | 695 | |
| CTX M I | F: TCCAGAATAAGGAATCCCATGG R: TGCTTTACCCAGCGTCAGAT | 61 | 621 | |
| CTX M II | F: ACCGCCGATAATTCGCAGAT R: GATATCGTTGGTGGTGCCATAA | 61 | 588 | |
| DHA | F: GTGGTGGACAGCACCATTAAA R: CCTGCGGTATAGGTAGCCAGAT | 61 | 314 | |
| tetA | F: GCTACATCCTGCTTGCCTTC R: CATAGATCGCCGTGAAGAGG | 60 | 210 | [31] |
| tetB | F: TTGGTTAGGGGCAAGTTTTG R: GTAATGGGCCAATAACACCG | 60 | 659 | |
| tetC | CTTGAGAGCCTTCAACCCAG ATGGTCGTCATCTACTGCC | 60 | 418 | |
| tetD | AAACCATTACGGCATTCTGC GACCGGATACACCATCCATC | 60 | 787 | |
| tetE | F: AAACCACATCCTCCATACGC R: AAATAGGCCACAACCGTCAG | 60 | 278 | |
| tetG | F: GCTCGGTGGTATCTCTGCTC R: AGCAACAGAATCGGGAACAC | 60 | 844 | |
| tetO | F: GGAGGGGTTCAACCACAAAG R: CTATGTAAATAAAATGGATAG | 55 | 88 | |
| qnrA | F: ATTTCTCACGCCAGGATTTG R: TGCCAGGCACAGATCTTGAC | 60 | 516 | [32] |
| qnrB | F: CGACCTKAGCGGCACTGAAT R: GAGCAACGAYGCCTGGTAGYTG | 50 | 515 | |
| qnrC | F: GGGTTGTACATTTATTGAATC R: TCCACTTTACGAGGTTCT | 50 | 446 | |
| qnrD | F: CGAGATCAATTTACGGGGAATA R: AACAAGCTGAAGCGCCTG | 50 | 581 | |
| qnrS | F: GACGTGCTAACTTGCGTGAT R: TGGCATTGTTGGAAACTTG | 62 | 118 | [33] |
| sul-I | F: TCACCGAGGACTCCTTCTTC R: AATATCGGGATAGAGCGCAG | 60 | 316 | [34] |
| sul-II | F: TCCGGTGGAGGCCGGTATCTGG R: CGGGAATGCCATCTGCCTTGAG | 60 | 191 | [31] |
| sul-III | F: GAGCAAGATTTTTGGAATCG R: TCTGCAGCTAACCTAGGGCTTGGA | 51 | 880 | [34] |
| vim1 | F: AGTGGTGAGTATCCGACAG R: ATGAAAGTGCGTGGAGAC | 60 | 261 | [35] |
| vim2 | F: ATGTTCAAACTTTTGAGTAAG R: CTACTCAACGACTGAGCG | 60 | 801 | |
| ndm | F: GGTTTGGCGATCTGGTTTTC R: CGGAATGGCTCATCACGATC | 60 | 621 | |
| imp1 | F: CTACCGCAGCAGAGTCTTTG R: AACCAGTTTTGCCTTACCAT. | 55 | 587 | |
| imp2 | F: GTTTTATGTGTATGCTTCC R: AGCCTGTTCCCATGTAC | 55 | 678 | |
| cat-1 | F: CCTATAACCAGACCGTTCAG R: TCACAGACGGCATGATGAAC | 56 | 495 | [36] |
| cat-2 | F: CCGGATTGACCTGAATACCT R: TCACATACTGCATGATGAAC | 56 | 572 | |
| floR | F: AACCCGCCCTCTGGATCAAGTCAA R: CAAATCACGGGCCACGCTGTATC | 60 | 548 | |
| dhfr 1 | F: GTGAAACTATCACTAATGGTAGCT R: ACCCTTTTGCCAGATTTGGTAACT | 54 | 470 |
| Enterococci | Enterobacteriaceae | ||||
|---|---|---|---|---|---|
| No. of Samples | M ± SD 2 | n. of Samples | M ± SD 2 | ||
| Fresh vegetables | 11 | 3.44 ± 1.28 | Fresh vegetables | 23 | 3.09 ± 1.09 |
| RTE 1 vegetable | 1 | 1 | RTE 1 vegetable | 14 | 3.29 ± 1.22 |
| Total | 12 | 3.24 ± 1.41 | 37 | 3.17 ± 1.21 | |
| Strain ID | Species | AMR Profile | |
|---|---|---|---|
| R | I | ||
| 1 | E. 1 cloacae | AMP10, AMC30 | |
| 2 | E. 2 coli | AMP10, AMC30, TE30 | |
| 3 | H. 3 alvei | AMP10, AMC30 | CAZ30 |
| 4 | H. 3 alvei | AMP10, AMC30 | CAZ30 |
| 5 | E. 1 cloacae | AMP10, AMC30 | |
| 6 | E. 1 cloacae | AMP10, AMC30, STX25, TE30, C30 | K30, NA30 |
| 7 | P. 4 fluorescens | AMP10, AMC30, CTX30, NA30, TE30, C30, IPM10 | |
| 8 | Acinetobacter | AMP10, AMC30 | |
| 10 | P. 4 aeruginosa | AMP10, AMC30, K30, CN10, S10, STX25, NA30, TE30, C30 | |
| 11 | C. 5 freundii | AMP10, AMC30 | |
| 12 | E. 1 cloacae | AMP10, AMC30 | |
| 13 | C. 5 freundii | AMP10, AMC30 | |
| 14 | M. 6 morganii | AMP10, AMC30, TE30, C30 | |
| 15 | P. 4 aeruginosa | AMP10, AMC30, K30, CN10, S10, STX25, NA30, TE30, C30 | CTX30, CAZ30 |
| 16 | E. 1 cloacae | AMP10 | AMC30 |
| 17 | C. 5 freundii | AMP10, AMC30 | |
| 18 | P. 7 rettgeri | AMC30, TE30 | |
| 19 | C. 5 freundii | AMP10, AMC30 | |
| 20 | P. 4 aeruginosa | AMP10, AMC30, K30, CN10, S10, STX25, NA30, TE30, C30 | CTX30, CAZ30 |
| 21 | E. 1 cloacae | AMP10, AMC30 | |
| 22 | E. 1 cloacae | AMP10, AMC30 | |
| 23 | K. 8 pneumoniae | AMP10, K30, CN10, S10, TE30 | |
| 24 | E. 1 cloacae | AMP10, AMC30, CN10 | K30, S10, TE30, C30 |
| 26 | K. 8 oxytoca | AMP10, S10 | K30, CN10 |
| 27 | Citrobacter | K30 | AMP10 |
| 28 | K. 8 pneumoniae | AMP10, AMC30 | S10 |
| 29 | Enterobacter (EMP) | AMP10, K30 | S10 |
| 30 | K. 8 pneumoniae | AMP10 | S10 |
| 31 | E. 1 cloacae | AMP10, AMC30, K30, S10 | |
| 35 | P. 7 stuartii | AMP10, AMC30, S30, TE30 | K30 |
| 36 | E. 1 cloacae | AMP10, AMC30, S30, TE30 | |
| 38 | E. 1 cloacae | AMP10, AMC30, NA30, C30 | STX25, CIP5 |
| 49 | P. 4 fluorescens | AMP10, AMC30, CTX30, STX25, NA30 | |
| 50 | E. 1 cloacae | AMP10, AMC30, CTX30, CN10 | C30 |
| 51 | K. 8 pneumoniae spp. ozaenae | CTX30 | AMP10, AMC30, CN10, C30 |
| 52 | E. 1 cloacae | AMP10, AMC30, CN10 | |
| 53 | E. 1 cloacae | AMP10, AMC30, CTX30, NA30 | S10, IPM10 |
| 54 | E. 1 cloacae | AMP10, AMC30, CN10; I: CTX30, K30, IPM10 | |
| 56 | P. 4 aeuruginosa | AMP10, AMC30, CTX30, K30, CN10, STX25, NA30 | TE30 |
| 57 | R. 9 ornithlytica | AMP10, AMC30, CTX30, CAZ30, S10, CIP 5, C30 | NA30 |
| 58 | Cronobacter spp. | R: AMP10, AMC30, CTX30, S10, C30, IPM10 | CAZ30, K30, CN10, NA30 |
| 59 | Pantoea spp. | K30, STX25, CIP 5, C30 | AMP10, AMC30, CTX30, CAZ30 |
| 60 | K. 8 oxytoca | AMP10, AMC30, K30 | S10 |
| 61 | A. 10 hydrophila | AMP10, AMC30, K30 | CIP 5 |
| 62 | C. 5 freundii | AMP10, AMC30, S10 | CIP 5 |
| 65 | R. 11 pickettii | AMP10, AMC30 | NA30, C30 |
| 67 | E. 1 cloacae | AMP10, AMC30; I: K30, S10, IPM10 | |
| 68 | P. 7 rettgerii | AMC30, TE30 | CAZ30, S10 |
| 69 | E. 1 cloacae | AMP10, AMC30 | K30, TE30 |
| 70 | E. 3 coli type 1 | AMP10, AMC30, S10 | NA30, TE30 |
| 75 | K. 8 pneumoniae spp. pneumoniae | AMP10, AMC30, C30 | CAZ30 |
| Strain ID | Species | AMR Profile | |
|---|---|---|---|
| R | I | ||
| 37 | E. 1 cloacae | AMP10, AMC30 | |
| 39 | R. 2 ornithinolytica | AMP10 | |
| 40 | C. 3 freundii | AMP10 | AMC30 |
| 41 | Pantoea spp. | AMP10 | |
| 42 | P. 4 fluorescens | AMP10, AMC30, NA30 | CAZ30 |
| 43 | K. 5 pneumoniae spp. ozaenae | AMP10, K30 | |
| 44 | K. 5 oxytoca | AMP10 | |
| 45 | C. 3 youngae | AMP10, K30 | |
| 46 | E. 6 coli | AMP10 | |
| 47 | R. 2 aquatilis | AMP10, K30 | |
| 48 | K. 5 pneumoniae spp. pneumoniae | AMP10, AMC30, K30 | S10 |
| 55 | E. 1 cloacae | AMP10, AMC30, TE30 | CTX30, CAZ30, S10, C30, IPM10 |
| 63 | E. 6 coli tipo 1 | AMP10, S10 | AMC30, CTX30, CAZ30 |
| 64 | M. 7 morganii | AMP10, AMC30 | CTX30, K30, S10, IPM10 |
| 66 | K. 5 pneumoniae | S10 | AMP10, AMC30 |
| 71 | P. 4 aeuruginosa | AMP10, AMC30, CTX30, K30, S10, STX25, NA30, TE30 | C30 |
| 72 | E. 1 cloacae | AMP10, AMC30, S10, CIP5 | CTX30 |
| 73 | C. 3 youngae | AMP10, S10, C30 | AMC30, TE30 |
| 74 | C. 3 youngae | C30 | AMC30, TE30 |
| 76 | C. 3 freundii | AMP10, AMC30, CAZ30, NA30, TE30 | S10 |
| 77 | E. 1 cloacae | AMP10, AMC30 | |
| 78 | A. 8 hydrophila | AMP10 | AMC30, CAZ30, K30 |
| 79 | E. 1 cloacae | AMP10, AMC30 | |
| 80 | H. 9 alvei | AMP10, AMC30, CTX30, CAZ30 | |
| 81 | E. 1 cloacae | AMP10, AMC30, CAZ30 | CTX30, IPM10 |
| 82 | P. 4 aeruginosa | AMP10, AMC30, K30, STX25, NA30 | CTX30, S10, TE30 |
| 83 | K. 5 oxytoca | AMP10 | AMC30, NA30 |
| AMP | AMC | CTX | CAZ | K | CN | S | STX | CIP | NA | TE | C | IPM | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| R (%) | 88.5 | 69.2 | 12.8 | 6.4 | 20.5 | 11.5 | 20.5 | 11.5 | 3.8 | 15.4 | 19.2 | 16.7 | 2.6 |
| I (%) | 7.7 | 15.4 | 12.8 | 15.4 | 14.1 | 3.9 | 18.0 | 1.3 | 3.9 | 7.7 | 9.0 | 7.7 | 7.7 |
| S (%) | 3.8 | 15.4 | 74.4 | 78.2 | 65.4 | 84.6 | 61.5 | 87.2 | 92.3 | 76.9 | 71.8 | 75.6 | 89.7 |
| GENE | N | % | |
|---|---|---|---|
| β-lactams | TEM | 14/76 | 18.4% |
| CTX-M IV | 5/76 | 6.6% | |
| SHV | 3/76 | 3.9% | |
| DHA | 3/76 | 3.9% | |
| CTX-MI | 3/76 | 3.9% | |
| CMY-II | 2/76 | 2.6% | |
| OXA | 2/76 | 2.6% | |
| CTX-MII | 0/76 | 0% | |
| Tetracyclines | tetA | 8/22 | 36.4% |
| tetW | 4/22 | 18.2% | |
| tetB | 1/22 | 4.5% | |
| tetC | 1/22 | 4.5% | |
| tetD | 1/22 | 4.5% | |
| tetE | 1/22 | 4.5% | |
| tetG | 0/22 | 0% | |
| tetO | 0/22 | 0% | |
| Sulphonamides | sul-I | 4/11 | 36% |
| sul-II | 0/11 | 0% | |
| sul-III | 2/11 | 18% | |
| Quinolones | qnrD | 7/22 | 31.8% |
| qnrB | 3/22 | 13.6% | |
| qnrC | 2/22 | 9.1% | |
| qnrS | 1/22 | 4.5% | |
| qnrA | 0/22 | 0% | |
| Carbapenems | vim1 | 0/7 | 0% |
| vim2 | 0/7 | 0% | |
| ndm | 0/7 | 0% | |
| imp1 | 0/17 | 0% | |
| imp2 | 0/17 | 0% | |
| Chloramphenicol | cat1 | 0/17 | 0% |
| cat2 | 0/17 | 0% | |
| floR | 0/17 | 0% | |
| dfloR | 0/17 | 0% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castello, A.; Massaro, C.; Seghers, E.; Ferraro, C.; Costa, A.; Alduina, R.; Cardamone, C. Isolation and Molecular Characterization of Antimicrobial-Resistant Bacteria from Vegetable Foods. Pathogens 2025, 14, 682. https://doi.org/10.3390/pathogens14070682
Castello A, Massaro C, Seghers E, Ferraro C, Costa A, Alduina R, Cardamone C. Isolation and Molecular Characterization of Antimicrobial-Resistant Bacteria from Vegetable Foods. Pathogens. 2025; 14(7):682. https://doi.org/10.3390/pathogens14070682
Chicago/Turabian StyleCastello, Annamaria, Chiara Massaro, Erine Seghers, Clelia Ferraro, Antonella Costa, Rosa Alduina, and Cinzia Cardamone. 2025. "Isolation and Molecular Characterization of Antimicrobial-Resistant Bacteria from Vegetable Foods" Pathogens 14, no. 7: 682. https://doi.org/10.3390/pathogens14070682
APA StyleCastello, A., Massaro, C., Seghers, E., Ferraro, C., Costa, A., Alduina, R., & Cardamone, C. (2025). Isolation and Molecular Characterization of Antimicrobial-Resistant Bacteria from Vegetable Foods. Pathogens, 14(7), 682. https://doi.org/10.3390/pathogens14070682