
Gut–Vaginal Microbiome Crosstalk in Ovarian Cancer: Implications for Early Diagnosis

 , ,
, ,
Abstract
1. Introduction
2. Characteristics of the Gut and Vaginal Microbiota in Ovarian Cancer
2.1. Characteristics of the Gut and Vaginal Microbiota in Healthy Women
2.2. Characteristics of the Gut and Vaginal Microbiota in Patients with Ovarian Cancer
3. Potential Mechanisms of Action of the Gut and Vaginal Microbiomes in Ovarian Cancer
3.1. Crosstalk Between Gut and Vaginal Microbiomes
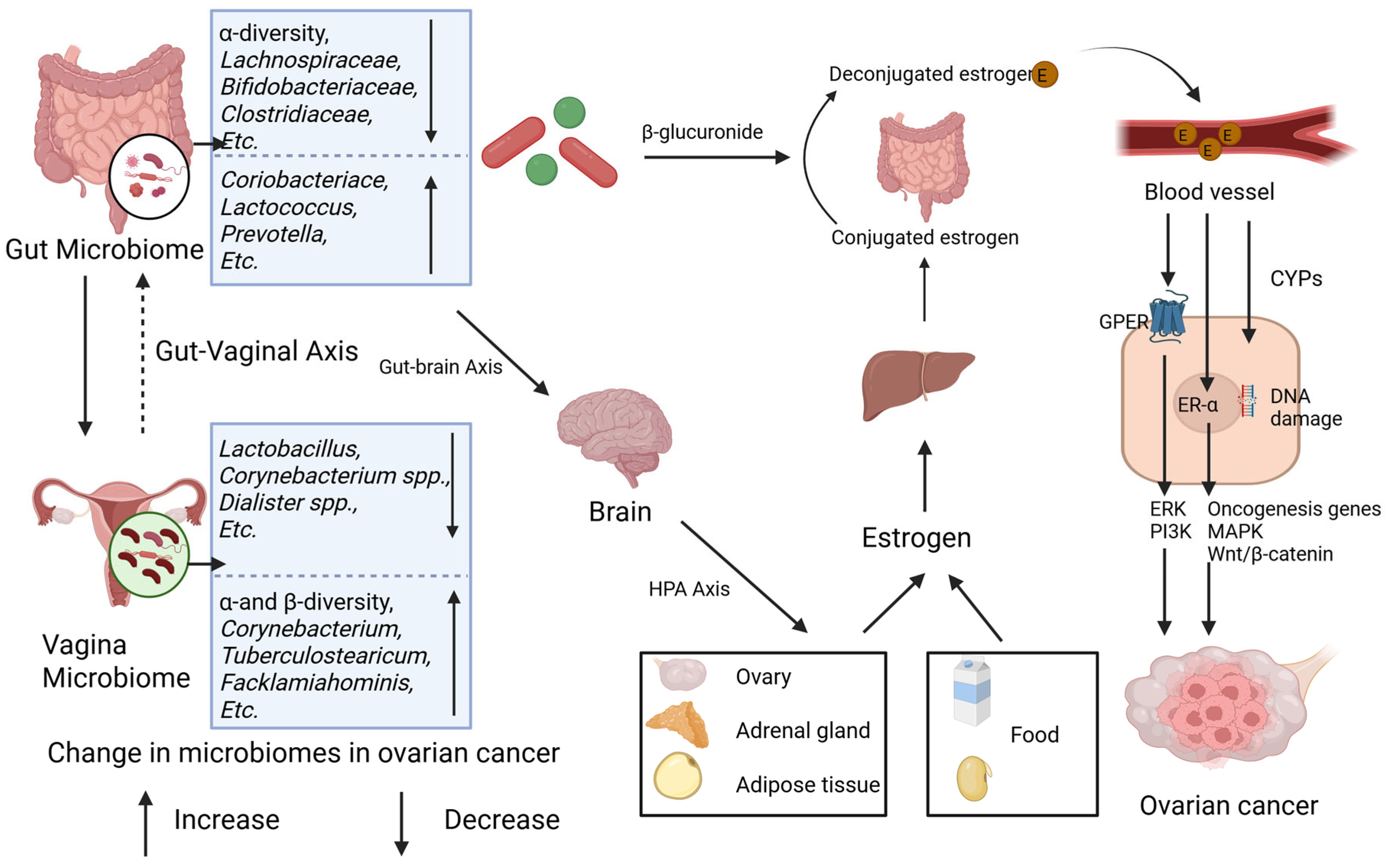
3.2. Estrogen Metabolism Potentiation of the Gut and Vaginal Microbiomes in Ovarian Cancer
3.2.1. Estrogen Metabolism
3.2.2. Epidemiological Evidence and Mechanisms of the Association Between Estrogen and Ovarian Cancer
3.2.3. Microbiome–Estrogen-Mediated Ovarian Cancer
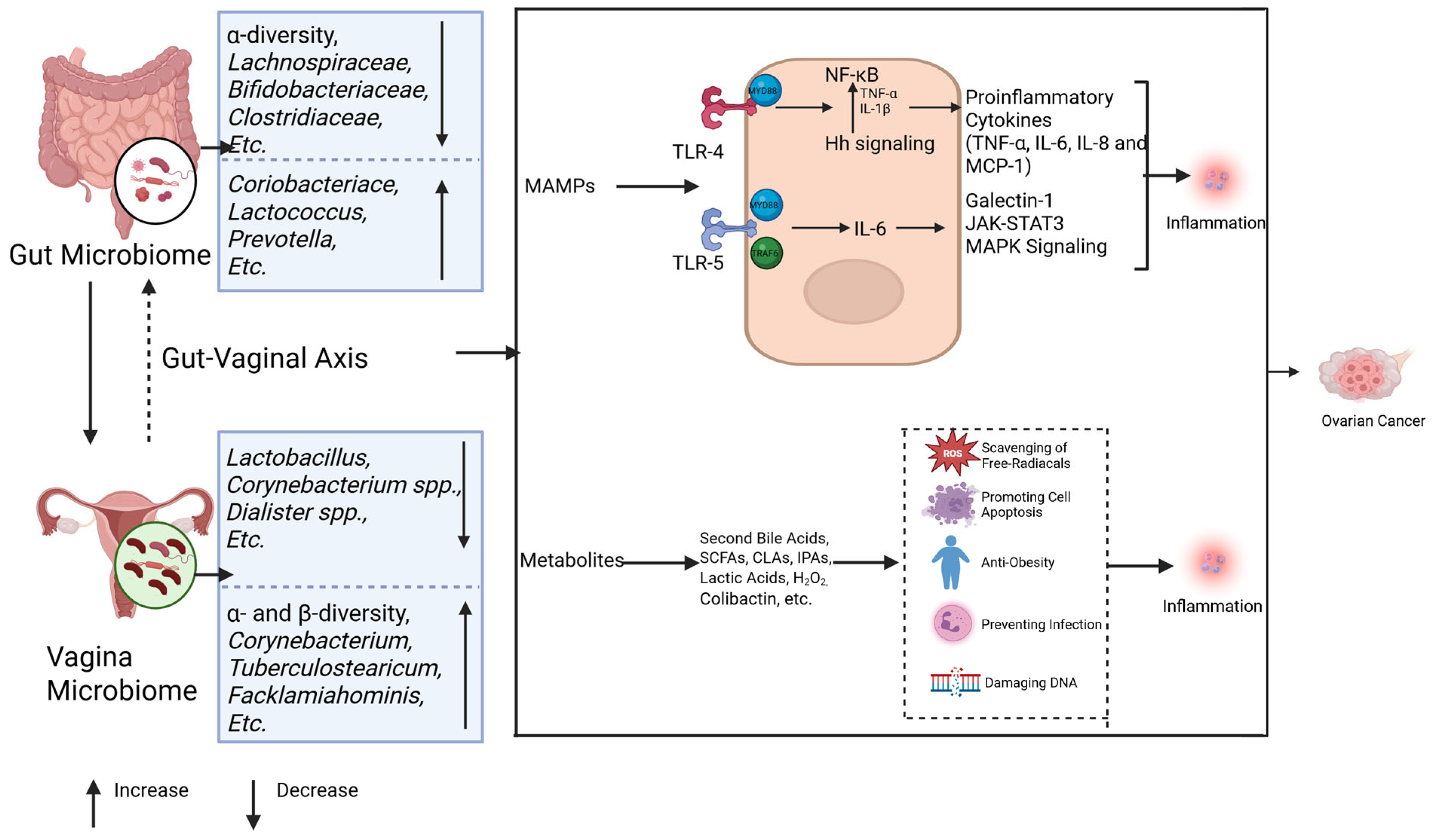
3.3. Chronic Inflammatory Priming by the Gut and Vaginal Microbiomes in Ovarian Cancer
3.3.1. The Gut Microbiota Influences Ovarian Cancer Development Through Inflammation
3.3.2. Vaginal Microbes Influence Ovarian Cancer Development Through Inflammation
3.4. Epigenetic Modifications by the Gut and Vaginal Microbiomes and Epigenetics in Ovarian Cancer
3.4.1. Epigenetic Modifications and Ovarian Cancer
3.4.2. Epigenetic Regulation by the Gut Microbiota in Ovarian Cancer
3.4.3. Epigenetic Regulation by the Vaginal Microbiota in Ovarian Cancer
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Laversanne, M.; Sung, H.; Ferlay, J.; Siegel, R.L.; Soerjomataram, I.; Jemal, A. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2024, 74, 229–263. [Google Scholar] [CrossRef]
- Torre, L.A.; Trabert, B.; DeSantis, C.E.; Miller, K.D.; Samimi, G.; Runowicz, C.D.; Gaudet, M.M.; Jemal, A.; Siegel, R.L. Ovarian cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 284–296. [Google Scholar] [CrossRef]
- Huang, J.; Chan, W.C.; Ngai, C.H.; Lok, V.; Zhang, L.; Lucero-Prisno, D.E.; Xu, W.; Zheng, Z.-J.; Elcarte, E.; Withers, M.; et al. Worldwide Burden, Risk Factors, and Temporal Trends of Ovarian Cancer: A Global Study. Cancers 2022, 14, 2230. [Google Scholar] [CrossRef]
- Liu, J.; Berchuck, A.; Backes, F.J.; Cohen, J.; Grisham, R.; Leath, C.A.; Martin, L.; Matei, D.; Miller, D.S.; Robertson, S.; et al. NCCN Guidelines® Insights: Ovarian Cancer/Fallopian Tube Cancer/Primary Peritoneal Cancer, Version 3.2024. J. Natl. Compr. Cancer Netw. 2024, 22, 512–519. [Google Scholar] [CrossRef] [PubMed]
- Gaillard, S.; Lacchetti, C.; Armstrong, D.K.; Cliby, W.A.; Edelson, M.I.; Garcia, A.A.; Ghebre, R.G.; Gressel, G.M.; Lesnock, J.L.; Meyer, L.A.; et al. Neoadjuvant Chemotherapy for Newly Diagnosed, Advanced Ovarian Cancer: ASCO Guideline Update. J. Clin. Oncol. 2025, 43, 868–891. [Google Scholar] [CrossRef]
- The Chicago Consensus on peritoneal surface malignancies: Management of ovarian neoplasms. Cancer 2020, 126, 2553–2560. [CrossRef] [PubMed]
- Webb, P.M.; Jordan, S.J. Global epidemiology of epithelial ovarian cancer. Nat. Rev. Clin. Oncol. 2024, 21, 389–400. [Google Scholar] [CrossRef] [PubMed]
- Bieging, K.T.; Mello, S.S.; Attardi, L.D. Unravelling mechanisms of p53-mediated tumour suppression. Nat. Rev. Cancer 2014, 14, 359–370. [Google Scholar] [CrossRef]
- Schmidt, S. The Tailpipe’s Tale: Traffic-Related Air Pollutants and Ovarian Cancer Risk. Environ. Health Perspect. 2024, 132, 124001. [Google Scholar] [CrossRef]
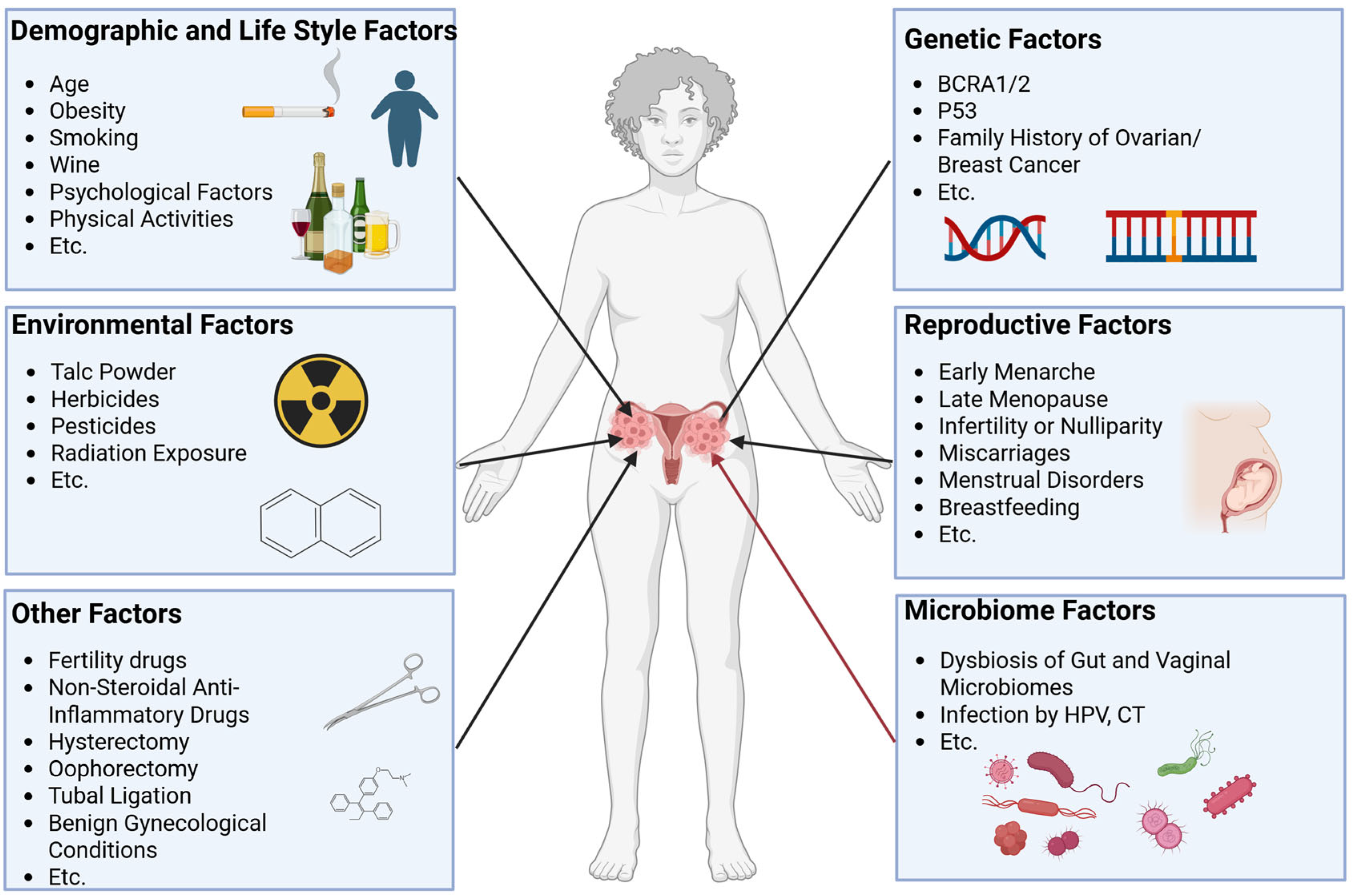
- Ali, A.T.; Al-Ani, O.; Al-Ani, F. Epidemiology and risk factors for ovarian cancer. Prz. Menopauzalny 2023, 22, 93–104. [Google Scholar] [CrossRef]
- La Vecchia, C. Ovarian cancer: Epidemiology and risk factors. Eur. J. Cancer Prev. 2017, 26, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S.; Oler, E.; Peters, H.; Guo, A.; Girod, S.; Han, S.; Saha, S.; Lui, V.W.; LeVatte, M.; Gautam, V.; et al. MiMeDB: The Human Microbial Metabolome Database. Nucleic Acids Res. 2023, 51, D611–D620. [Google Scholar] [CrossRef]
- Belkaid, Y.; Hand, T.W. Role of the microbiota in immunity and inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef] [PubMed]
- Postler, T.S.; Ghosh, S. Understanding the Holobiont: How Microbial Metabolites Affect Human Health and Shape the Immune System. Cell Metab. 2017, 26, 110–130. [Google Scholar] [CrossRef]
- Chen, Y.; Xiao, L.; Zhou, M.; Zhang, H. The microbiota: A crucial mediator in gut homeostasis and colonization resistance. Front. Microbiol. 2024, 15, 1417864. [Google Scholar] [CrossRef]
- Ma, C.; Jiang, M.; Li, J.; Zeng, Z.; Wu, Y.; Cheng, R.; Lin, H.; Pang, J.; Yin, F.; Jia, Y.; et al. Plasma Epstein-Barr Virus DNA load for diagnostic and prognostic assessment in intestinal Epstein-Barr Virus infection. Front. Cell. Infect. Microbiol. 2025, 14, 1526633. [Google Scholar] [CrossRef] [PubMed]
- You, L.; Zhou, J.; Xin, Z.; Hauck, J.S.; Na, F.; Tang, J.; Zhou, X.; Lei, Z.; Ying, B. Novel directions of precision oncology: Circulating microbial DNA emerging in cancer-microbiome areas. Precis. Clin. Med. 2022, 5, pbac005. [Google Scholar] [CrossRef]
- Rutkowski, M.R.; Stephen, T.L.; Svoronos, N.; Allegrezza, M.J.; Tesone, A.J.; Perales-Puchalt, A.; Brencicova, E.; Escovar-Fadul, X.; Nguyen, J.M.; Cadungog, M.G.; et al. Microbially driven TLR5-dependent signaling governs distal malignant progression through tumor-promoting inflammation. Cancer Cell 2015, 27, 27–40. [Google Scholar] [CrossRef]
- Nené, N.R.; Reisel, D.; Leimbach, A.; Franchi, D.; Jones, A.; Evans, I.; Knapp, S.; Ryan, A.; Ghazali, S.; Timms, J.F.; et al. Association between the cervicovaginal microbiome, BRCA1 mutation status, and risk of ovarian cancer: A case-control study. Lancet Oncol. 2019, 20, 1171–1182. [Google Scholar] [CrossRef]
- Jacobson, D.; Moore, K.; Gunderson, C.; Rowland, M.; Austin, R.; Honap, T.P.; Xu, J.; Warinner, C.; Sankaranarayanan, K.; Lewis, C.M. Shifts in gut and vaginal microbiomes are associated with cancer recurrence time in women with ovarian cancer. PeerJ 2021, 9, e11574. [Google Scholar] [CrossRef]
- Mungenast, F.; Thalhammer, T. Estrogen biosynthesis and action in ovarian cancer. Front. Endocrinol. 2014, 5, 192. [Google Scholar] [CrossRef]
- Borella, F.; Carosso, A.R.; Cosma, S.; Preti, M.; Collemi, G.; Cassoni, P.; Bertero, L.; Benedetto, C. Gut Microbiota and Gynecological Cancers: A Summary of Pathogenetic Mechanisms and Future Directions. ACS Infect. Dis. 2021, 7, 987–1009. [Google Scholar] [CrossRef] [PubMed]
- Matei, D.; Nephew, K.P. Epigenetic Attire in Ovarian Cancer: The Emperor’s New Clothes. Cancer Res. 2020, 80, 3775–3785. [Google Scholar] [CrossRef]
- Lozupone, C.A.; Stombaugh, J.I.; Gordon, J.I.; Jansson, J.K.; Knight, R. Diversity, stability and resilience of the human gut microbiota. Nature 2012, 489, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Kandari, A.; Odat, M.e.A.; Alzaid, F.; Scott, K.P. Biotics and bacterial function: Impact on gut and host health. ISME J. 2024, 18, wrae226. [Google Scholar] [CrossRef] [PubMed]
- Santos-Marcos, J.A.; Rangel-Zuñiga, O.A.; Jimenez-Lucena, R.; Quintana-Navarro, G.M.; Garcia-Carpintero, S.; Malagon, M.M.; Landa, B.B.; Tena-Sempere, M.; Perez-Martinez, P.; Lopez-Miranda, J.; et al. Influence of gender and menopausal status on gut microbiota. Maturitas 2018, 116, 43–53. [Google Scholar] [CrossRef]
- Dong, C.; Guan, Q.; Xu, W.; Zhang, X.; Jin, B.; Yu, S.; Xu, X.; Xia, Y. Disentangling the age-related manner in the associations between gut microbiome and women’s health: A multi-cohort microbiome study. Gut Microbes 2023, 15, 2290320. [Google Scholar] [CrossRef]
- Zhang, X.; Zhong, H.; Li, Y.; Shi, Z.; Ren, H.; Zhang, Z.; Zhou, X.; Tang, S.; Han, X.; Lin, Y.; et al. Sex- and age-related trajectories of the adult human gut microbiota shared across populations of different ethnicities. Nat. Aging 2021, 1, 87–100. [Google Scholar] [CrossRef]
- Mayneris-Perxachs, J.; Arnoriaga-Rodríguez, M.; Luque-Córdoba, D.; Priego-Capote, F.; Pérez-Brocal, V.; Moya, A.; Burokas, A.; Maldonado, R.; Fernández-Real, J.-M. Gut microbiota steroid sexual dimorphism and its impact on gonadal steroids: Influences of obesity and menopausal status. Microbiome 2020, 8, 136. [Google Scholar] [CrossRef]
- Renall, N.; Lawley, B.; Vatanen, T.; Merz, B.; Douwes, J.; Corbin, M.; Te Morenga, L.; Kruger, R.; Breier, B.H.; Tannock, G.W. The fecal microbiotas of women of Pacific and New Zealand European ethnicities are characterized by distinctive enterotypes that reflect dietary intakes and fecal water content. Gut Microbes 2023, 15, 2178801. [Google Scholar] [CrossRef]
- Zhong, H.; Penders, J.; Shi, Z.; Ren, H.; Cai, K.; Fang, C.; Ding, Q.; Thijs, C.; Blaak, E.E.; Stehouwer, C.D.A.; et al. Impact of early events and lifestyle on the gut microbiota and metabolic phenotypes in young school-age children. Microbiome 2019, 7, 2. [Google Scholar] [CrossRef] [PubMed]
- Anahtar, M.N.; Gootenberg, D.B.; Mitchell, C.M.; Kwon, D.S. Cervicovaginal Microbiota and Reproductive Health: The Virtue of Simplicity. Cell Host Microbe 2018, 23, 159–168. [Google Scholar] [CrossRef]
- France, M.; Alizadeh, M.; Brown, S.; Ma, B.; Ravel, J. Towards a deeper understanding of the vaginal microbiota. Nat. Microbiol. 2022, 7, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Gajer, P.; Brotman, R.M.; Bai, G.; Sakamoto, J.; Schütte, U.M.E.; Zhong, X.; Koenig, S.S.K.; Fu, L.; Ma, Z.S.; Zhou, X.; et al. Temporal dynamics of the human vaginal microbiota. Sci. Transl. Med. 2012, 4, 132ra152. [Google Scholar] [CrossRef] [PubMed]
- Ravel, J.; Gajer, P.; Abdo, Z.; Schneider, G.M.; Koenig, S.S.K.; McCulle, S.L.; Karlebach, S.; Gorle, R.; Russell, J.; Tacket, C.O.; et al. Vaginal microbiome of reproductive-age women. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. S1), 4680–4687. [Google Scholar] [CrossRef]
- Mancabelli, L.; Tarracchini, C.; Milani, C.; Lugli, G.A.; Fontana, F.; Turroni, F.; van Sinderen, D.; Ventura, M. Vaginotypes of the human vaginal microbiome. Environ. Microbiol. 2021, 23, 1780–1792. [Google Scholar] [CrossRef]
- Greenbaum, S.; Greenbaum, G.; Moran-Gilad, J.; Weintraub, A.Y. Ecological dynamics of the vaginal microbiome in relation to health and disease. Am. J. Obstet. Gynecol. 2019, 220, 324–335. [Google Scholar] [CrossRef]
- Champer, M.; Wong, A.M.; Champer, J.; Brito, I.L.; Messer, P.W.; Hou, J.Y.; Wright, J.D. The role of the vaginal microbiome in gynaecological cancer. BJOG Int. J. Obstet. Gynaecol. 2018, 125, 309–315. [Google Scholar] [CrossRef]
- Smith, S.B.; Ravel, J. The vaginal microbiota, host defence and reproductive physiology. J. Physiol. 2017, 595, 451–463. [Google Scholar] [CrossRef]
- Shen, L.; Zhang, W.; Yuan, Y.; Zhu, W.; Shang, A. Vaginal microecological characteristics of women in different physiological and pathological period. Front. Cell. Infect. Microbiol. 2022, 12, 959793. [Google Scholar] [CrossRef]
- Asangba, A.E.; Chen, J.; Goergen, K.M.; Larson, M.C.; Oberg, A.L.; Casarin, J.; Multinu, F.; Kaufmann, S.H.; Mariani, A.; Chia, N.; et al. Diagnostic and prognostic potential of the microbiome in ovarian cancer treatment response. Sci. Rep. 2023, 13, 730. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Xu, X.; Zeng, X.; Jin, R.; Wang, S.; Jiang, H.; Tang, Y.; Chen, G.; Wei, J.; Chen, T.; et al. Gut microbiota dysbiosis promotes the development of epithelial ovarian cancer via regulating Hedgehog signaling pathway. Gut Microbes 2023, 15, 2221093. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, F.; Perrone, A.M.; Rampelli, S.; Coluccelli, S.; Barone, M.; Ravegnini, G.; Fabbrini, M.; Brigidi, P.; De Iaco, P.; Turroni, S. Gut Microbiota Dynamics during Chemotherapy in Epithelial Ovarian Cancer Patients Are Related to Therapeutic Outcome. Cancers 2021, 13, 3999. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Deng, C.; Li, Y.; He, S.; Liu, Y.; Pan, S.; Xu, W.; Fang, L.; Zhu, Y.; Wang, Y.; et al. Machine learning-derived diagnostic model of epithelial ovarian cancer based on gut microbiome signatures. J. Transl. Med. 2025, 23, 319. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, X.-R.; Park, J.-L.; Kim, J.-H.; Zhang, L.; Ma, J.-L.; Liu, W.-D.; Deng, D.-J.; You, W.-C.; Kim, Y.-S.; et al. Genome-wide DNA methylation profiles altered by Helicobacter pylori in gastric mucosa and blood leukocyte DNA. Oncotarget 2016, 7, 37132–37144. [Google Scholar] [CrossRef]
- Trabert, B.; Waterboer, T.; Idahl, A.; Brenner, N.; Brinton, L.A.; Butt, J.; Coburn, S.B.; Hartge, P.; Hufnagel, K.; Inturrisi, F.; et al. Antibodies Against Chlamydia trachomatis and Ovarian Cancer Risk in Two Independent Populations. J. Natl. Cancer Inst. 2019, 111, 129–136. [Google Scholar] [CrossRef]
- Sharma, H.; Tal, R.; Clark, N.A.; Segars, J.H. Microbiota and pelvic inflammatory disease. Semin. Reprod. Med. 2014, 32, 43–49. [Google Scholar] [CrossRef]
- Antonio, M.A.D.; Rabe, L.K.; Hillier, S.L. Colonization of the rectum by Lactobacillus species and decreased risk of bacterial vaginosis. J. Infect. Dis. 2005, 192, 394–398. [Google Scholar] [CrossRef]
- El Aila, N.A.; Tency, I.; Claeys, G.; Verstraelen, H.; Saerens, B.; Santiago, G.L.D.S.; De Backer, E.; Cools, P.; Temmerman, M.; Verhelst, R.; et al. Identification and genotyping of bacteria from paired vaginal and rectal samples from pregnant women indicates similarity between vaginal and rectal microflora. BMC Infect. Dis. 2009, 9, 167. [Google Scholar] [CrossRef]
- Shin, H.; Martinez, K.A.; Henderson, N.; Jay, M.; Schweizer, W.; Bogaert, D.; Park, G.; Bokulich, N.A.; Blaser, M.J.; Dominguez-Bello, M.G. Partial convergence of the human vaginal and rectal maternal microbiota in late gestation and early post-partum. npj Biofilms Microbiomes 2023, 9, 37. [Google Scholar] [CrossRef]
- Fudaba, M.; Kamiya, T.; Tachibana, D.; Koyama, M.; Ohtani, N. Bioinformatics Analysis of Oral, Vaginal, and Rectal Microbial Profiles during Pregnancy: A Pilot Study on the Bacterial Co-Residence in Pregnant Women. Microorganisms 2021, 9, 1027. [Google Scholar] [CrossRef] [PubMed]
- Di Pierro, F.; Criscuolo, A.A.; Dei Giudici, A.; Senatori, R.; Sesti, F.; Ciotti, M.; Piccione, E. Oral administration of Lactobacillus crispatus M247 to papillomavirus-infected women: Results of a preliminary, uncontrolled, open trial. Minerva Obstet. Gynecol. 2021, 73, 621–631. [Google Scholar] [CrossRef]
- Jang, S.-E.; Jeong, J.-J.; Choi, S.-Y.; Kim, H.; Han, M.J.; Kim, D.-H. Lactobacillus rhamnosus HN001 and Lactobacillus acidophilus La-14 Attenuate Gardnerella vaginalis-Infected Bacterial Vaginosis in Mice. Nutrients 2017, 9, 531. [Google Scholar] [CrossRef]
- Yuan, C.; Gaskins, A.J.; Blaine, A.I.; Zhang, C.; Gillman, M.W.; Missmer, S.A.; Field, A.E.; Chavarro, J.E. Association Between Cesarean Birth and Risk of Obesity in Offspring in Childhood, Adolescence, and Early Adulthood. JAMA Pediatr. 2016, 170, e162385. [Google Scholar] [CrossRef]
- Kristensen, K.; Henriksen, L. Cesarean section and disease associated with immune function. J. Allergy Clin. Immunol. 2016, 137, 587–590. [Google Scholar] [CrossRef]
- Plottel, C.S.; Blaser, M.J. Microbiome and malignancy. Cell Host Microbe 2011, 10, 324–335. [Google Scholar] [CrossRef] [PubMed]
- Kwa, M.; Plottel, C.S.; Blaser, M.J.; Adams, S. The Intestinal Microbiome and Estrogen Receptor-Positive Female Breast Cancer. J. Natl. Cancer Inst. 2016, 108, djw029. [Google Scholar] [CrossRef] [PubMed]
- Ervin, S.M.; Li, H.; Lim, L.; Roberts, L.R.; Liang, X.; Mani, S.; Redinbo, M.R. Gut microbial β-glucuronidases reactivate estrogens as components of the estrobolome that reactivate estrogens. J. Biol. Chem. 2019, 294, 18586–18599. [Google Scholar] [CrossRef]
- Buchta, V. Vaginal microbiome. Ces. Gynekol. 2018, 83, 371–379. [Google Scholar]
- Claes, I.J.; Vargas García, C.E.; Lebeer, S. Novel opportunities for the exploitation of host-microbiome interactions in the intestine. Curr. Opin. Biotechnol. 2015, 32, 28–34. [Google Scholar] [CrossRef]
- Favaron, A.; Turkgeldi, E.; Elbadawi, M.; Gaisford, S.; Basit, A.W.; Orlu, M. Do probiotic interventions improve female unexplained infertility? A critical commentary. Reprod. Biomed. Online 2024, 48, 103734. [Google Scholar] [CrossRef]
- Miko, E.; Barakonyi, A. The Role of Hydrogen-Peroxide (H2O2) Produced by Vaginal Microbiota in Female Reproductive Health. Antioxidants 2023, 12, 1055. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, H.-T.; Zhou, S.-J.; Zhou, H.-H.; Xiong, Y.; Yang, J.; Zhou, Y.-B.; Chen, D.-J.; Liu, J.-M. Effects of vaginal seeding on gut microbiota, body mass index, and allergy risks in infants born through cesarean delivery: A randomized clinical trial. Am. J. Obstet. Gynecol. MFM 2023, 5, 100793. [Google Scholar] [CrossRef]
- Zhao, F.; Hu, X.; Ying, C. Advances in Research on the Relationship between Vaginal Microbiota and Adverse Pregnancy Outcomes and Gynecological Diseases. Microorganisms 2023, 11, 991. [Google Scholar] [CrossRef] [PubMed]
- Bar, O.; Sudhof, L.S.; Yockey, L.J.; Bergerat, A.; Moriel, N.; Andrews, E.; Ananthakrishnan, A.N.; Xavier, R.J.; Yassour, M.; Mitchell, C.M. Comparison of vaginal microbiota between women with inflammatory bowel disease and healthy controls. PLoS ONE 2023, 18, e0284709. [Google Scholar] [CrossRef]
- Wen, Q.; Wang, S.; Min, Y.; Liu, X.; Fang, J.; Lang, J.; Chen, M. Associations of the gut, cervical, and vaginal microbiota with cervical cancer: A systematic review and meta-analysis. BMC Womens Health 2025, 25, 65. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Yi, Z.; Jiao, Y.; Shen, Z.; Yang, F.; Zhu, S. Gut Microbiota and Adipose Tissue Microenvironment Interactions in Obesity. Metabolites 2023, 13, 821. [Google Scholar] [CrossRef]
- Yoon, K.; Kim, N. Roles of Sex Hormones and Gender in the Gut Microbiota. J. Neurogastroenterol. Motil. 2021, 27, 314–325. [Google Scholar] [CrossRef]
- Raftogianis, R.; Creveling, C.; Weinshilboum, R.; Weisz, J. Estrogen metabolism by conjugation. J. Natl. Cancer Inst. Monogr. 2000, 2000, 113–124. [Google Scholar] [CrossRef]
- Ruggiero, R.J.; Likis, F.E. Estrogen: Physiology, pharmacology, and formulations for replacement therapy. J. Midwifery Womens Health 2002, 47, 130–138. [Google Scholar] [CrossRef]
- Cui, J.; Shen, Y.; Li, R. Estrogen synthesis and signaling pathways during aging: From periphery to brain. Trends Mol. Med. 2013, 19, 197–209. [Google Scholar] [CrossRef]
- Prossnitz, E.R.; Barton, M. The G protein-coupled oestrogen receptor GPER in health and disease: An update. Nat. Rev. Endocrinol. 2023, 19, 407–424. [Google Scholar] [CrossRef] [PubMed]
- Seyed Hameed, A.S.; Rawat, P.S.; Meng, X.; Liu, W. Biotransformation of dietary phytoestrogens by gut microbes: A review on bidirectional interaction between phytoestrogen metabolism and gut microbiota. Biotechnol. Adv. 2020, 43, 107576. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Lee, B.; Choi, H.; Kim, T.; Kim, Y.; Kim, Y.B. Impact of Hormone Replacement Therapy on Risk of Ovarian Cancer in Postmenopausal Women with De Novo Endometriosis or a History of Endometriosis. Cancers 2023, 15, 1708. [Google Scholar] [CrossRef]
- Petrick, J.L.; Joslin, C.E.; Johnson, C.E.; Camacho, T.F.; Peres, L.C.; Bandera, E.V.; Barnard, M.E.; Beeghly, A.; Bethea, T.N.; Dempsey, L.F.; et al. Menopausal hormone therapy use and risk of ovarian cancer by race: The ovarian cancer in women of African ancestry consortium. Br. J. Cancer 2023, 129, 1956–1967. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.W.; Wu, A.H.; Wiensch, A.; Mukherjee, B.; Terry, K.L.; Harris, H.R.; Carney, M.E.; Jensen, A.; Cramer, D.W.; Berchuck, A.; et al. Estrogen Plus Progestin Hormone Therapy and Ovarian Cancer: A Complicated Relationship Explored. Epidemiology 2020, 31, 402–408. [Google Scholar] [CrossRef]
- Johansson, Å.; Schmitz, D.; Höglund, J.; Hadizadeh, F.; Karlsson, T.; Ek, W.E. Investigating the Effect of Estradiol Levels on the Risk of Breast, Endometrial, and Ovarian Cancer. J. Endocr. Soc. 2022, 6, bvac100. [Google Scholar] [CrossRef]
- Cunat, S.; Hoffmann, P.; Pujol, P. Estrogens and epithelial ovarian cancer. Gynecol. Oncol. 2004, 94, 25–32. [Google Scholar] [CrossRef]
- Liu, A.; Zhang, D.; Yang, X.; Song, Y. Estrogen receptor alpha activates MAPK signaling pathway to promote the development of endometrial cancer. J. Cell. Biochem. 2019, 120, 17593–17601. [Google Scholar] [CrossRef]
- Goad, J.; Ko, Y.-A.; Kumar, M.; Jamaluddin, M.F.B.; Tanwar, P.S. Oestrogen fuels the growth of endometrial hyperplastic lesions initiated by overactive Wnt/β-catenin signalling. Carcinogenesis 2018, 39, 1105–1116. [Google Scholar] [CrossRef]
- Liu, X.; Zhan, T.; Gao, Y.; Cui, S.; Liu, W.; Zhang, C.; Zhuang, S. Benzophenone-1 induced aberrant proliferation and metastasis of ovarian cancer cells via activated ERα and Wnt/β-catenin signaling pathways. Environ. Pollut. 2022, 292, 118370. [Google Scholar] [CrossRef]
- Simpkins, F.; Jang, K.; Yoon, H.; Hew, K.E.; Kim, M.; Azzam, D.J.; Sun, J.; Zhao, D.; Ince, T.A.; Liu, W.; et al. Dual Src and MEK Inhibition Decreases Ovarian Cancer Growth and Targets Tumor Initiating Stem-like Cells. Clin. Cancer Res. 2018, 24, 4874–4886. [Google Scholar] [CrossRef] [PubMed]
- Petrie, W.K.; Dennis, M.K.; Hu, C.; Dai, D.; Arterburn, J.B.; Smith, H.O.; Hathaway, H.J.; Prossnitz, E.R. G protein-coupled estrogen receptor-selective ligands modulate endometrial tumor growth. Obstet. Gynecol. Int. 2013, 2013, 472720. [Google Scholar] [CrossRef]
- Pollet, R.M.; D’Agostino, E.H.; Walton, W.G.; Xu, Y.; Little, M.S.; Biernat, K.A.; Pellock, S.J.; Patterson, L.M.; Creekmore, B.C.; Isenberg, H.N.; et al. An Atlas of β-Glucuronidases in the Human Intestinal Microbiome. Structure 2017, 25, 967–977.E5. [Google Scholar] [CrossRef]
- Tetel, M.J.; de Vries, G.J.; Melcangi, R.C.; Panzica, G.; O’Mahony, S.M. Steroids, stress and the gut microbiome-brain axis. J. Neuroendocrinol. 2018, 30, e12548. [Google Scholar] [CrossRef] [PubMed]
- Maeng, L.Y.; Beumer, A. Never fear, the gut bacteria are here: Estrogen and gut microbiome-brain axis interactions in fear extinction. Int. J. Psychophysiol. 2023, 189, 66–75. [Google Scholar] [CrossRef]
- Dinan, T.G.; Scott, L.V. Anatomy of melancholia: Focus on hypothalamic-pituitary-adrenal axis overactivity and the role of vasopressin. J. Anat. 2005, 207, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Oyola, M.G.; Handa, R.J. Hypothalamic-pituitary-adrenal and hypothalamic-pituitary-gonadal axes: Sex differences in regulation of stress responsivity. Stress 2017, 20, 476–494. [Google Scholar] [CrossRef]
- de Visser, K.E.; Joyce, J.A. The evolving tumor microenvironment: From cancer initiation to metastatic outgrowth. Cancer Cell 2023, 41, 374–403. [Google Scholar] [CrossRef]
- Zhao, H.; Wu, L.; Yan, G.; Chen, Y.; Zhou, M.; Wu, Y.; Li, Y. Inflammation and tumor progression: Signaling pathways and targeted intervention. Signal Transduct. Target. Ther. 2021, 6, 263. [Google Scholar] [CrossRef]
- Hu, Y.; Wang, W.; Ma, W.; Wang, W.; Ren, W.; Wang, S.; Fu, F.; Li, Y. Impact of psychological stress on ovarian function: Insights, mechanisms and intervention strategies (Review). Int. J. Mol. Med. 2025, 55, 34. [Google Scholar] [CrossRef] [PubMed]
- Huldani, H.; Abdul-Jabbar Ali, S.; Al-Dolaimy, F.; Hjazi, A.; Denis Andreevich, N.; Oudaha, K.H.; Almulla, A.F.; Alsaalamy, A.; Kareem Oudah, S.; Mustafa, Y.F. The potential role of interleukins and interferons in ovarian cancer. Cytokine 2023, 171, 156379. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Xu, K.; Liu, H.; Liu, G.; Bai, M.; Peng, C.; Li, T.; Yin, Y. Impact of the Gut Microbiota on Intestinal Immunity Mediated by Tryptophan Metabolism. Front. Cell. Infect. Microbiol. 2018, 8, 13. [Google Scholar] [CrossRef]
- Fachi, J.L.; Sécca, C.; Rodrigues, P.B.; Mato, F.C.P.d.; Di Luccia, B.; Felipe, J.d.S.; Pral, L.P.; Rungue, M.; Rocha, V.d.M.; Sato, F.T.; et al. Acetate coordinates neutrophil and ILC3 responses against C. difficile through FFAR2. J. Exp. Med. 2020, 217, e20190489. [Google Scholar] [CrossRef] [PubMed]
- Woods, D.C.; White, Y.A.R.; Dau, C.; Johnson, A.L. TLR4 activates NF-κB in human ovarian granulosa tumor cells. Biochem. Biophys. Res. Commun. 2011, 409, 675–680. [Google Scholar] [CrossRef]
- Kelly, M.G.; Alvero, A.B.; Chen, R.; Silasi, D.-A.; Abrahams, V.M.; Chan, S.; Visintin, I.; Rutherford, T.; Mor, G. TLR-4 signaling promotes tumor growth and paclitaxel chemoresistance in ovarian cancer. Cancer Res. 2006, 66, 3859–3868. [Google Scholar] [CrossRef]
- Lupi, L.A.; Cucielo, M.S.; Silveira, H.S.; Gaiotte, L.B.; Cesário, R.C.; Seiva, F.R.F.; de Almeida Chuffa, L.G. The role of Toll-like receptor 4 signaling pathway in ovarian, cervical, and endometrial cancers. Life Sci. 2020, 247, 117435. [Google Scholar] [CrossRef]
- Kasperczyk, H.; Baumann, B.; Debatin, K.-M.; Fulda, S. Characterization of sonic hedgehog as a novel NF-kappaB target gene that promotes NF-kappaB-mediated apoptosis resistance and tumor growth in vivo. FASEB J. 2009, 23, 21–33. [Google Scholar] [CrossRef]
- Wang, Y.; Jin, G.; Li, Q.; Wang, Z.; Hu, W.; Li, P.; Li, S.; Wu, H.; Kong, X.; Gao, J.; et al. Hedgehog Signaling Non-Canonical Activated by Pro-Inflammatory Cytokines in Pancreatic Ductal Adenocarcinoma. J. Cancer 2016, 7, 2067–2076. [Google Scholar] [CrossRef]
- Browning, L.; Patel, M.R.; Horvath, E.B.; Tawara, K.; Jorcyk, C.L. IL-6 and ovarian cancer: Inflammatory cytokines in promotion of metastasis. Cancer Manag. Res. 2018, 10, 6685–6693. [Google Scholar] [CrossRef]
- Horowitz, N.S.; Hua, J.; Powell, M.A.; Gibb, R.K.; Mutch, D.G.; Herzog, T.J. Novel cytotoxic agents from an unexpected source: Bile acids and ovarian tumor apoptosis. Gynecol. Oncol. 2007, 107, 344–349. [Google Scholar] [CrossRef]
- Sipos, A.; Kerekes, É.; Szeőcs, D.; Szarvas, F.; Schwarcz, S.; Tóth, E.; Ujlaki, G.; Mikó, E.; Bai, P. Ursodeoxycholic acid prompts glycolytic dominance, reductive stress and epithelial-to-mesenchymal transition in ovarian cancer cells through NRF2 activation. Cell Death Discov. 2025, 11, 134. [Google Scholar] [CrossRef]
- Sun, M.; Wu, W.; Liu, Z.; Cong, Y. Microbiota metabolite short chain fatty acids, GPCR, and inflammatory bowel diseases. J. Gastroenterol. 2017, 52, 1–8. [Google Scholar] [CrossRef]
- Zhao, L.-Y.; Mei, J.-X.; Yu, G.; Lei, L.; Zhang, W.-H.; Liu, K.; Chen, X.-L.; Kołat, D.; Yang, K.; Hu, J.-K. Role of the gut microbiota in anticancer therapy: From molecular mechanisms to clinical applications. Signal Transduct. Target. Ther. 2023, 8, 201. [Google Scholar] [CrossRef] [PubMed]
- Tossetta, G.; Marzioni, D. Natural and synthetic compounds in Ovarian Cancer: A focus on NRF2/KEAP1 pathway. Pharmacol. Res. 2022, 183, 106365. [Google Scholar] [CrossRef] [PubMed]
- Long, Y.; Shi, H.; Ye, J.; Qi, X. Exploring Strategies to Prevent and Treat Ovarian Cancer in Terms of Oxidative Stress and Antioxidants. Antioxidants 2025, 14, 114. [Google Scholar] [CrossRef] [PubMed]
- Puschhof, J.; Sears, C.L. Microbial metabolites damage DNA. Science 2022, 378, 358–359. [Google Scholar] [CrossRef]
- Mostafavi Abdolmaleky, H.; Zhou, J.-R. Gut Microbiota Dysbiosis, Oxidative Stress, Inflammation, and Epigenetic Alterations in Metabolic Diseases. Antioxidants 2024, 13, 985. [Google Scholar] [CrossRef]
- Rahbar Saadat, Y.; Pourseif, M.M.; Zununi Vahed, S.; Barzegari, A.; Omidi, Y.; Barar, J. Modulatory Role of Vaginal-Isolated Lactococcus lactis on the Expression of miR-21, miR-200b, and TLR-4 in CAOV-4 Cells and In Silico Revalidation. Probiotics Antimicrob. Proteins 2020, 12, 1083–1096. [Google Scholar] [CrossRef]
- Pathak, S.; Wilczyński, J.R.; Paradowska, E. Factors in Oncogenesis: Viral Infections in Ovarian Cancer. Cancers 2020, 12, 561. [Google Scholar] [CrossRef]
- Xie, X.; Yang, M.; Ding, Y.; Chen, J. Microbial infection, inflammation and epithelial ovarian cancer. Oncol. Lett. 2017, 14, 1911–1919. [Google Scholar] [CrossRef]
- Sipos, A.; Ujlaki, G.; Mikó, E.; Maka, E.; Szabó, J.; Uray, K.; Krasznai, Z.; Bai, P. The role of the microbiome in ovarian cancer: Mechanistic insights into oncobiosis and to bacterial metabolite signaling. Mol. Med. 2021, 27, 33. [Google Scholar] [CrossRef]
- Qazi, S.; Raza, K. In silico approach to understand epigenetics of POTEE in ovarian cancer. J. Integr. Bioinform. 2021, 18, 20210028. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ali, A.; Xie, J. Detection of clinically important BRCA gene mutations in ovarian cancer patients using next generation sequencing analysis. Am. J. Cancer Res. 2023, 13, 5005–5020. [Google Scholar]
- Fu, M.; Deng, F.; Chen, J.; Fu, L.; Lei, J.; Xu, T.; Chen, Y.; Zhou, J.; Gao, Q.; Ding, H. Current data and future perspectives on DNA methylation in ovarian cancer (Review). Int. J. Oncol. 2024, 64, 62. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, F.; Zheng, J.; Jiang, K.; Ai, H.; Liu, L.; Mao, D. MAPRE3 as an epigenetic target of EZH2 restricts ovarian cancer proliferation in vitro and in vivo. Exp. Cell Res. 2024, 435, 113913. [Google Scholar] [CrossRef]
- Wang, J.; Zhong, F.; Li, J.; Yue, H.; Li, W.; Lu, X. The epigenetic factor CHD4 contributes to metastasis by regulating the EZH2/β-catenin axis and acts as a therapeutic target in ovarian cancer. J. Transl. Med. 2023, 21, 38. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Li, W.; Guo, J.; Liu, L.; Wang, Y. Development and Validation of Prognostic Characteristics Associated With Chromatin Remodeling-Related Genes in Ovarian Cancer. Cancer Med. 2025, 14, e70634. [Google Scholar] [CrossRef] [PubMed]
- González-Cantó, E.; Monteiro, M.; Aghababyan, C.; Ferrero-Micó, A.; Navarro-Serna, S.; Mellado-López, M.; Tomás-Pérez, S.; Sandoval, J.; Llueca, A.; Herreros-Pomares, A.; et al. Reduced Levels of miR-145-3p Drive Cell Cycle Progression in Advanced High-Grade Serous Ovarian Cancer. Cells 2024, 13, 1904. [Google Scholar] [CrossRef]
- Zhang, Y.; Ling, Y.; Zhou, Y.; Shi, X.; Shen, F.; Zhou, J.; Chen, Y.; Yang, F.; Gu, Y.; Wang, J. Research Advances in the Roles of N6-Methyladenosine Modification in Ovarian Cancer. Cancer Control 2024, 31, 10732748241256819. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, C.; Goel, A. Role of gut microbiota in epigenetic regulation of colorectal Cancer. Biochim. Biophys. Acta Rev. Cancer 2021, 1875, 188490. [Google Scholar] [CrossRef]
- Li, D.; Li, Y.; Yang, S.; Lu, J.; Jin, X.; Wu, M. Diet-gut microbiota-epigenetics in metabolic diseases: From mechanisms to therapeutics. Biomed. Pharmacother. 2022, 153, 113290. [Google Scholar] [CrossRef] [PubMed]
- Woo, V.; Alenghat, T. Epigenetic regulation by gut microbiota. Gut Microbes 2022, 14, 2022407. [Google Scholar] [CrossRef]
- Queen, J.; Shaikh, F.; Sears, C.L. Understanding the mechanisms and translational implications of the microbiome for cancer therapy innovation. Nat. Cancer 2023, 4, 1083–1094. [Google Scholar] [CrossRef] [PubMed]
- Pepke, M.L.; Hansen, S.B.; Limborg, M.T. Unraveling host regulation of gut microbiota through the epigenome-microbiome axis. Trends Microbiol. 2024, 32, 1229–1240. [Google Scholar] [CrossRef]
- Xu, M.; Guo, Y.; Wang, F.; Lin, C.; Cao, D.; Yan, Y.; Chai, S.; Zhao, Y.; Deng, S.; Wei, J.; et al. Enterolactone combined with m6A Reader IGF2BP3 inhibits malignant angiogenesis and disease progression in ovarian cancer. Phytomedicine 2025, 136, 156343. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Li, Z.; Zhang, M. Emerging roles of intratumor microbiota in cancer: Tumorigenesis and management strategies. J. Transl. Med. 2024, 22, 837. [Google Scholar] [CrossRef]
- Owens, F.; Souchak, J.; Nazaire, V.; Akkaoui, J.; Shil, R.; Carbajal, C.; Panda, K.; Delgado, D.C.; Claassen, I.; Moreno, S.; et al. A Connection Between the Gut Microbiome and Epigenetic Modification in Age-Related Cancer: A Narrative Review. Aging Dis. 2025, 17, 2. [Google Scholar] [CrossRef]
- Xu, Y.; He, C.; Xi, Y.; Zhang, Y.; Bai, Y. Gut microbiota and immunosenescence in cancer. Semin. Cancer Biol. 2024, 104–105, 32–45. [Google Scholar] [CrossRef]
- Chen, L.; Zhai, Y.; Wang, Y.; Fearon, E.R.; Núñez, G.; Inohara, N.; Cho, K.R. Altering the Microbiome Inhibits Tumorigenesis in a Mouse Model of Oviductal High-Grade Serous Carcinoma. Cancer Res. 2021, 81, 3309–3318. [Google Scholar] [CrossRef]
- Wahid, M.; Dar, S.A.; Jawed, A.; Mandal, R.K.; Akhter, N.; Khan, S.; Khan, F.; Jogaiah, S.; Rai, A.K.; Rattan, R. Microbes in gynecologic cancers: Causes or consequences and therapeutic potential. Semin. Cancer Biol. 2022, 86 Pt 2, 1179–1189. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Feng, Y.; Yang, C.; Wang, D.; Zhang, D.; Luo, X.; Zhang, H.; Huang, H.; Zhang, H.; Jiang, Y.; et al. Association between vaginal microbiota and the progression of ovarian cancer. J. Med. Virol. 2023, 95, e28898. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.-L.; Xu, X.-Q.; Liu, X.-L.; Guo, Q.-Q.; Fan, Y.-N.; He, B.-X.; Zhang, W.-Z. Emerging role of m6A methylation modification in ovarian cancer. Cancer Cell Int. 2021, 21, 663. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, N.D.; Hong, H.; Ahmad, S.; Holloway, R.W. The gut microbiome and cancer immunotherapeutics: A review of emerging data and implications for future gynecologic cancer research. Crit. Rev. Oncol. Hematol. 2021, 157, 103165. [Google Scholar] [CrossRef]
- Wu, J.; Li, J.; Yan, M.; Xiang, Z. Gut and oral microbiota in gynecological cancers: Interaction, mechanism, and therapeutic value. npj Biofilms Microbiomes 2024, 10, 104. [Google Scholar] [CrossRef]
- Scott, A.J.; Alexander, J.L.; Merrifield, C.A.; Cunningham, D.; Jobin, C.; Brown, R.; Alverdy, J.; O’Keefe, S.J.; Gaskins, H.R.; Teare, J.; et al. International Cancer Microbiome Consortium consensus statement on the role of the human microbiome in carcinogenesis. Gut 2019, 68, 1624–1632. [Google Scholar] [CrossRef]
- Chen, J.; Chen, X.; Ma, J. Causal relationships of gut microbiota and blood metabolites with ovarian cancer and endometrial cancer: A Mendelian randomization study. J. Ovarian Res. 2025, 18, 54. [Google Scholar] [CrossRef]
- Zhang, L.; Cao, T.; Liu, K.; Sun, P.; Wang, W.; Guo, J. Genetically predicted blood metabolites mediate relationships between gut microbiota and ovarian cancer: A Mendelian randomization study. Front. Cell. Infect. Microbiol. 2024, 14, 1451880. [Google Scholar] [CrossRef]
- Wang, Y.; Gao, S.; Liu, Y.; Li, Y.; Yao, H.; Han, Y.; Liu, X. Association between gut microbiota, plasma metabolites, and ovarian cancer: A Mendelian randomization study. Medicine 2024, 103, e40479. [Google Scholar] [CrossRef]
- Webb, P.M.; Jordan, S.J. Epidemiology of epithelial ovarian cancer. Best Pract. Res. Clin. Obstet. Gynaecol. 2017, 41, 3–14. [Google Scholar] [CrossRef]
- Zeng, S.; Wang, X.L.; Yang, H. Radiomics and radiogenomics: Extracting more information from medical images for the diagnosis and prognostic prediction of ovarian cancer. Mil. Med. Res. 2024, 11, 77. [Google Scholar] [CrossRef] [PubMed]
- Cai, G.; Huang, F.; Gao, Y.; Li, X.; Chi, J.; Xie, J.; Zhou, L.; Feng, Y.; Huang, H.; Deng, T.; et al. Artificial intelligence-based models enabling accurate diagnosis of ovarian cancer using laboratory tests in China: A multicentre, retrospective cohort study. Lancet Digit. Health 2024, 6, e176–e186. [Google Scholar] [CrossRef] [PubMed]
- Vergote, I.; Gonzalez-Martin, A.; Lorusso, D.; Gourley, C.; Mirza, M.R.; Kurtz, J.E.; Okamoto, A.; Moore, K.; Kridelka, F.; McNeish, I.; et al. Clinical research in ovarian cancer: Consensus recommendations from the Gynecologic Cancer InterGroup. Lancet Oncol. 2022, 23, e374–e384. [Google Scholar] [CrossRef] [PubMed]
- Tung, N.; Ricker, C.; Messersmith, H.; Balmaña, J.; Domchek, S.; Stoffel, E.M.; Almhanna, K.; Arun, B.; Chavarri-Guerra, Y.; Cohen, S.A.; et al. Selection of Germline Genetic Testing Panels in Patients with Cancer: ASCO Guideline. J. Clin. Oncol. 2024, 42, 2599–2615. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Study | Study Design | Participants | Associated Outcomes |
|---|---|---|---|
| Nené 2019 [19] | Case–control study |
| Vaginal microbiota:
|
| Jacobson 2021 [20] | Cohort Study |
| Gut microbiota:
|
| Asangba 2023 [41] | Cohort study |
| Vaginal microbiota:
|
| Hu 2023 [42] | Case–control study |
| Gut microbiota:
|
| D’Amico 2021 [43] | Cohort study |
| Gut microbiota:
|
| Chen C. 2025 [44] | Case–control study |
| Gut microbiota:
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, H.; Zeng, Z.; Zhang, H.; Jia, Y.; Pang, J.; Chen, J.; Zhang, H. Gut–Vaginal Microbiome Crosstalk in Ovarian Cancer: Implications for Early Diagnosis. Pathogens 2025, 14, 635. https://doi.org/10.3390/pathogens14070635
Lin H, Zeng Z, Zhang H, Jia Y, Pang J, Chen J, Zhang H. Gut–Vaginal Microbiome Crosstalk in Ovarian Cancer: Implications for Early Diagnosis. Pathogens. 2025; 14(7):635. https://doi.org/10.3390/pathogens14070635
Chicago/Turabian StyleLin, Hao, Zhen Zeng, Hong Zhang, Yongbin Jia, Jiangmei Pang, Jingjing Chen, and Hu Zhang. 2025. "Gut–Vaginal Microbiome Crosstalk in Ovarian Cancer: Implications for Early Diagnosis" Pathogens 14, no. 7: 635. https://doi.org/10.3390/pathogens14070635
APA StyleLin, H., Zeng, Z., Zhang, H., Jia, Y., Pang, J., Chen, J., & Zhang, H. (2025). Gut–Vaginal Microbiome Crosstalk in Ovarian Cancer: Implications for Early Diagnosis. Pathogens, 14(7), 635. https://doi.org/10.3390/pathogens14070635