
Pathogens Detected in Questing Ixodes ricinus Ticks in a Mountainous Area in Greece

Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sampling
2.3. Tick Identification and Transportation
2.4. Nucleic Acid Isolation and Pathogen Detection
2.5. Sanger Sequencing and Phylogenetic Analysis
3. Results
3.1. Ticks
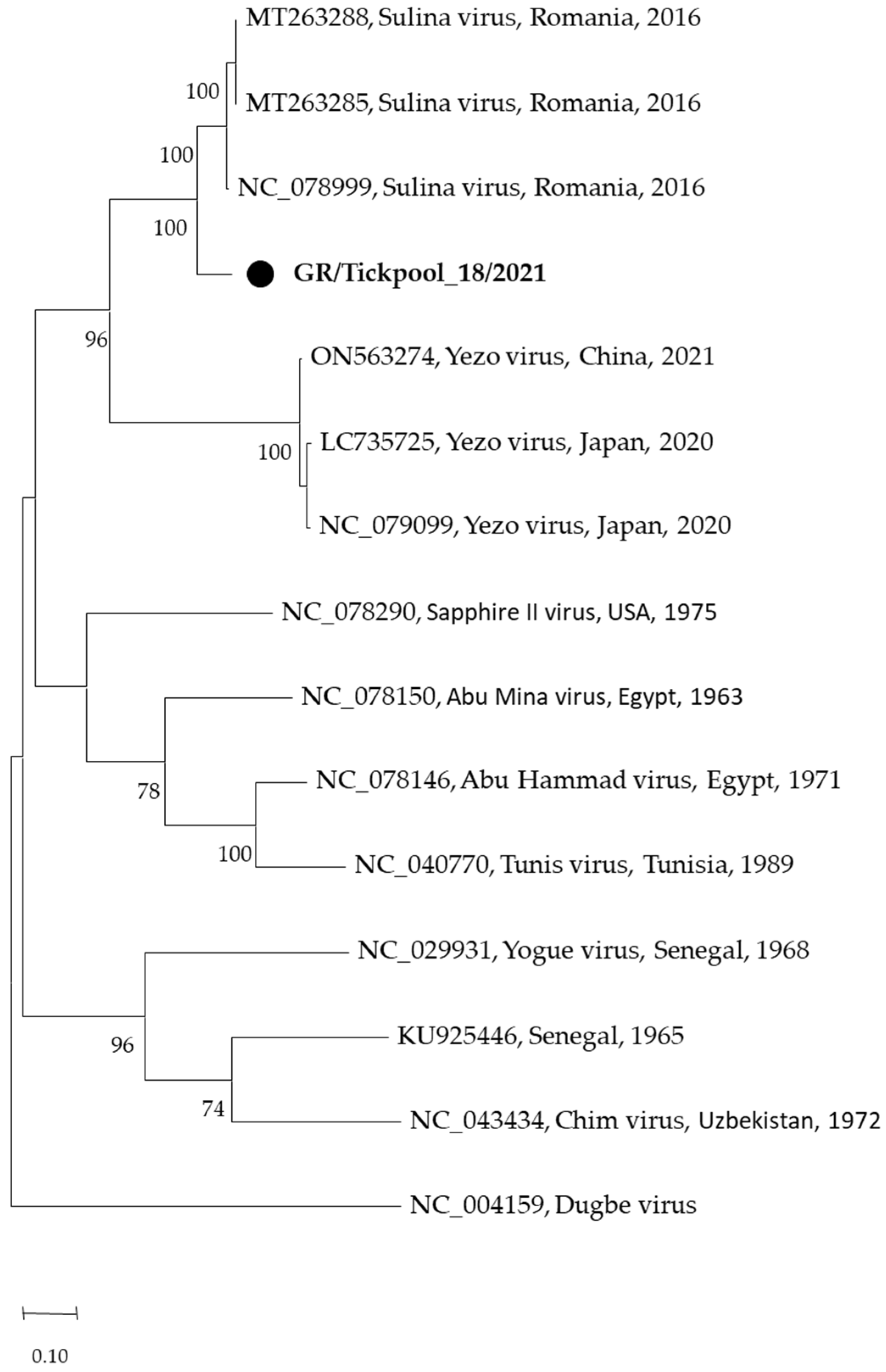
3.2. Pathogen Detection
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gray, J.S.; Kahl, O.; Lane, R.S.; Levin, M.L.; Tsao, J.I. Diapause in ticks of the medically important Ixodes ricinus species complex. Ticks Tick-Borne Dis. 2016, 7, 992–1003. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Pena, A.; Ayllon, N.; de la Fuente, J. Impact of climate trends on tick-borne pathogen transmission. Front. Physiol. 2012, 3, 64. [Google Scholar] [CrossRef] [PubMed]
- Heyman, P.; Cochez, C.; Hofhuis, A.; van der Giessen, J.; Sprong, H.; Porter, S.R.; Losson, B.; Saegerman, C.; Donoso-Mantke, O.; Niedrig, M.; et al. A clear and present danger: Tick-borne diseases in Europe. Expert Rev. Anti Infect. Ther. 2010, 8, 33–50. [Google Scholar] [CrossRef] [PubMed]
- Papa, A.; Chaligiannis, I.; Xanthopoulou, K.; Papaioakim, M.; Papanastasiou, S.; Sotiraki, S. Ticks parasitizing humans in Greece. Vector Borne Zoonotic Dis. 2011, 11, 539–542. [Google Scholar] [CrossRef]
- Papadopoulos, B.; Morel, P.C.; Aeschlimann, A. Ticks of domestic animals in the Macedonia region of Greece. Vet. Parasitol. 1996, 63, 25–40. [Google Scholar] [CrossRef] [PubMed]
- Chaligiannis, I.; Musella, V.; Rinaldi, L.; Cringoli, G.; de la Fuente, J.; Papa, A.; Sotiraki, S. Species diversity and spatial distribution of ixodid ticks on small ruminants in Greece. Parasitol. Res. 2016, 115, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Pavlidou, V.; Gerou, S.; Kahrimanidou, M.; Papa, A. Ticks infesting domestic animals in northern Greece. Exp. Appl. Acarol. 2008, 45, 195–198. [Google Scholar] [CrossRef] [PubMed]
- Papa, A.; Tsioka, K.; Daskou, M.A.; Minti, F.; Papadopoulou, E.; Melidou, A.; Giadinis, N. Application of 16S rRNA next generation sequencing in ticks in Greece. Heliyon 2020, 6, e04542. [Google Scholar] [CrossRef]
- Papa, A.; Tsioka, K.; Kontana, A.; Papadopoulos, C.; Giadinis, N. Bacterial pathogens and endosymbionts in ticks. Ticks Tick-Borne Dis. 2017, 8, 31–35. [Google Scholar] [CrossRef]
- Papadopoulos, O.; Paschaleri-Papadopoulou, E.; Deligaris, N.; Doukas, G. Isolation of tick borne encephalitis virus from a flock of goats with abortions and fatal disease (preliminary report). Vet News Greece 1971, 3, 112–114. [Google Scholar]
- Papa, A.; Pavlidou, V.; Antoniadis, A. Greek goat encephalitis virus strain isolated from Ixodes ricinus, Greece. Emerg. Infect. Dis. 2008, 14, 330–332. [Google Scholar] [CrossRef] [PubMed]
- ELSTAT. 2021. Available online: https://elstat-outsourcers.statistics.gr/Census2022_GR.pdf (accessed on 20 January 2024).
- Zianis, D.; Mencuccini, M. Aboveground biomass relationships for beech (Fagus moesiaca Cz.) trees in Vermio Mountain, Northern Greece, and generalised equations for Fagus sp. Ann. For. Sci. 2003, 60, 439–448. [Google Scholar] [CrossRef]
- Estrada-Peña, A.; Mihalca, A.D.; Petney, T.N. Ticks of Europe and North Africa; Springer Cham: New York, NY, USA, 2018. [Google Scholar] [CrossRef]
- Biggerstaff, B.J. PooledInfRate: A Microsoft® Office Excel© Add-In to Compute Prevalence Estimates from Pooled Samples, Version 4.0; Centers for Disease Control and Prevention: Fort Collins, CO, USA, 2009. [Google Scholar]
- Mysterud, A.; Stigum, V.M.; Jaarsma, R.I.; Sprong, H. Genospecies of Borrelia burgdorferi sensu lato detected in 16 mammal species and questing ticks from northern Europe. Sci. Rep. 2019, 9, 5088. [Google Scholar] [CrossRef] [PubMed]
- Regnery, R.L.; Spruill, C.L.; Plikaytis, B.D. Genotypic identification of rickettsiae and estimation of intraspecies sequence divergence for portions of two rickettsial genes. J. Bacteriol. 1991, 173, 1576–1589. [Google Scholar] [CrossRef] [PubMed]
- Scaramozzino, N.; Crance, J.M.; Jouan, A.; DeBriel, D.A.; Stoll, F.; Garin, D. Comparison of flavivirus universal primer pairs and development of a rapid, highly sensitive heminested reverse transcription-PCR assay for detection of flaviviruses targeted to a conserved region of the NS5 gene sequences. J. Clin. Microbiol. 2001, 39, 1922–1927. [Google Scholar] [CrossRef] [PubMed]
- Klimentov, A.S.; Butenko, A.M.; Khutoretskaya, N.V.; Shustova, E.Y.; Larichev, V.F.; Isaeva, O.V.; Karganova, G.G.; Lukashev, A.N.; Gmyl, A.P. Development of pan-phlebovirus RT-PCR assay. J. Virol. Methods 2016, 232, 29–32. [Google Scholar] [CrossRef] [PubMed]
- Honig, J.E.; Osborne, J.C.; Nichol, S.T. The high genetic variation of viruses of the genus Nairovirus reflects the diversity of their predominant tick hosts. Virology 2004, 318, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Hansford, K.M.; McGinley, L.; Wheeler, B.W.; Tschirren, B.; Medlock, J.M. Ixodes ricinus density, Borrelia prevalence and the density of infected nymphs along an urban-rural gradient in southern England. Zoonoses Public Health 2023, 70, 304–314. [Google Scholar] [CrossRef]
- Pettersson, J.H.; Shi, M.; Bohlin, J.; Eldholm, V.; Brynildsrud, O.B.; Paulsen, K.M.; Andreassen, A.; Holmes, E.C. Characterizing the virome of Ixodes ricinus ticks from northern Europe. Sci. Rep. 2017, 7, 10870. [Google Scholar] [CrossRef]
- Tomazatos, A.; von Possel, R.; Pekarek, N.; Holm, T.; Rieger, T.; Baum, H.; Bialonski, A.; Maranda, I.; Erdelyi-Molnar, I.; Spinu, M.; et al. Discovery and genetic characterization of a novel orthonairovirus in Ixodes ricinus ticks from Danube Delta. Infect. Genet. Evol. 2021, 88, 104704. [Google Scholar] [CrossRef] [PubMed]
- Vanmechelen, B.; Laenen, L.; Vergote, V.; Maes, P. Grotenhout Virus, a Novel Nairovirus Found in Ixodes ricinus in Belgium. Genome Announc. 2017, 5, 10-1128. [Google Scholar] [CrossRef] [PubMed]
- Tack, W.; Madder, M.; Baeten, L.; De Frenne, P.; Verheyen, K. The abundance of Ixodes ricinus ticks depends on tree species composition and shrub cover. Parasitology 2012, 139, 1273–1281. [Google Scholar] [CrossRef] [PubMed]
- Qviller, L.; Viljugrein, H.; Loe, L.E.; Meisingset, E.L.; Mysterud, A. The influence of red deer space use on the distribution of Ixodes ricinus ticks in the landscape. Parasites Vectors 2016, 9, 545. [Google Scholar] [CrossRef] [PubMed]
- Simser, J.A.; Palmer, A.T.; Fingerle, V.; Wilske, B.; Kurtti, T.J.; Munderloh, U.G. Rickettsia monacensis sp. nov., a spotted fever group Rickettsia, from ticks (Ixodes ricinus) collected in a European city park. Appl. Environ. Microbiol. 2002, 68, 4559–4566. [Google Scholar] [CrossRef] [PubMed]
- de Sousa, R.; dos Santos, M.L.; Cruz, C.; Almeida, V.; Garrote, A.R.; Ramirez, F.; Seixas, D.; Manata, M.J.; Maltez, F. Rare Case of Rickettsiosis Caused by Rickettsia monacensis, Portugal, 2021. Emerg. Infect. Dis. 2022, 28, 1068–1071. [Google Scholar] [CrossRef]
- Jado, I.; Oteo, J.A.; Aldámiz, M.; Gil, H.; Escudero, R.; Ibarra, V.; Portu, J.; Portillo, A.; Lezaun, M.J.; García-Amil, C.; et al. Rickettsia monacensis and human disease, Spain. Emerg. Infect. Dis. 2007, 13, 1405–1407. [Google Scholar] [CrossRef] [PubMed]
- Madeddu, G.; Mancini, F.; Caddeo, A.; Ciervo, A.; Babudieri, S.; Maida, I.; Fiori, M.L.; Rezza, G.; Mura, M.S. Rickettsia monacensis as cause of Mediterranean spotted fever-like illness, Italy. Emerg. Infect. Dis. 2012, 18, 702–704. [Google Scholar] [CrossRef] [PubMed]
- Parola, P.; Paddock, C.D.; Socolovschi, C.; Labruna, M.B.; Mediannikov, O.; Kernif, T.; Abdad, M.Y.; Stenos, J.; Bitam, I.; Fournier, P.E.; et al. Update on tick-borne rickettsioses around the world: A geographic approach. Clin. Microbiol. Rev. 2013, 26, 657–702. [Google Scholar] [CrossRef]
- Papa, A.; Dalla, V.; Petala, A.; Maltezou, H.C.; Maltezos, E. Fatal Mediterranean spotted fever in Greece. Clin. Microbiol. Infect. 2010, 16, 589–592. [Google Scholar] [CrossRef]
- Stanek, G.; Strle, F. Lyme borreliosis-from tick bite to diagnosis and treatment. FEMS Microbiol. Rev. 2018, 42, 233–258. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control. Factsheet on Lyme Borreliosis, for Healthcare Professionals; European Centre for Disease Prevention and Control: Stockholm, Sweden, 2015. [Google Scholar]
- Diaz, P.; Arnal, J.L.; Remesar, S.; Perez-Creo, A.; Venzal, J.M.; Vazquez-Lopez, M.E.; Prieto, A.; Fernandez, G.; Lopez, C.M.; Panadero, R.; et al. Molecular identification of Borrelia spirochetes in questing Ixodes ricinus from northwestern Spain. Parasites Vectors 2017, 10, 615. [Google Scholar] [CrossRef] [PubMed]
- Glass, A.; Springer, A.; Raulf, M.K.; Fingerle, V.; Strube, C. 15-year Borrelia prevalence and species distribution monitoring in Ixodes ricinus/inopinatus populations in the city of Hanover, Germany. Ticks Tick-Borne Dis. 2023, 14, 102074. [Google Scholar] [CrossRef]
- Kachrimanidou, M.; Papa, A.; Chochlakis, D.; Pavlidou, V.; Psaroulaki, A. Molecular evidence for Anaplasma phagocytophilum in Ixodes ricinus ticks from Greece. Vector-Borne Zoonotic Dis. 2011, 11, 1391–1393. [Google Scholar] [CrossRef]
- Tsiodras, S.; Spanakis, N.; Spanakos, G.; Pervanidou, D.; Georgakopoulou, T.; Campos, E.; Petra, T.; Kanellopoulos, P.; Georgiadis, G.; Antalis, E.; et al. Fatal human anaplasmosis associated with macrophage activation syndrome in Greece and the Public Health response. J. Infect. Public Health 2017, 10, 819–823. [Google Scholar] [CrossRef] [PubMed]
- Chochlakis, D.; Psaroulaki, A.; Kokkini, S.; Kostanatis, S.; Arkalati, E.; Karagrannaki, E.; Tsiatis, K.; Tselentis, Y.; Gikas, A. First evidence of Anaplasma infection in Crete, Greece. Report of six human cases. Clin. Microbiol. Infect. 2009, 15 (Suppl. S2), 8–9. [Google Scholar] [CrossRef] [PubMed]
- Daniel, S.A.; Manika, K.; Arvanitidou, M.; Diza, E.; Symeonidis, N.; Antoniadis, A. Serologic evidence of human granulocytic ehrlichiosis, Greece. Emerg. Infect. Dis. 2002, 8, 643–644. [Google Scholar] [CrossRef] [PubMed]
- Chochlakis, D.; Papaeustathiou, A.; Minadakis, G.; Psaroulaki, A.; Tselentis, Y. A serosurvey of Anaplasma phagocytophilum in blood donors in Crete, Greece. Eur. J. Clin. Microbiol. Infect. Dis. 2008, 27, 473–475. [Google Scholar] [CrossRef]
- Papa, A.; Kontana, A.; Tsioka, K.; Chaligiannis, I.; Sotiraki, S. Novel phleboviruses detected in ticks, Greece. Ticks Tick-Borne Dis. 2016, 7, 690–693. [Google Scholar] [CrossRef]
- Papa, A.; Kontana, A.; Tsioka, K.; Saratsis, A.; Sotiraki, S. Novel phlebovirus detected in Haemaphysalis parva ticks in a Greek island. Ticks Tick-Borne Dis. 2017, 8, 157–160. [Google Scholar] [CrossRef]
- Vanmechelen, B.; Merino, M.; Vergote, V.; Laenen, L.; Thijssen, M.; Martí-Carreras, J.; Claerebout, E.; Maes, P. Exploration of the Ixodes ricinus virosphere unveils an extensive virus diversity including novel coltiviruses and other reoviruses. Virus Evol. 2021, 7, veab066. [Google Scholar] [CrossRef] [PubMed]
- Ohlendorf, V.; Marklewitz, M.; Kopp, A.; Yordanov, S.; Drosten, C.; Junglen, S. Huge diversity of phleboviruses in ticks from Strandja Nature Park, Bulgaria. Ticks Tick-Borne Dis. 2019, 10, 697–703. [Google Scholar] [CrossRef] [PubMed]
- Bratuleanu, B.E.; Răileanu, C.; Bennouna, A.; Chretien, D.; Bigot, T.; Guardado-Calvo, P.; Savuta, G.; Moutailler, S.; Eloit, M.; Temmam, S. Diversity of viruses in Ixodes ricinus in Europe including novel and potential arboviruses. Trans. Emerg. Dis. 2023. [Google Scholar] [CrossRef]
- Kodama, F.; Yamaguchi, H.; Park, E.; Tatemoto, K.; Sashika, M.; Nakao, R.; Terauchi, Y.; Mizuma, K.; Orba, Y.; Kariwa, H.; et al. A novel nairovirus associated with acute febrile illness in Hokkaido, Japan. Nat. Commun. 2021, 12, 5539. [Google Scholar] [CrossRef] [PubMed]
- Sameroff, S.; Tokarz, R.; Vucelja, M.; Jain, K.; Oleynik, A.; Boljfetic, M.; Bjedov, L.; Yates, R.A.; Margaletic, J.; Oura, C.A.L.; et al. Virome of Ixodes ricinus, Dermacentor reticulatus, and Haemaphysalis concinna Ticks from Croatia. Viruses 2022, 14, 929. [Google Scholar] [CrossRef]
- Wang, Y.C.; Wei, Z.; Lv, X.; Han, S.; Wang, Z.; Fan, C.; Zhang, X.; Shao, J.; Zhao, Y.H.; Sui, L.; et al. A new nairo-like virus associated with human febrile illness in China. Emerg. Microbes Infect. 2021, 10, 1200–1208. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tick Species | Number (%) |
|---|---|
| Ixodes ricinus (female) | 157 (35.7) |
| Ixodes ricinus (male) | 190 (43.2) |
| Ixodes ricinus (nymph) | 49 (11.1) |
| Ixodes ricinus (larva) | 2 (0.5) |
| Dermacentor marginatus (female) | 5 (1.1) |
| Dermacentor marginatus (male) | 4 (0.9) |
| Haemaphysalis punctata (female) | 2 (0.5) |
| Haemaphysalis punctata (male) | 1 (0.2) |
| Haemaphysalis inermis (female) | 9 (2.0) |
| Haemaphysalis inermis (male) | 2 (0.5) |
| Haemaphysalis inermis (nymph) | 2 (0.5) |
| Haemaphysalis parva (female) | 2 (0.5) |
| Haemaphysalis parva (male) | 2 (0.5) |
| Ixodes frontalis (nymph) | 6 (1.4) |
| Ixodes gibbosus (female) | 2 (0.5) |
| Rhipicephalus sanguineus s.l. (male) | 1 (0.2) |
| Rhipicephalus turanicus s.l. (female) | 2 (0.5) |
| Rhipicephalus turanicus s.l. (male) | 2 (0.5) |
| Total | 440 (100) |
| Microorganism | Positive Pools (%) | MIR% (CI) | MLE% (CI) | |
|---|---|---|---|---|
| Bacteria | Rickettsia spp. | 51 (63.75) | 12.81 (9.53–16.10) | 18.45 (14.02–23.53) |
| Borrelia spp. | 36 (45.0) | 8.79 (6.01–11.58) | 10.92 (7.80–14.72) | |
| A. phagocytophilum | 2 (2.5) | 0.50 (0.00–1.20) | 0.51 (0.09–1.64) | |
| Viruses | Phleboviruses | 3 (3.75) | 0.75 (0.00–3.05) | 0.77 (0.20–2.05) |
| Nairoviruses | 7 (8.75) | 1.76 (0.47–3.05) | 1.82 (0.80–3.56) | |
| Microorganism | No. of Pools (%) |
|---|---|
| Rickettsia spp. | 22 (27.5) |
| Borrelia spp. | 8 (10.0) |
| Rickettsia spp. + Borrelia spp. | 21 (26.25) |
| Rickettsia spp. + A. phagocytophilum | 1 (1.25) |
| Borrelia spp. + Nairovirus | 1 (1.25) |
| Rickettsia spp. + Borrelia spp. + A. phagocytophilum | 1 (1.25) |
| Rickettsia spp. + Phlebovirus + Nairovirus | 1 (1.25) |
| Rickettsia spp. + Borrelia spp. + Phlebovirus + Nairovirus | 2 (2.5) |
| Rickettsia spp. + Borrelia spp. + Nairovirus | 3 (3.75) |
| Negative | 20 (25.0) |
| Total | 80 (100.0) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsioka, K.; Saratsis, A.; Pappa, S.; Papa, A. Pathogens Detected in Questing Ixodes ricinus Ticks in a Mountainous Area in Greece. Pathogens 2024, 13, 449. https://doi.org/10.3390/pathogens13060449
Tsioka K, Saratsis A, Pappa S, Papa A. Pathogens Detected in Questing Ixodes ricinus Ticks in a Mountainous Area in Greece. Pathogens. 2024; 13(6):449. https://doi.org/10.3390/pathogens13060449
Chicago/Turabian StyleTsioka, Katerina, Anastasios Saratsis, Styliani Pappa, and Anna Papa. 2024. "Pathogens Detected in Questing Ixodes ricinus Ticks in a Mountainous Area in Greece" Pathogens 13, no. 6: 449. https://doi.org/10.3390/pathogens13060449
APA StyleTsioka, K., Saratsis, A., Pappa, S., & Papa, A. (2024). Pathogens Detected in Questing Ixodes ricinus Ticks in a Mountainous Area in Greece. Pathogens, 13(6), 449. https://doi.org/10.3390/pathogens13060449