
Sex Pheromone Receptor Ste2 Orchestrates Chemotropic Growth towards Pine Root Extracts in the Pitch Canker Pathogen Fusarium circinatum

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Fungal Isolates, Culture Conditions and DNA Extraction
2.2. Identification of ste2 Gene
2.3. Gene Disruption and Complementation
2.4. Quantitative Chemotropism Plate Assay
3. Results
3.1. Identification of ste2 Gene
3.2. Gene Disruption and Complementation
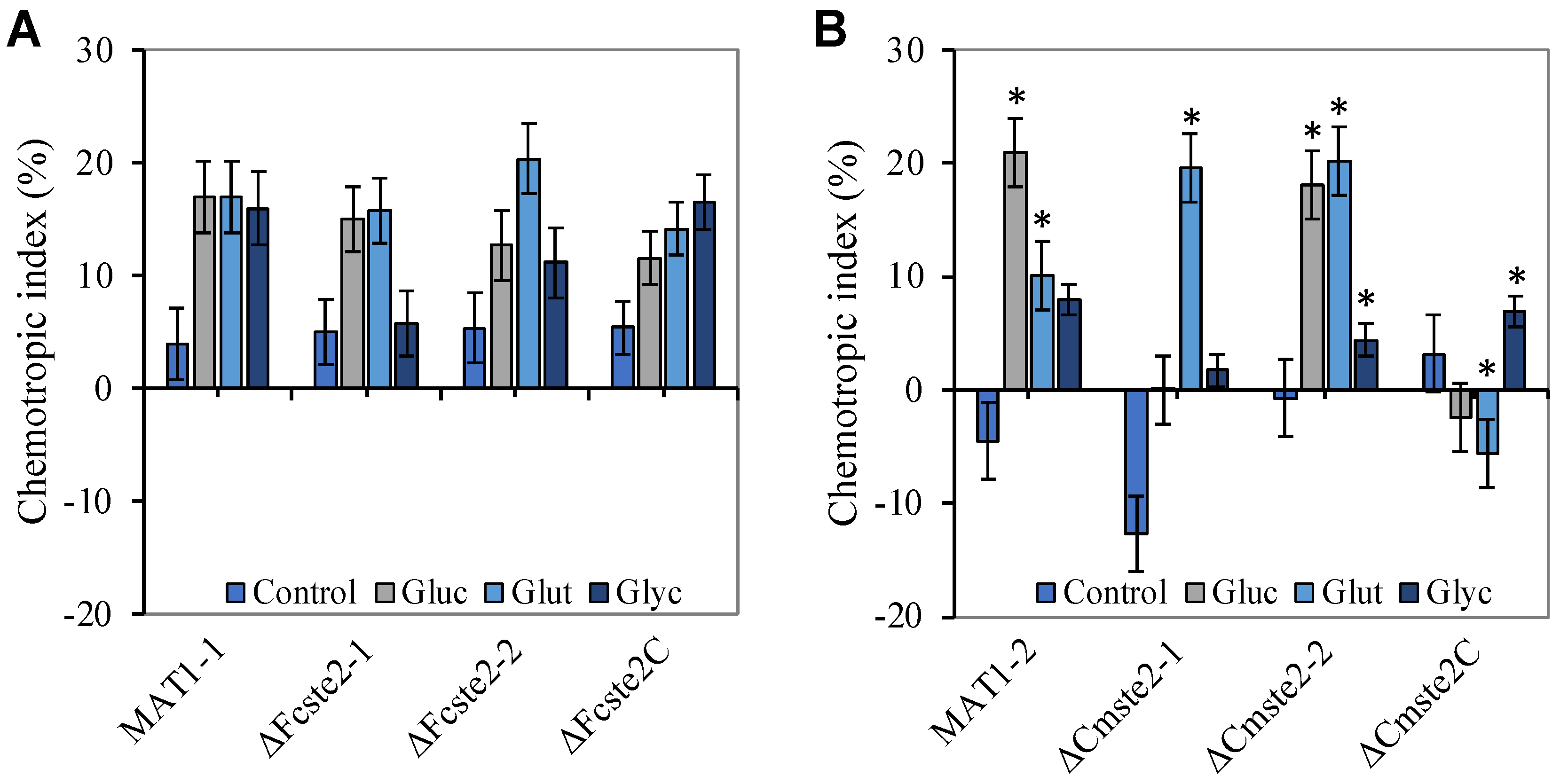
3.3. Deletion of ste2 Does Not Affect Chemotropism towards Glucose, Glycerol and Glutamate
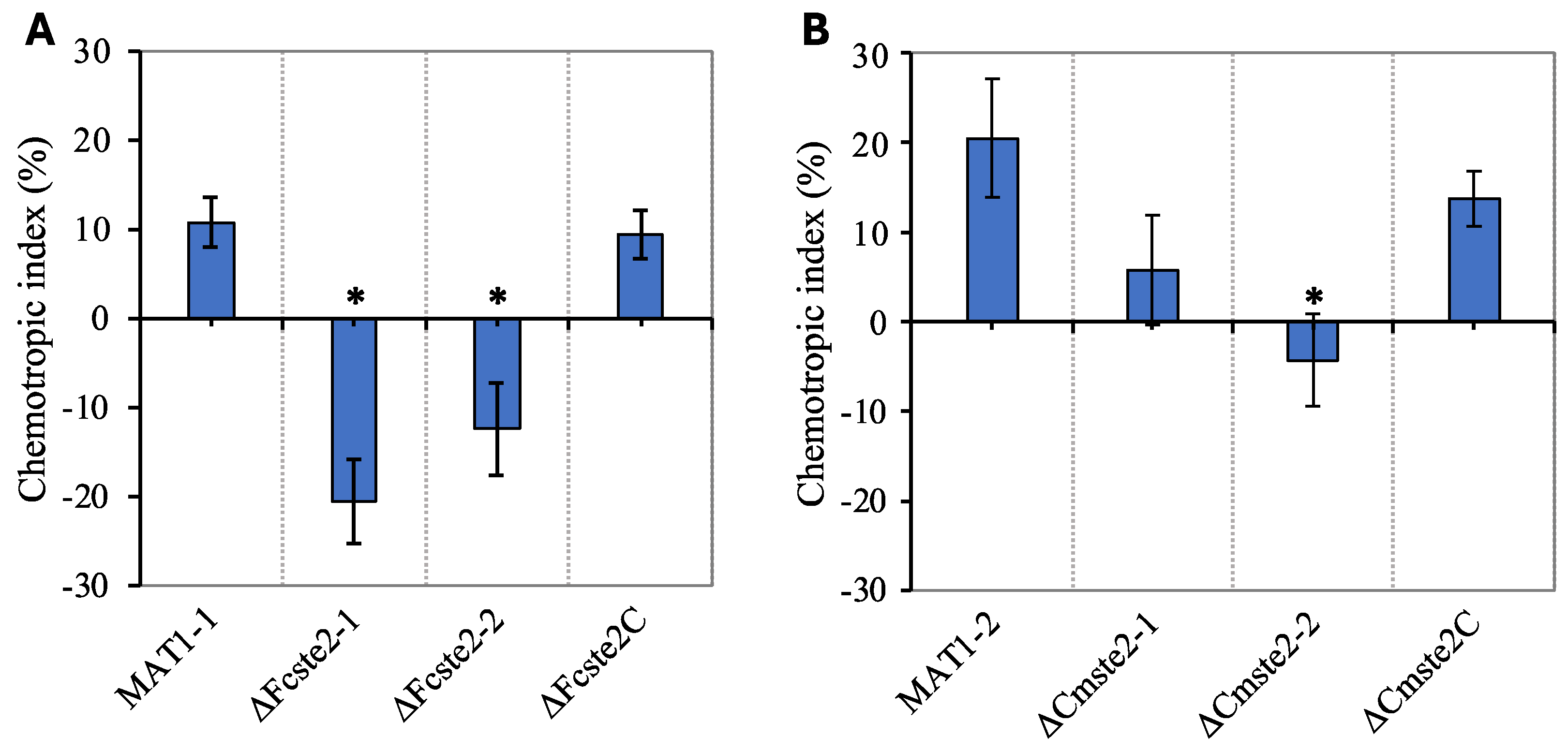
3.4. Ste2 Is Required for Chemotropism towards α-Factor Pheromone
3.5. Pine Root Exudate Provokes Ste2-Dependent Chemotropism
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Dijck, P.; Brown, N.A.; Goldman, G.H.; Rutherford, J.; Xue, C.; Van Zeebroeck, G. Nutrient sensing at the plasma membrane of fungal cells. Microbiol. Spectr. 2017, 5, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Oliveira, R.S.; Freitas, H.; Zhang, C. Biochemical and molecular mechanisms of plant-microbe-metal interactions: Relevance for phytoremediation. Front. Plant Sci. 2016, 7, 918. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Dong, F.; Liu, Q.; Lin, W.; Hu, C.; Yuan, Z. Soil metagenomics reveals effects of continuous sugarcane cropping on the structure and functional pathway of rhizospheric microbial community. Front. Microbiol. 2021, 12, 627569. [Google Scholar] [CrossRef] [PubMed]
- Bais, H.P.; Weir, T.L.; Perry, L.G.; Gilroy, S.; Vivanco, J.M. The role of root exudates in rhizosphere interactions with plants and other organisms. Annu. Rev. Plant Biol. 2006, 57, 233–266. [Google Scholar] [CrossRef] [PubMed]
- Haichar, F.E.Z.; Santaella, C.; Heulin, T.; Achouak, W. Root exudates mediated interactions belowground. Soil Biol. Biochem. 2014, 77, 69–80. [Google Scholar] [CrossRef]
- Huang, W.; Zwimpfer, E.; Hervé, M.R.; Bont, Z.; Erb, M. Neighbourhood effects determine plant–herbivore interactions below-ground. J. Ecol. 2018, 106, 347–356. [Google Scholar] [CrossRef]
- Beccaccioli, M.; Scala, V.; Reverberi, M. Communication with plants. In Encyclopedia of Mycology; Zaragoza, Ó., Casadevall, A., Eds.; Elsevier: Oxford, UK, 2021. [Google Scholar]
- Clark-Cotton, M.R.; Jacobs, K.C.; Lew, D.J. Chemotropism and cell-cell fusion in fungi. Microbiol. Mol. Biol. Rev. 2022, 86, e00165-21. [Google Scholar] [CrossRef] [PubMed]
- Turrà, D.; Di Pietro, A. Chemotropic sensing in fungus–plant interactions. Curr. Opin. Plant Biol. 2015, 26, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Vangalis, V.; Markakis, E.A.; Knop, M.; Di Pietro, A.; Typas, M.A.; Papaioannou, I.A. Components of TOR and MAP kinase signaling control chemotropism and pathogenicity in the fungal pathogen Verticillium dahliae. Microbiol. Res. 2023, 271, 127361. [Google Scholar] [CrossRef]
- Tsai, A.Y.-L.; Oota, M.; Sawa, S. Chemotactic host-finding strategies of plant endoparasites and endophytes. Front. Plant Sci. 2020, 11, 1167. [Google Scholar] [CrossRef]
- Turrà, D.; Nordzieke, D.; Vitale, S.; El Ghalid, M.; Di Pietro, A. Hyphal chemotropism in fungal pathogenicity. Semin. Cell Dev. Biol. 2016, 57, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Bary, A. Comparative Morphology and Biology of the Fungi, Mycetozoa and Bacteria; Clarendon Press: Liepzig, Germany, 1887. [Google Scholar]
- Nordzieke, D.E.; Fernandes, T.R.; El Ghalid, M.; Turrà, D.; Di Pietro, A. NADPH oxidase regulates chemotropic growth of the fungal pathogen Fusarium oxysporum towards the host plant. New Phytol. 2019, 224, 1600–1612. [Google Scholar] [CrossRef] [PubMed]
- Turrà, D.; Ghalid, M.E.; Rossi, F.; Di Pietro, A. Fungal pathogen uses sex pheromone receptor for chemotrophic sensing of host plant signals. Nature 2015, 527, 521–524. [Google Scholar] [CrossRef] [PubMed]
- El Ghalid, M. Identification of Compounds Secreted by Tomato Roots that Elicit Chemotropic Growth in Fusarium oxysporum. Ph.D. Thesis, University of Córdoba, Córdoba, Spain, 2015. [Google Scholar]
- Sridhar, P.S.; Trofimova, D.; Subramaniam, R.; González-Peña Fundora, D.; Foroud, N.A.; Allingham, J.S.; Loewen, M.C. Ste2 receptor-mediated chemotropism of Fusarium graminearum contributes to its pathogenicity against wheat. Sci. Rep. 2020, 10, 10770. [Google Scholar] [CrossRef] [PubMed]
- Dagdas, Y.F.; Bozkurt, T.O. Fungal sex receptors recalibrated to detect host plants. Cell Host Microbe 2015, 18, 637–638. [Google Scholar] [CrossRef] [PubMed]
- Wilson, A.M.; Wilken, P.M.; Wingfield, M.J.; Wingfield, B.D. Genetic networks that govern sexual reproduction in the Pezizomycotina. Microbiol. Mol. Biol. Rev. 2021, 85, e00020–e00021. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Wright, S.J.; Park, G.; Ouyang, S.; Krystofova, S.; Borkovich, K.A. Roles for receptors, pheromones, G proteins, and mating type genes during sexual reproduction in Neurospora crassa. Genetics 2012, 190, 1389–1404. [Google Scholar] [CrossRef] [PubMed]
- Dyer, P.S.; Inderbitzin, P.; Debuchy, R. 14 Mating-type structure, function, regulation and evolution in the Pezizomycotina. In Growth, Differentiation and Sexuality; Wendland, J., Ed.; Springer International Publishing: Cham, Switzerland, 2016; pp. 351–385. [Google Scholar]
- Gonçalves-Sá, J.; Murray, A. Asymmetry in sexual pheromones Is not required for ascomycete mating. Curr. Biol. 2011, 21, 1337–1346. [Google Scholar] [CrossRef] [PubMed]
- Vitale, S.; Di Pietro, A.; Turrà, D. Autocrine pheromone signalling regulates community behaviour in the fungal pathogen Fusarium oxysporum. Nat. Microbiol. 2019, 4, 1443–1449. [Google Scholar] [CrossRef] [PubMed]
- Hinterdobler, W.; Li, G.; Turrà, D.; Schalamun, M.; Kindel, S.; Sauer, U.; Beier, S.; Iglesias, A.R.; Compant, S.; Vitale, S.; et al. Integration of chemosensing and carbon catabolite repression impacts fungal enzyme regulation and plant associations. bioRxiv 2021. bioRxiv:2021.05. [Google Scholar] [CrossRef]
- Schrick, K.; Garvik, B.; Hartwell, L.H. Mating in Saccharomyces cerevisiae: The role of the pheromone signal transduction pathway in the chemotropic response to pheromone. Genetics 1997, 147, 19–32. [Google Scholar] [CrossRef]
- Irzykowska, L.; Bocianowski, J.; Waśkiewicz, A.; Weber, Z.; Karolewski, Z.; Goliński, P.; Kostecki, M.; Irzykowski, W. Genetic variation of Fusarium oxysporum isolates forming fumonisin B1 and moniliformin. J. Appl. Genet. 2012, 53, 237–247. [Google Scholar] [CrossRef]
- Turgeon, B.G. Application of mating type gene technology to problems in fungal biology. Annu. Rev. Phytopathol. 1998, 36, 115–137. [Google Scholar] [CrossRef] [PubMed]
- Wingfield, M.J.; Hammerbacher, A.; Ganley, R.J.; Steenkamp, E.T.; Gordon, T.R.; Wingfield, B.D.; Coutinho, T.A. Pitch canker caused by Fusarium circinatum—A growing threat to pine plantations and forests worldwide. Austral. Plant Pathol. 2008, 37, 319–334. [Google Scholar] [CrossRef]
- Martín-Rodrigues, N.; Sanchez-Zabala, J.; Salcedo, I.; Majada, J.; González-Murua, C.; Duñabeitia, M.K. New insights into radiata pine seedling root infection by Fusarium circinatum. Plant Pathol. 2015, 64, 1336–1348. [Google Scholar] [CrossRef]
- Martin, S.H.; Wingfield, B.D.; Wingfield, M.J.; Steenkamp, E.T. Structure and evolution of the Fusarium mating type locus: New insights from the Gibberella fujikuroi complex. Fungal Genet. Biol. 2011, 48, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Wallace, M.M.; Covert, S.F. Molecular mating type assay for Fusarium circinatum. Appl. Environ. Microbiol. 2000, 66, 5506–5508. [Google Scholar] [CrossRef]
- Pfenning, L.H.; Costa, S.S.; De Melo, M.P.; Costa, M.; Ventura, J.A.; Auer, C.G.; Dos Santos, Á.F. First report and characterization of Fusarium circinatum, the causal agent of pitch canker in Brazil. Trop. Plant Pathol. 2014, 39, 210–216. [Google Scholar] [CrossRef]
- Gordon, T.R.; Storer, A.J.; Okamoto, D. Population structure of the pitch canker pathogen, Fusarium subglutinans f. sp. pini, in California. Mycol. Res. 1996, 100, 850–854. [Google Scholar] [CrossRef]
- Desjardins, A.E.; Plattner, R.D.; Gordon, T.R. Gibberella fujikuroi mating population A and Fusarium subglutinans from teosinte species and maize from Mexico and Central America. Mycol. Res. 2000, 104, 865–872. [Google Scholar] [CrossRef]
- Phasha, M.M.; Wingfield, M.J.; Wingfield, B.D.; Coetzee, M.P.A.; Hallen-Adams, H.; Fru, F.; Swalarsk-Parry, B.S.; Yilmaz, N.; Duong, T.A.; Steenkamp, E.T. Ras2 is important for growth and pathogenicity in Fusarium circinatum. Fungal Genet. Biol. 2021, 150, 103541. [Google Scholar] [CrossRef]
- Hirokawa, T.; Boon-Chieng, S.; Mitaku, S. SOSUI: Classification and secondary structure prediction system for membrane proteins. Bioinformatics 1998, 14, 378–379. [Google Scholar] [CrossRef]
- Kim, H.-K.; Lee, T.; Yun, S.-H. A putative pheromone signaling pathway is dispensable for self-fertility in the homothallic ascomycete Gibberella zeae. Fungal Genet. Biol. 2008, 45, 1188–1196. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Leslie, J.F.; Bowden, R.L. Expression and function of sex pheromones and receptors in thehomothallic ascomycete Gibberella zeae. Eukaryot. Cell 2008, 7, 1211–1221. [Google Scholar] [CrossRef] [PubMed]
- Fairhead, C.; Llorente, B.; Denis, F.; Soler, M.; Dujon, B. New vectors for combinatorial deletions in yeast chromosomes and for gap-repair cloning using ‘split-marker’ recombination. Yeast 1996, 12, 1439–1457. [Google Scholar] [CrossRef]
- Chung, K.-R.; Lee, M.-H. Split-marker-mediated transformation and targeted gene disruption in filamentous fungi. In Genetic Transformation Systems in Fungi, Volume 2; van den Berg, M.A., Maruthachalam, K., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 175–180. [Google Scholar]
- Slinski, S.; Kirkpatrick, S.C.; Gordon, T.R. Inheritance of virulence in Fusarium circinatum, the cause of pitch canker in trees. Plant Pathol. 2016, 65, 1292–1296. [Google Scholar] [CrossRef]
- Proctor, R.H.; Busman, M.; Seo, J.-A.; Lee, Y.W.; Plattner, R.D. A fumonisin biosynthetic gene cluster in Fusarium oxysporum strain O-1890 and the genetic basis for B versus C fumonisin production. Fungal Genet. Biol. 2008, 45, 1016–1026. [Google Scholar] [CrossRef] [PubMed]
- Hallen-Adams, H.E.; Cavinder, B.L.; Trail, F. Fusarium graminearum from expression analysis to functional analysis. In Fungal Genomics: Methods and Protocols; Xu, J.-R., Bluhm, B., Eds.; Humana Press: New York, NY, USA; Dordrecht, The Netherlands; Heidelberg, Germany; London, UK, 2011; Volume 722, pp. 79–101. [Google Scholar]
- Martin, S.H.; Wingfield, B.D.; Wingfield, M.J.; Steenkamp, E.T. Causes and consequences of variability in peptide mating pheromones of ascomycete fungi. Mol. Biol. Evol. 2011, 28, 1987–2003. [Google Scholar] [CrossRef]
- Eisel, D.; Seth, O.; Grünewald-Janho, S.; Kruchen, B.; Rüger, B. DIG Application Manual for Filter Hybridization, 4th ed.; Roche Diagnostics GmbH: Mannheim, Germany, 2008; Volume 2. [Google Scholar]
- Green, M.R.; Sambrook, J. Molecular Cloning. A Laboratory Manual, 4th ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2012; Volume 448. [Google Scholar]
- Brown, N.A.; Schrevens, S.; van Dijck, P.; Goldman, G.H. Fungal G-protein-coupled receptors: Mediators of pathogenesis and targets for disease control. Nat. Microbiol. 2018, 3, 402–414. [Google Scholar] [CrossRef] [PubMed]
- Burkholder, A.C.; Hartwell, L.H. The yeast α-factor receptor: Structural properties deduced from the sequence of the STE2 gene. Nucleic Acids Res. 1985, 13, 8463–8475. [Google Scholar] [CrossRef] [PubMed]
- Passardi, F.; Penel, C.; Dunand, C. Performing the paradoxical: How plant peroxidases modify the cell wall. Trends Plant Sci. 2004, 9, 534–540. [Google Scholar] [CrossRef]
- Kidwai, M.; Ahmad, I.Z.; Chakrabarty, D. Class III peroxidase: An indispensable enzyme for biotic/abiotic stress tolerance and a potent candidate for crop improvement. Plant Cell Rep. 2020, 39, 1381–1393. [Google Scholar] [CrossRef] [PubMed]
- Vives-Peris, V.; de Ollas, C.; Gómez-Cadenas, A.; Pérez-Clemente, R.M. Root exudates: From plant to rhizosphere and beyond. Plant Cell Rep. 2020, 39, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Cao, S.; Wang, Z.; Xu, H.; Liang, J.; Liu, H.; Wang, G.; Ding, M.; Wang, Q.; Gong, C.; et al. An expanded subfamily of G-protein-coupled receptor genes in Fusarium graminearum required for wheat infection. Nat. Microbiol. 2019, 4, 1582–1591. [Google Scholar] [CrossRef]
- Kistler, H.C. Genetic diversity in the plant-pathogenic fungus Fusarium oxysporum. Phytopathology 1997, 87, 474–479. [Google Scholar] [CrossRef] [PubMed]
- Fradin, E.F.; Thomma, B.P. Physiology and molecular aspects of Verticillium wilt diseases caused by V. dahliae and V. albo-atrum. Mol. Plant Pathol. 2006, 7, 71–86. [Google Scholar] [CrossRef] [PubMed]
- Fayyaz, A.; Robinson, G.; Chang, P.L.; Bekele, D.; Yimer, S.; Carrasquilla-Garcia, N.; Negash, K.; Surendrarao, A.; von Wettberg, E.J.B.; Kemal, S.-A.; et al. Hiding in plain sight: Genome-wide recombination and a dynamic accessory genome drive diversity in Fusarium oxysporum f.sp. ciceris. Proc. Nat. Acad. Sci. USA 2023, 120, e2220570120. [Google Scholar] [CrossRef] [PubMed]
- de Jonge, R.; Bolton, M.D.; Kombrink, A.; van den Berg, G.C.M.; Yadeta, K.A.; Thomma, B.P.H.J. Extensive chromosomal reshuffling drives evolution of virulence in an asexual pathogen. Genome Res. 2013, 23, 1271–1282. [Google Scholar] [CrossRef] [PubMed]
- Usami, T.; Itoh, M.; Amemiya, Y. Asexual fungus Verticillium dahliae is potentially heterothallic. J. Gen. Plant Pathol. 2009, 75, 422–427. [Google Scholar] [CrossRef]
- Fokkens, L.; Rep, M. Population genomics reveals meiotic recombination in Fusarium oxysporum. Proc. Nat. Acad. Sci. USA 2023, 120, e2309677120. [Google Scholar] [CrossRef]
- Vangalis, V.; Knop, M.; Typas, M.A.; Papaioannou, I.A. Conidial fusion in the asexual fungus Verticillium dahliae. bioRxiv 2020. bioRxiv:2020-12. [Google Scholar] [CrossRef]
- Lee, S.C.; Ni, M.; Li, W.; Shertz, C.; Heitman, J. The evolution of sex: A perspective from the fungal kingdom. Microbiol. Mol. Biol. Rev. 2010, 74, 298–340. [Google Scholar] [CrossRef] [PubMed]
- Partida-Hanon, A.; Maestro-López, M.; Vitale, S.; Turrà, D.; Di Pietro, A.; Martínez-del-Pozo, Á.; Bruix, M. Structure of fungal α mating pheromone in membrane mimetics suggests a possible role for regulation at the water-membrane interface. Front. Microbiol. 2020, 11, 2020. [Google Scholar] [CrossRef] [PubMed]
- Vitale, S.; Partida-Hanon, A.; Serrano, S.; Martínez-del-Pozo, Á.; Di Pietro, A.; Turrà, D.; Bruix, M. Structure-activity relationship of a mating pheromone from the fungal pathogen Fusarium oxysporum. J. Biol. Chem. 2017, 292, 3591–3602. [Google Scholar] [CrossRef] [PubMed]
- Jones, N.B.; Ford, C.M.; Light, M.E.; Nadel, N.L.; Greyling, I.; Fourie, G.; Wingfield, M.J.; Morris, A.R. Effect on nursery and field performance of Pinus patula seedlings after inoculation with Fusarium circinatum. South. For. 2014, 76, 125–136. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Strain | Genotype 1 | Source 2 |
|---|---|---|
| FSP34 | MAT1-1 wild type | FABI |
| CMWF1218 | MAT1-2 wild type | FABI |
| ΔFcste2-1 | Δste2: FSP34: GFP | This study |
| ΔFcste2-2 | Δste2: FSP34: GFP | This study |
| ΔCmste2-1 | Δste2: CMWF1218: GFP | This study |
| ΔCmste2-2 | Δste2: CMWF1218: GFP | This study |
| ΔFcste2C | Δste2 + Ste2: FSP34: GFP | This study |
| ΔCmste2C | Δste2 + Ste2: CMWF1218: GFP | This study |
| CMWF1219 | MAT1-2 wild type | FABI |
| CMWF1217 | MAT1-1 wild type | FABI |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramaswe, J.B.; Steenkamp, E.T.; De Vos, L.; Fru, F.F.; Adegeye, O.O.; Wingfield, B.D. Sex Pheromone Receptor Ste2 Orchestrates Chemotropic Growth towards Pine Root Extracts in the Pitch Canker Pathogen Fusarium circinatum. Pathogens 2024, 13, 425. https://doi.org/10.3390/pathogens13050425
Ramaswe JB, Steenkamp ET, De Vos L, Fru FF, Adegeye OO, Wingfield BD. Sex Pheromone Receptor Ste2 Orchestrates Chemotropic Growth towards Pine Root Extracts in the Pitch Canker Pathogen Fusarium circinatum. Pathogens. 2024; 13(5):425. https://doi.org/10.3390/pathogens13050425
Chicago/Turabian StyleRamaswe, Jane B., Emma T. Steenkamp, Lieschen De Vos, Felix F. Fru, Omotayo O. Adegeye, and Brenda D. Wingfield. 2024. "Sex Pheromone Receptor Ste2 Orchestrates Chemotropic Growth towards Pine Root Extracts in the Pitch Canker Pathogen Fusarium circinatum" Pathogens 13, no. 5: 425. https://doi.org/10.3390/pathogens13050425
APA StyleRamaswe, J. B., Steenkamp, E. T., De Vos, L., Fru, F. F., Adegeye, O. O., & Wingfield, B. D. (2024). Sex Pheromone Receptor Ste2 Orchestrates Chemotropic Growth towards Pine Root Extracts in the Pitch Canker Pathogen Fusarium circinatum. Pathogens, 13(5), 425. https://doi.org/10.3390/pathogens13050425