
Toxicity Screening of Fungal Extracts and Metabolites, Xenobiotic Chemicals, and Indoor Dusts with In Vitro and Ex Vivo Bioassay Methods

 , , , , and
, , , , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
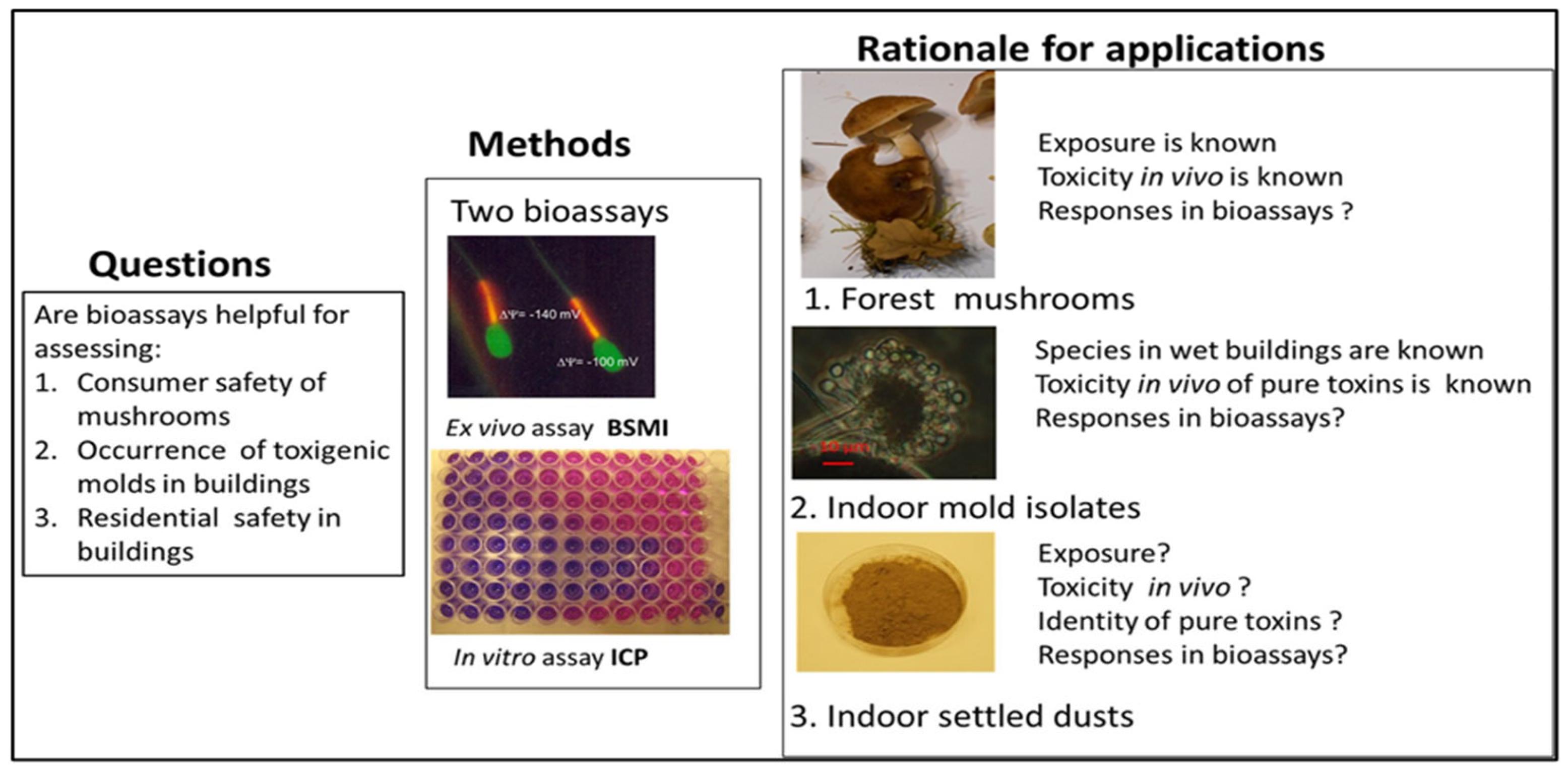
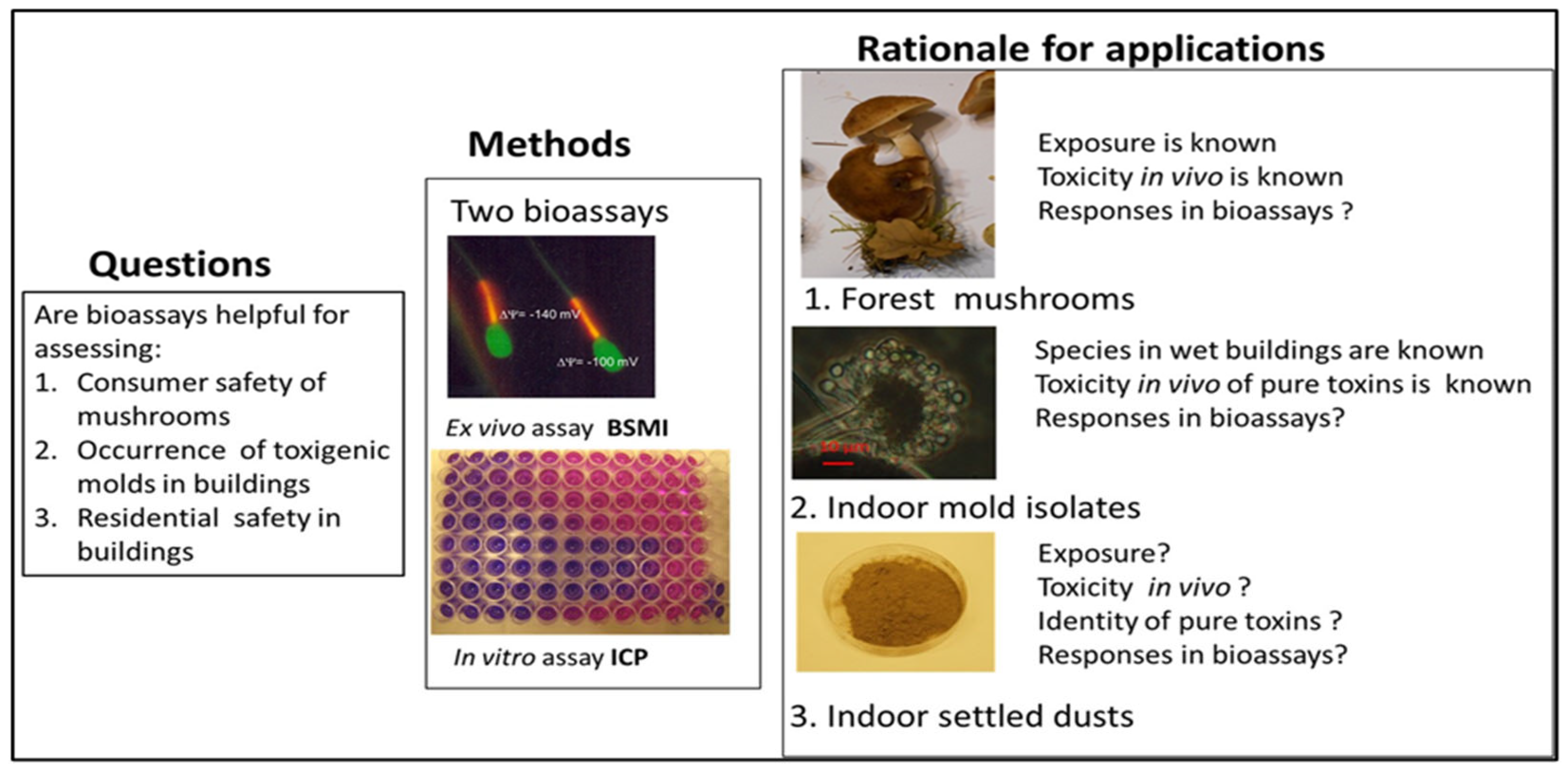
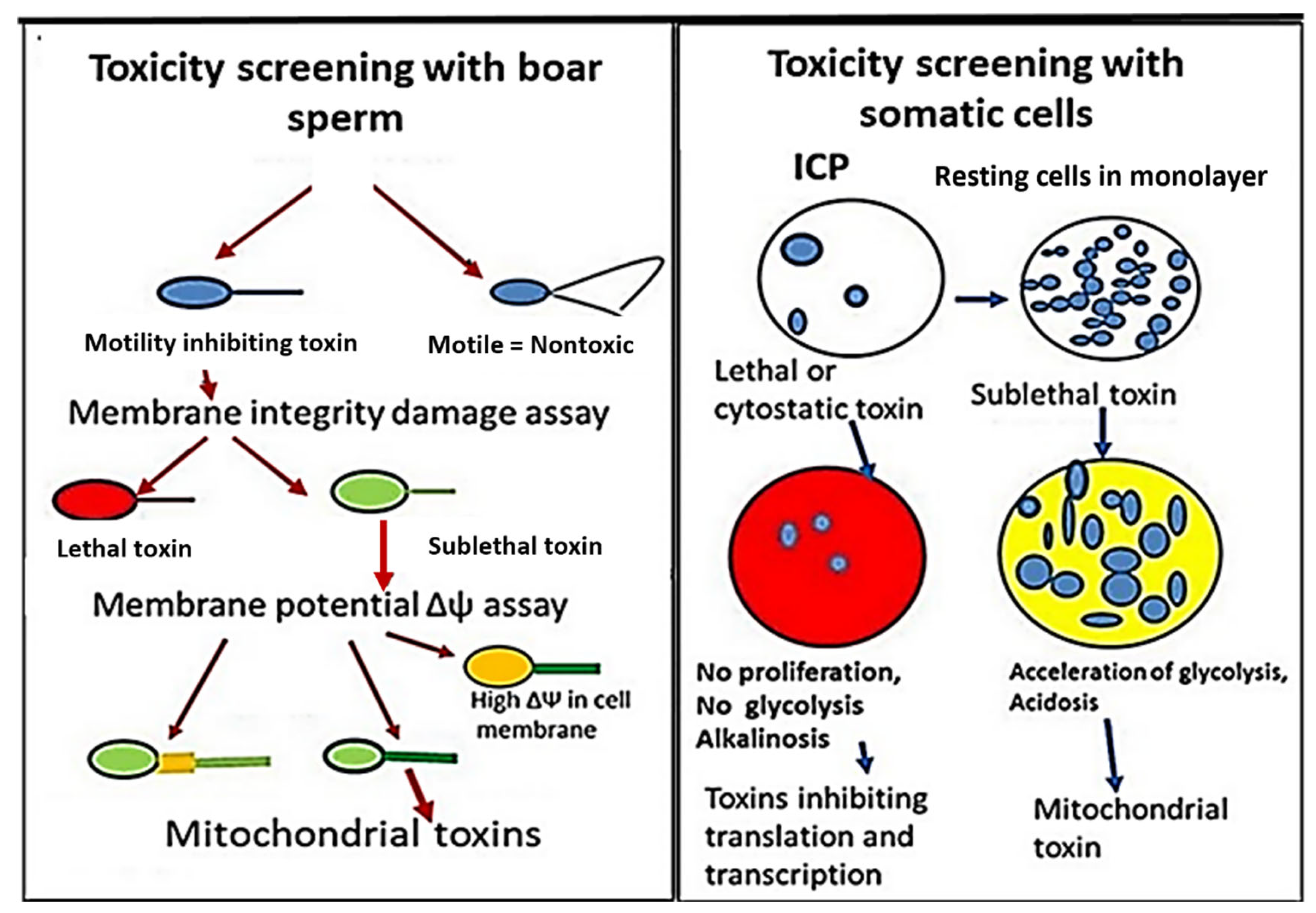
2.1.1. Rationale for Testing Forest Mushrooms, Indoor Molds, and Indoor Dusts with the Combination of Two Complementary Bioassays
2.1.2. Rationale for Applying Two Complementary Bioassays, Boar Spermatozoa Motility Inhibition Assay (BSMI), and Inhibition of Cell Proliferation Assay (ICP)
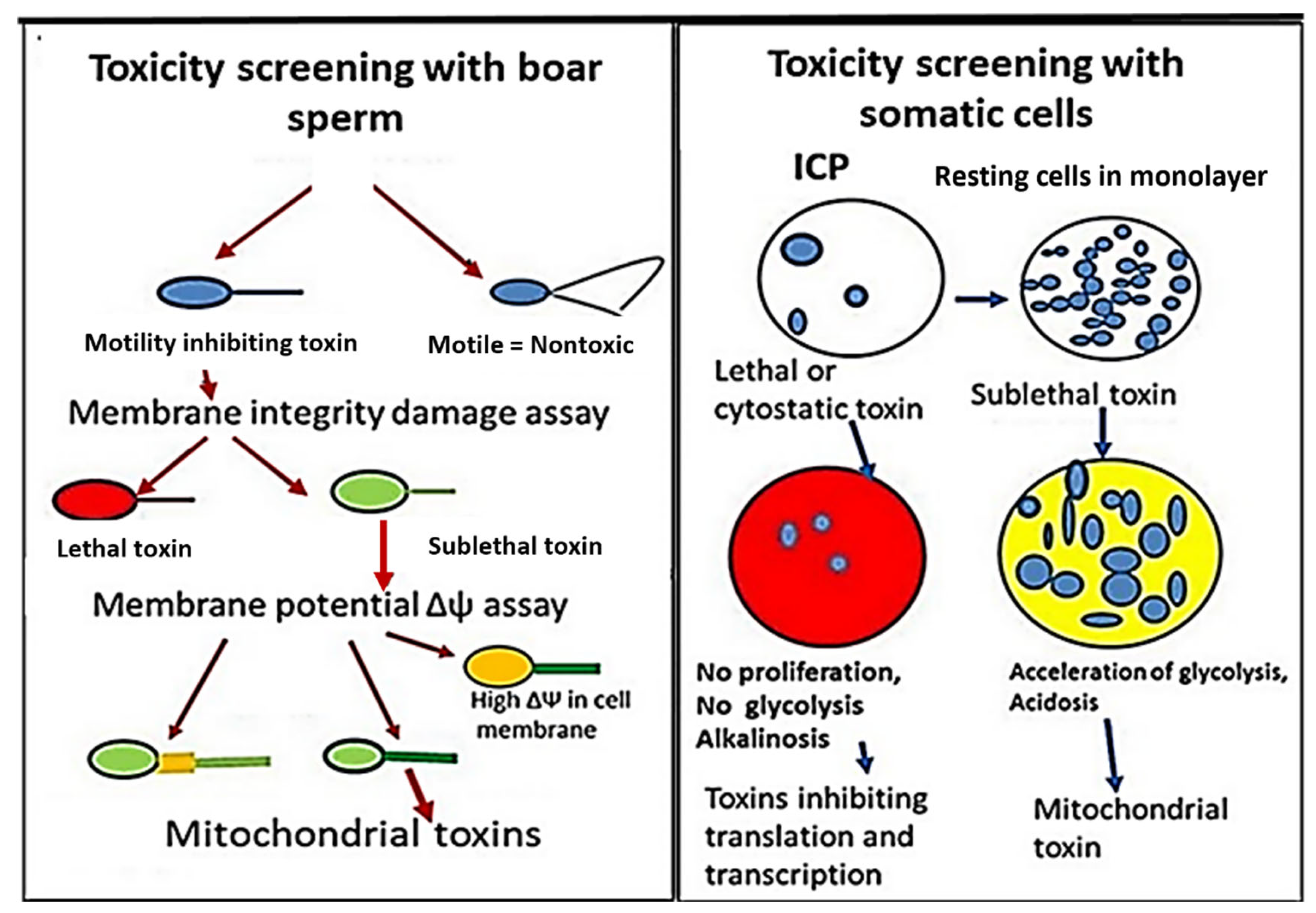
2.2. Estimation of Boar Sperm Motility Inhibition—The BSMI Assay
2.3. Plasma Membrane Integrity Depletion Assay and Membrane Potential Δψ Assay
2.4. Inhibition of Cell Proliferation Assay—ICP
2.5. Collection and Identification of Mushrooms
2.6. Isolation of Mold Colonies
2.7. The Toxicity Screening Assays of Molds and Mushrooms
2.8. Toxicity Testing of Pure Substances and Mycotoxins with the BSMI and ICP Assays
2.9. Toxicity Testing of Extracts of Settled Indoor Dusts
2.10. Identification of the Toxigenic Molds
2.11. Reagents and Supplies
3. Results
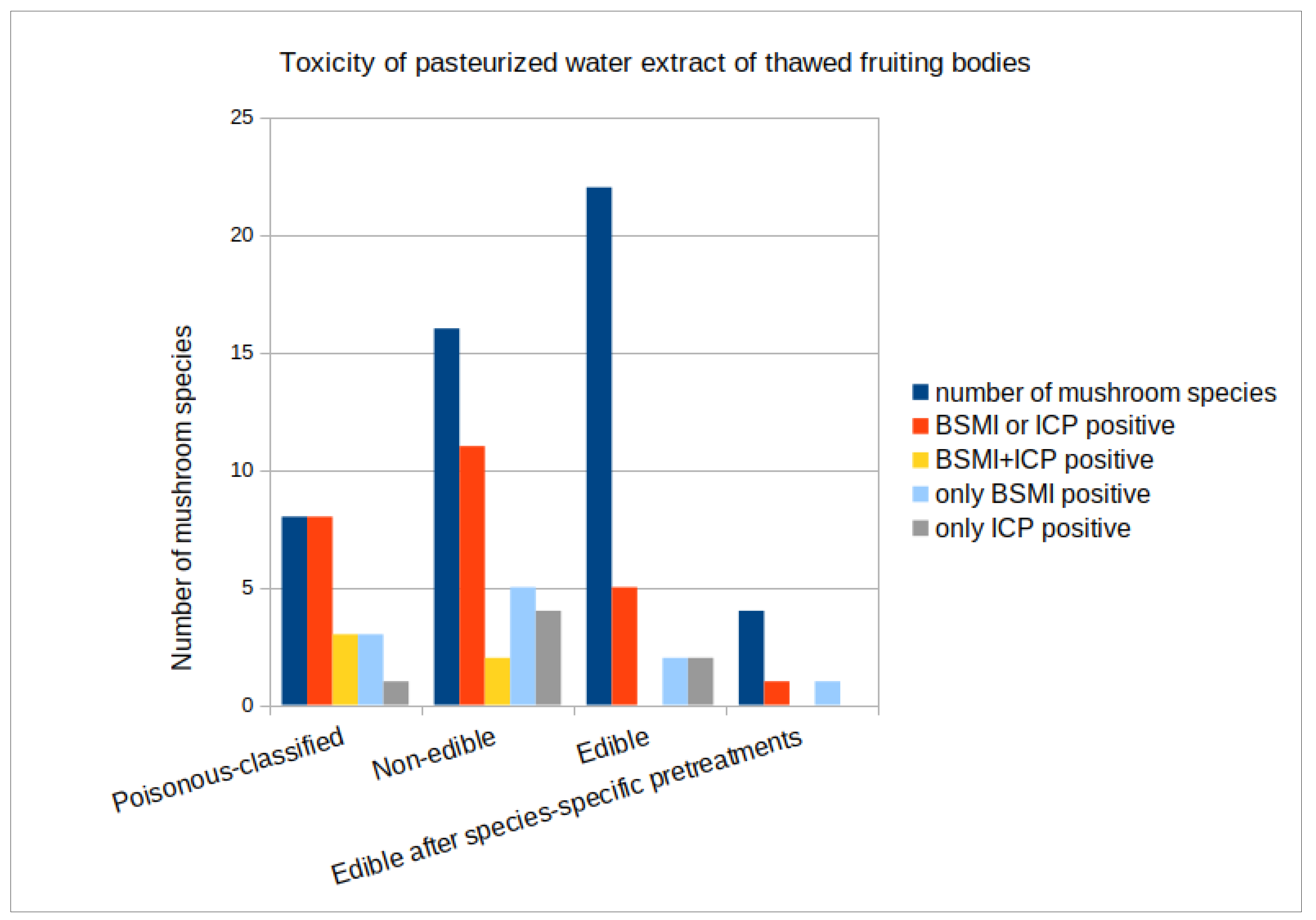
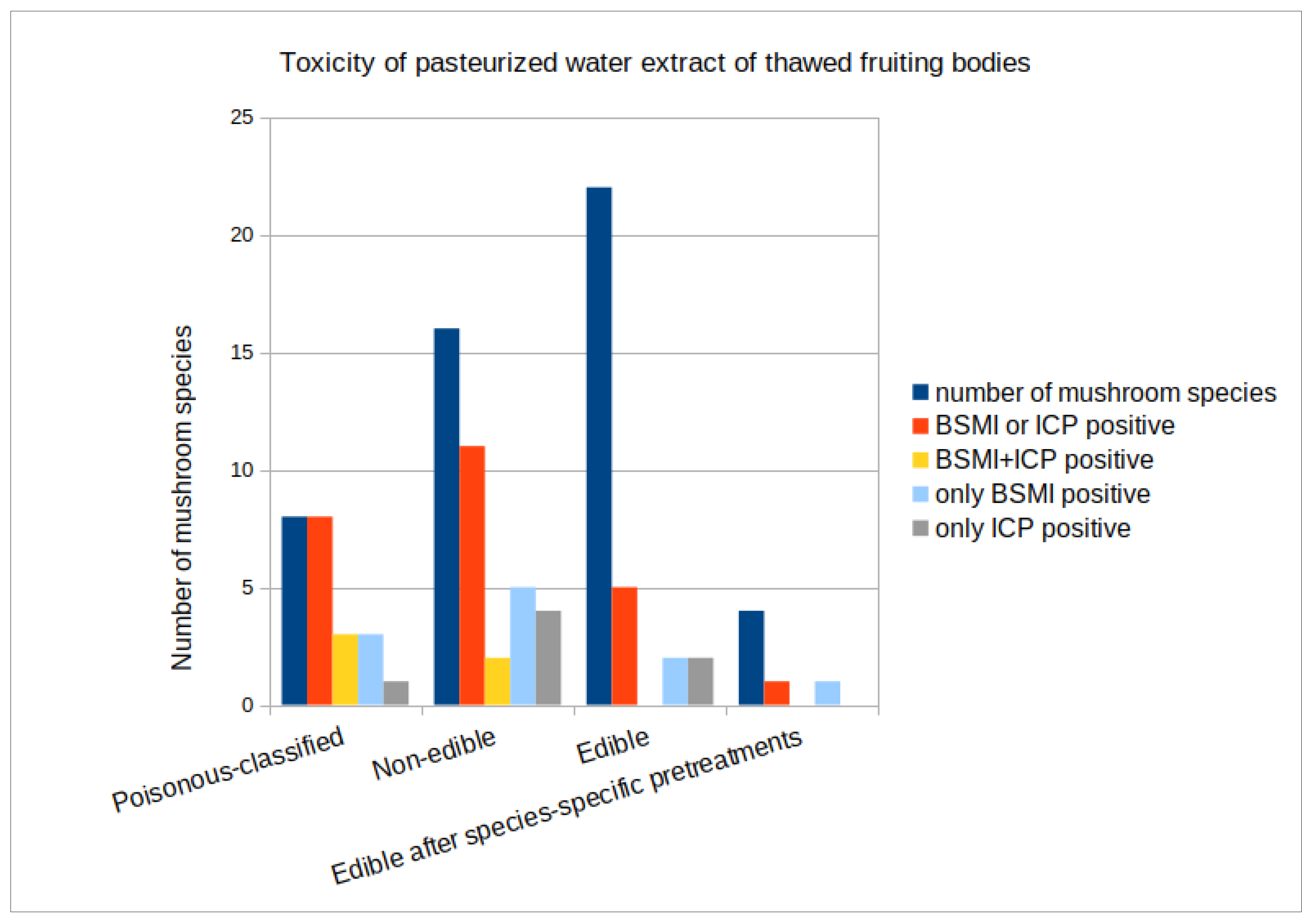
3.1. Toxicity of Forest Mushrooms
3.2. Toxicity of Indoor Mold Isolates
3.3. Toxicity of known Mycotoxins, Fungal Metabolites, and Xenobiotic Chemicals
3.4. Toxicity of Settled Indoor Dusts
3.5. Summary of Toxicity Responses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
| GenBank Accession | Species Name |
| PP165371 | Amanita fulva |
| PP152353 | Amanita virosa |
| PP152355 | Galerina marginata |
| PP152354 | Megacollybia platyphylla |
| PP165373 | Tricholoma sp. |
| PP165374 | Albatrellus ovinus |
| PP165376 | Tapinella atrotomentosa |
| PP165377 | Cortinarius sp. |
| PP165372 | Hygrophorus agathosmus |
| PP165375 | Clitocybe nebularis |
References
- De Schepper, J.; van Oorschot, Y.; Jaspers, R.J.; Hamers, T.; Lamoree, M.H.; Behnisch, P.; Besselink, H.; Houtman, C.J. The contribution of PFAS to thyroid hormone-displacing activity in Dutch waters: A comparison between two in vitro bioassays with chemical analysis. Environ. Int. 2023, 181, 108256. [Google Scholar] [CrossRef]
- Hamers, T.; Kortenkamp, A.; Scholze, M.; Molenaar, D.; Cenijn, P.H.; Weiss, J.M. Transthyretin-binding activity of complex mixtures representing the composition of thyroid hormone disrupting contaminants in house dust and human serum. Environ. Health Perspect. 2020, 128, 17015. [Google Scholar] [CrossRef]
- Collet, B.; Simon, E.; van der Linden, S.; el Abdellaoui, N.; Naderman, M.; Man, H.; Middelhof, I.; van der Burg, B.; Besselink, H.; Brouwer, A. Evaluation of a panel of in vitro methods for assessing thyroid receptor β and transthyretin transporter disrupting activities. Reprod. Toxicol. 2020, 96, 432–444. [Google Scholar] [CrossRef] [PubMed]
- Andersson, M.A.; Nikulin, M.; Köljalg, U.; Andersson, M.C.; Rainey, F.; Reijula, K.; Hintikka, E.L.; Salkinoja-Salonen, M. Bacteria, molds, and toxins in water-damaged building materials. Appl. Environ. Microbiol. 1997, 63, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Andersson, M.A.; Jääskeläinen, E.L.; Teplova, V.V.; Veijalainen, P.; Apetroaie, C.; Hoornstra, D.; Kroppenstedt, R.M.; Salkinoja-Salonen, M.S. Indoor bacilli and streptomycetes produce substances toxic to mammalian cells. In Bioaerosols, Fungi, Bacteria, Mycotoxins and Human Health: Patho-Physiology, Clinical Effects, Exposure Assessment, Prevention and Control in Indoor Environments and Work; Johanning, E., Ed.; Fungal Research Group Foundation: Albany, NY, USA, 2005; pp. 292–299. [Google Scholar]
- Andersson, M.A.; Mikkola, R.; Raulio, M.; Kredics, L.; Maijala, P.; Salkinoja-Salonen, M.S. Acrebol, a novel toxic peptaibol produced by an Acremonium exuviarum indoor isolate. J. Appl. Microbiol. 2009, 106, 909–923. [Google Scholar] [CrossRef] [PubMed]
- Mikkola, R.; Andersson, M.A.; Grigoriev, P.; Teplova, V.V.; Saris, N.E.; Rainey, F.A.; Salkinoja-Salonen, M.S. Bacillus amyloliquefaciens strains isolated from moisture-damaged buildings produced surfactin and a substance toxic to mammalian cells. Arch. Microbiol. 2004, 181, 314–323. [Google Scholar] [CrossRef] [PubMed]
- Vicente-Carrillo, A.; Edebert, I.; Garside, H.; Cotgreave, I.; Rigler, R.; Loitto, V.; Magnusson, K.E.; Rodríguez-Martínez, H. Boar spermatozoa successfully predict mitochondrial modes of toxicity: Implications for drug toxicity testing and the 3R principles. Toxicol. In Vitro 2015, 29, 582–591. [Google Scholar] [CrossRef]
- Ajao, C.; Andersson, M.A.; Teplova, V.V.; Nagy, S.; Gahmberg, C.G.; Andersson, L.C.; Hautaniemi, M.; Kakasi, B.; Roivainen, M.; Salkinoja-Salonen, M. Mitochondrial toxicity of triclosan on mammalian cells. Toxicol. Rep. 2015, 2, 624–637. [Google Scholar] [CrossRef]
- Castagnoli, E.; Salo, J.; Toivonen, M.S.; Marik, T.; Mikkola, R.; Kredics, L.; Vicente-Carrillo, A.; Nagy, S.; Andersson, M.T.; Andersson, M.A.; et al. An evaluation of boar spermatozoa as a biosensor for the detection of sublethal and lethal toxicity. Toxins 2018, 10, 463. [Google Scholar] [CrossRef]
- Andary, C.; Privat, G.; Bourrier, M.-J. Variations of monomethylhydrazine content in Gyromitra esculenta. Mycologia 1985, 77, 259–264. [Google Scholar] [CrossRef]
- Boden, K.; Drysdale, A.; Mogey, G. Constituents of Amanita muscaria. Nature 1965, 206, 1359–1360. [Google Scholar] [CrossRef] [PubMed]
- Patocka, J.; Wu, R.; Nepovimova, E.; Valis, M.; Wu, W.; Kuca, K. Chemistry and toxicology of major bioactive substances in Inocybe mushrooms. Int. J. Mol. Sci. 2021, 22, 2218. [Google Scholar] [CrossRef] [PubMed]
- Flammer, R.; Gallen, S. Hemolysis in mushroom poisoning: Facts and hypotheses. Schweiz. Med. Wochenschr. 1983, 113, 1555–1561. [Google Scholar] [PubMed]
- Shao, D.; Tang, S.; Healy, R.A.; Imerman, P.M.; Schrunk, D.E.; Rumbeiha, W.K. A novel orellanine containing mushroom Cortinarius armillatus. Toxicon 2016, 114, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Kemami Wangun, H.V.; Hertweck, C. Squarrosidine and pinillidine: 3,3′-fused bis(styrylpyrones) from Pholiota squarrosa and Phellinus pini. Eur. J. Org. Chem. 2007, 2007, 3292–3295. [Google Scholar] [CrossRef]
- Litten, W. The most poisonous mushrooms. Sci. Am. 1975, 232, 90–101. [Google Scholar] [CrossRef]
- List, P.H.; Luft, P. Gyromitrin, the poison of Gyromitra esculenta. 16. On the fungi contents. Arch. Pharmaz. 1968, 301, 294–305. [Google Scholar] [CrossRef]
- Magnusson, G.; Thoren, S.; Dahmen, J.; Leander, K. Fungal Extractives VIII. Two sesquiterpene furans from Lactarius. Acta Chem. Scand. 1974, 28, 841–846. [Google Scholar] [CrossRef]
- Salo, P.; Niemelä, T.; Salo, U. Suomen Sieniopas; Publisher WSOY: Tampere, Finland, 2006. [Google Scholar]
- Tyler, V.E.; Smith, A. Chromatographic detection of Amanita toxins in Galerina venenata. Mycologia 1963, 55, 358–359. [Google Scholar] [CrossRef]
- Nielsen, K.F. Mold Growth on Building Materials. Secondary Metabolites, Mycotoxins and Biomarkers. Ph.D. Thesis, Technical University of Denmark, Kongens Lyngby, Denmark, 2002. [Google Scholar]
- Andersen, B.; Frisvad, J.C.; Dunn, R.R.; Thrane, U. A pilot study on baseline fungi and moisture indicator fungi in Danish homes. J. Fungi 2021, 7, 71. [Google Scholar] [CrossRef]
- Andersson, M.; Varga, A.; Mikkola, R.; Vornanen-Winqvist, C.; Salo, J.; Kredics, L.; Kocsubé, S.; Salonen, H. Aspergillus was the dominant genus found during diversity tracking of cultivable potentially pathogenic indoor fungal isolates. Pathogens 2022, 11, 1171. [Google Scholar] [CrossRef] [PubMed]
- Nikulin, M.; Reijula, K.; Jarvis, B.B.; Hintikka, E.-L. Experimental lung mycotoxicosis in mice induced by Stachybotrys atra. Int. J. Exp. Pathol. 1996, 77, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Täubel, M.; Karvonen, A.M.; Reponen, T.; Hyvärinen, A.; Vesper, S.; Pekkanena, J. Application of the environmental relative moldiness index in Finland. Appl. Environ. Microbiol. 2016, 82, 578–584. [Google Scholar] [CrossRef] [PubMed]
- Vesper, S.; Barnes, C.; Ciaccio, C.E.; Johanns, A.; Kennedy, K.; Murphy, J.S.; Nunez-Alvarez, A.; Sandel, T.; Cox, D.; Dewalt, G.; et al. Higher Environmental Relative Moldiness Index (ERMI) values measured in homes of asthmatic children in Boston, Kansas City, and San Diego. J. Asthma 2013, 5, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Bloom, E.; Nyman, E.; Must, A.; Pehrson, C.; Larsson, L. Molds and mycotoxins in indoor environments—A survey in water-damaged buildings. J. Occup. Environ. Hygiene 2009, 6, 671–678. [Google Scholar] [CrossRef] [PubMed]
- Lindemann, V.; Schleiner, T.; Maier, U.; Fels, H.; Cramer, B.; Humpf, H. Analysis of mold and mycotoxins in naturally infested indoor building materials. Mycotoxin Res. 2022, 38, 205–220. [Google Scholar] [CrossRef]
- Hwang, H.M.; Park, E.; Young, T.; Hammock, B. Occurrence of endocrine-disrupting chemicals in indoor dust. Sci. Total Environ. 2008, 404, 26–35. [Google Scholar] [CrossRef]
- Fahimipour, A.; Ben Mamaar, S.; McFarland, A.G.; Blaustein, R.A.; Chen, J.; Glawe, A.J.; Kline, J.; Green, J.L.; Halden, R.U.; Van Den Wymelenberg, K.; et al. Antimicrobial chemicals associate with microbial function and antibiotic resistance indoors. mSystems 2018, 3, e00200-18. [Google Scholar] [CrossRef]
- Bonvallot, N.; Mandin, C.; Mercier, F.; Le Bot, B.; Glorennec, P. Health ranking of ingested semi-volatile organic compounds in house dust: An application to France. Indoor Air 2010, 20, 458–472. [Google Scholar] [CrossRef]
- Shi, T.; Wang, Y. Heavy metals in indoor dust: Spatial distribution, influencing factors and potential health risks. Sci. Total Environ. 2021, 755, 142367. [Google Scholar] [CrossRef]
- Li, T.; Yu, Y.; Sun, Z.; Duan, J. A comprehensive understanding of ambient particulate matter and its components on the adverse health effects based from epidemiological and laboratory evidence. Part Fibre Toxicol. 2022, 19, 67. [Google Scholar] [CrossRef]
- Pryor, J.; Hughes, C.; Foster, W.; Hales, B.; Robaire, B. Critical windows of exposure for children’s health: The reproductive system in animals and humans. Environ. Health Perspect. 2000, 108, 491–503. [Google Scholar] [CrossRef]
- Sousa, A.C.; Rato, L. The Impact of Endocrine-Disrupting Chemicals in Male Fertility. Encyclopedia. Available online: https://encyclopedia.pub/entry/19465 (accessed on 26 January 2024).
- Chen, X.; Mou, L.; Qu, J.; Wu, L.; Liu, C. Adverse effects of triclosan exposure on health and potential molecular mechanisms. Sci. Total Environ. 2023, 879, 163068. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Abdallah, M.F.; Chen, X.; Rajkovic, A. Current knowledge of individual and combined toxicities of aflatoxin B1 and fumonisin B1 in vitro. Toxins 2023, 15, 653. [Google Scholar] [CrossRef]
- Cresia, D.; Thurman, J.; Jones, L.; Nealley, M.; York, C.; Wannemacher, R.; Bunner, D. Acute inhalation toxicity of T-2 mycotoxin in mice. Fund. Appl. Toxicol. 1987, 8, 230–235. [Google Scholar] [CrossRef]
- Mantle, P.G.; Nolan, C.C. Pathological outcomes in kidney and brain in male Fischer rats given dietary ochratoxin A, commencing at one year of age. Toxins 2010, 2, 1100–1110. [Google Scholar] [CrossRef]
- Tephly, T. The toxicity of methanol. Life Sci. 1991, 48, 1031–1041. [Google Scholar] [CrossRef] [PubMed]
- Dierick, K.; Van Coillie, E.; Swiecicka, I.; Meyfroidt, G.; Devlieger, H.; Meulemans, A.; Hoedemaekers, G.; Fourie, L.; Heyndrickx, M.; Mahillon, J. Fatal family outbreak of Bacillus cereus-associated food poisoning. J. Clin. Microbiol. 2005, 43, 4277–4279. [Google Scholar] [CrossRef] [PubMed]
- Mahler, H.; Pasi, A.; Kramer, J.M.; Schulte, P.; Scoging, A.C.; Baer, W.; Kraehenbuehl, S. Fulminant liver failure in association with the emetictoxin of Bacillus cereus. N. Engl. J. Med. 1997, 336, 1143–1148. [Google Scholar] [CrossRef] [PubMed]
- Andersson, M.; Szabolcs, N.; Mikkola, R.; Kurnitski, J.; Salonen, H. Toxicities of detergents used in cleaning chemicals and hygiene products in a test battery of ex vivo and in vitro assays. In Proceedings of the Sisäilmasto seminaari, Report SYI 37. Messukeskus, Helsinki, Finland, 14 March 2019; pp. 297–305. Available online: https://www.researchgate.net/publication/359269938 (accessed on 16 March 2019).
- Castagnoli, E.; Marik, T.; Mikkola, R.; Kredics, L.; Andersson, M.A.; Salonen, H.; Kurnitski, J. Indoor Trichoderma strains emitting peptaibols in guttation droplets. J. Appl. Microbiol. 2018, 125, 1408–1422. [Google Scholar] [CrossRef] [PubMed]
- Bencsik, O.; Papp, T.; Berta, M.; Zana, A.; Forgó, P.; Dombi, G.; Andersson, M.; Salkinoja-Salonen, M.; Vágvölgyi, C.; Szekeres, A. Ophiobolin A from Bipolaris oryzae perturbs motility and membrane integrities of porcine sperm and induces cell death on mammalian somatic cell lines. Toxins 2014, 6, 2857–2871. [Google Scholar] [CrossRef] [PubMed]
- Salo, J.M.; Kedves, O.; Mikkola, R.; Kredics, L.; Andersson, M.A.; Kurnitski, J.; Salonen, H. Detection of Chaetomium globosum, Ch. cochliodes and Ch. rectangulare during the diversity tracking of mycotoxin-producing Chaetomium-like isolates obtained in buildings in Finland. Toxins 2020, 12, 443. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Shi, L.; Liu, W.; Hu, X.; Hui, Y.; Di, M.; Xue, S.; Zheng, Y.; Yao, M.; Li, C.; et al. Aloe-emodin induces mitochondrial dysfunction and pyroptosis by activation of the caspase-9/3/gasdermin E axis in HeLa cells. Front. Pharmacol. 2022, 13, 854526. [Google Scholar] [CrossRef] [PubMed]
- Tonshin, A.A.; Teplova, V.V.; Andersson, M.A.; Salkinoja-Salonen, M.S. The Fusarium mycotoxins enniatins and beauvericin cause mitochondrial dysfunction by acting the mitochondrial volume regulation, oxidative phosphorylation and ion homeostasis. Toxicology 2010, 276, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Thiel, P.G. A molecular mechanism for the toxic action of moniliformin, a mycotoxin produced by Fusarium moniliforme. Biochem. Pharmacol. 1978, 27, 483–486. [Google Scholar] [CrossRef]
- Mikkola, R.; Andersson, M.A.; Hautaniemi, M.; Salkinoja-Salonen, M.S. Toxic indole alkaloids avrainvillamide and stephacidin B produced by a biocide tolerant indoor mold Aspergillus westerdijkiae. Toxicon 2015, 99, 58–67. [Google Scholar] [CrossRef]
- Salo, M.J.; Marik, T.; Bencsik, O.; Mikkola, R.; Kredics, L.; Szekeres, A.; Andersson, M.A.; Salonen, H.; Kurnitski, J. Screening mold colonies by using two toxicity assays revealed indoor strains of Aspergillus calidoustus producing ophiobolins G and K. Toxins 2019, 11, 683. [Google Scholar] [CrossRef]
- Hoornstra, D.; Andersson, M.A.; Mikkola, R.; Salkinoja-Salonen, M. A new method for in vitro detection of microbially produced mitochondrial toxins. Toxicol. In Vitro 2003, 17, 745–751. [Google Scholar] [CrossRef]
- Mikkola, R.; Andersson, M.A.; Kredics, L.; Grigoriev, P.A.; Sundell, N.; Salkinoja-Salonen, M.S. 20-Residue and 11-residue peptaibols from the fungus Trichoderma longibrachiatum are synergistic in forming Na+/K+-permeable channels and adverse action towards mammalian cells. FEBS J. 2012, 279, 4172–4190. [Google Scholar] [CrossRef] [PubMed]
- Rasimus, S.; Mikkola, R.; Andersson, M.; Teplova, V.; Venediktova, N.; Ek-Kommonen, C.; Salkinoja-Salonen, M. Psychrotolerant Paenibacillus tundrae from barley grains produces new cereulide-like depsipeptides, paenilide, and homopaenilide, highly toxic to mammalian cells. Appl. Environ. Microbiol. 2012, 78, 3732–3743. [Google Scholar] [CrossRef] [PubMed]
- Hoornstra, D.; Andersson, M.A.; Teplova, V.V.; Mikkola, R.; Uotila, L.M.; Andersson, L.C.; Roivainen, M.; Gahmberg, C.G.; Salkinoja-Salonen, M.S. Potato crop as a source of emetic Bacillus cereus and cereulide-induced mammalian cell toxicity. Appl. Environ. Microbiol. 2013, 79, 3534–3543. [Google Scholar] [CrossRef] [PubMed]
- Salin, J.; Ohtonen, P.; Andersson, M.A.; Syrjälä, H. The toxicity of wiped dust and airborne microbes in individual classrooms increase the risk of teachers’ work-related symptoms: A cross-sectional study. Pathogens 2021, 10, 1360. [Google Scholar] [CrossRef] [PubMed]
- Mikkola, R.; Andersson, M.A.; Grigoriev, P.; Heinonen, M.; Salkinoja-Salonen, M.S. The toxic mode of action of cyclic lipodepsipeptide fusaricidins, produced by Paenibacillus polymyxa, toward mammalian cells. J. Appl. Microbiol. 2017, 123, 436–449. [Google Scholar] [CrossRef] [PubMed]
- Salo, M.J.; Marik, T.; Mikkola, R.; Andersson, M.A.; Kredics, L.; Salonen, H.; Kurnitski, J. Penicillium expansum strain isolated from indoor building material was able to grow on gypsum board and emitted guttation droplets containing chaetoglobosins and communesins A, B and D. J. Appl. Microbiol. 2019, 127, 1135–1147. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press, Inc.: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Andersson, M.A.; Salo, J.; Mikkola, R.; Marik, T.; Kredics, L.; Kurnitski, J.; Salonen, H. Melinacidin-producing Acrostalagmus luteoalbus, a major constituent of mixed mycobiota contaminating insulation material in an outdoor wall. Pathogens 2021, 10, 843. [Google Scholar] [CrossRef]
- Ruuska, R.M.; Hintikka, E.-L. Leccinum species causing poisoning. Sienilehti 1989, 41, 59–60. [Google Scholar]
- Hintikka, E.-L. Toxicity of mushroom samples in cell culture system. Karstenia 1978, 18, 40–42. [Google Scholar] [CrossRef]
- Barceloux, D.G. (Ed.) Amatoxin-Containing Mushrooms. In Medical Toxicology of Natural Substances: Foods, Fungi, Medicinal Herbs, Plants, and Venomous Animals, 1st ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2008; pp. 261–284. [Google Scholar] [CrossRef]
- Pyysalo, H. Tests for gyromitrin, a poisonous compound in false morel Gyromitra esculenta. Z. Lebensm. Unters. Forsch. 1976, 160, 325–330. [Google Scholar] [CrossRef]
- Antkowiak, W.Z.; Gessner, W.P. The structures of orellanine and orelline. Tetrahedron Lett. 1979, 20, 1931–1934. [Google Scholar] [CrossRef]
- Garner, C.D.; Armstrong, E.M.; Berry, R.E.; Beddoes, R.L.; Collison, D.; Cooney, J.J.A.; Ertok, S.N.; Helliwell, M. Investigations of amavadin. J. Inorg. Biochem. 2000, 80, 17–20. [Google Scholar] [CrossRef]
- Van Loon, P.C.C.; Swinkels, H.A.T.I.; Van Griensven, L.J.L.D. Dry matter content in mushrooms (Agaricus bisporus) as an indicator for mushroom quality. Mushroom Sci. 2000, 15, 507–513. [Google Scholar]
- Nerin, C.; Ubeda, J.L.; Alfaro, P.; Dahmani, Y.; Aznar, M.; Canellas, E.; Ausejo, R.; Nerin, C.; Ubeda, J.; Alfaro, P. Compounds from multilayer plastic bags cause reproductive failures in artificial insemination. Sci. Rep. 2015, 4, 4913. [Google Scholar] [CrossRef] [PubMed]
- Kadirvel, G.; Diengdoh, J.; Deori, S.; Dewry, R.K.; Abedin, S.N.; Moirangthem, P. Cytotoxic effects of heavy metals on functional attributes of boar sperm: An in vitro study. Front. Environ. Sci. 2023, 11, 1296606. [Google Scholar] [CrossRef]
- Huttunen, K.; Tirkkonen, J.; Taubel, M.; Krop, E.; Mikkonen, E.; Pekkanen, J.; Hirvonen, M.-R. Inflamatory potential in relation to the microbial content of settled dust samples collected from moisture damaged and reference schools: Results of HITEA study. Indoor Air 2016, 26, 380–390. [Google Scholar] [CrossRef] [PubMed]
- Läärä, E.; Pelkonen, O. Toxtest-Ulkopuolinen Tieteellinen Arvio. Ordered by the Ministry of Social Affair and Health. 2013. Available online: https://thl.fi/documents/98567/1586976/Toxtest+ulkopuolinen+arviointi+kesa+2013.pdf/dae8142b-74c4-4795-9223-53aee9c63521 (accessed on 11 May 2022).
- Salin, J.T.; Salkinoja-Salonen, M.; Salin, P.J.; Nelo, K.; Holma, T.; Ohtonen, P.; Syrjälä, H. Building-related symptoms are linked to the in vitro toxicity of indoor dust and airborne microbial propagules in schools: A cross-sectional study. Environ. Res. 2017, 154, 234–239. [Google Scholar] [CrossRef] [PubMed]
- Tuuminen, T.; Andersson, A.; Hyvönen, S.; Lohi, J.; Vaali, K. Indoor air nontoxicity should be proven with special techniques prior claiming that it may cause a variety of mental disorders. J. Hyg. Environ. Health 2020, 229, 113545. [Google Scholar] [CrossRef] [PubMed]
- Wargocki, P.; Sundell, J.; Bischof, W.; Brundrett, G.; Fanger, P.O.; Gyntelberg, F.; Hanssen, S.O.; Harrison, P.; Pickering, A.; Seppänen, O.; et al. Ventilation and health in non-industrial indoor environments: Report 566 from a European Multidisciplinary Scientific Consensus Meeting (EUROVEN). Indoor Air 2002, 12, 113–128. [Google Scholar] [CrossRef]
- Vornanen-Winqvist, C.; Salonen, H.; Järvi, K.; Andersson, M.A.; Mikkola, R.; Marik, T.; Kredics, L.; Kurnitski, J. Effects of ventilation improvement on measured and perceived indoor air quality in a school building with a hybrid ventilation system. Int. J. Environ. Res. Public Health 2018, 15, 1414. [Google Scholar] [CrossRef]
- Zhang, L.; Ou, C.; Magana-Arachchi, D.; Vithanage, M.; Vanka, K.S.; Palanisami, T.; Masakorala, K.; Wijesekara, H.; Yan, Y.; Bolan, N.; et al. Indoor particulate matter in urban households: Sources, pathways, characteristics, health effects, and exposure mitigation. Int. J. Environ. Res. Public Health 2021, 18, 11055. [Google Scholar] [CrossRef]
- Riad, M.A.; Abd-Rabo, M.M.; Abd El Aziz, S.A.; El Behairy, A.; Badawy, M. Reproductive toxic impact of subchronic treatment with combined butylparaben and triclosan in weanling male rats. J. Biochem. Mol. Toxicol. 2018, 32, e22037. [Google Scholar] [CrossRef] [PubMed]
- Melin, V.; Melin, T.; Dessify, B.; Nguyen, C.; Shea, C.; Hrubec, T. Quaternary ammonium disinfectants cause subfertility in mice by targeting both male and female reproductive processes. Reprod. Toxicol. 2016, 59, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Esplugas, R.; Rovira, J.; Mari, M.; Fernández-Arribas, J.; Eljarrat, E.; Domingo, J.L.; Schuhmacher, M. Emerging and legacy flame retardants in indoor air and dust samples of Tarragona Province (Catalonia, Spain). Sci. Total Environ. 2022, 806, 150494. [Google Scholar] [CrossRef] [PubMed]
- Weschler, C. Changes in indoor pollutants since the 1950s. Atmos. Environ. 2009, 43, 153–169. [Google Scholar] [CrossRef]
- Fan, X.; Kubwabo, C.; Rasmussen, P.; Jones-Otazo, H. Simultaneous quantitation of parabens, triclosan, and methyl triclosan in indoor house dust using solid phase extraction and gas chromatography-mass spectrometry. J. Environ. Monit. 2010, 12, 1891–1897. [Google Scholar] [CrossRef]
- Belova, L.; Poma, G.; Roggeman, M.; Jeong, Y.; Kim, D.H.; Berghmans, P.; Peters, J.; Salamova, A.; van Nuijs, A.; Covaci, A. Identification and characterization of quaternary ammonium compounds in Flemish indoor dust by ion-mobility high-resolution mass spectrometry. Environ. Int. 2023, 177, 108021. [Google Scholar] [CrossRef] [PubMed]
- Bornehag, C.G.; Lundgren, B.; Weschler, C.J.; Sigsgaard, T.; Hagerhed-Engman, L.; Sundell, J. Phthalates in indoor dust and their association with building characteristics. Environ. Health Perspect. 2005, 113, 1399–1404. [Google Scholar] [CrossRef]
- Acir, I.-H.; Guenther, K. Endocrine-disrupting metabolites of alkylphenol ethoxylates—A critical review of analytical methods, environmental occurrences, toxicity, and regulation. Sci. Total Environ. 2018, 635, 1530–1546. [Google Scholar] [CrossRef]
- Maqboolab, F.; Mostafalouc, S.; Bahadara, H.; Abdollahiad, M. Review of endocrine disorders associated with environmental toxicants and possible involved mechanisms. Life Sci. 2016, 145, 265–273. [Google Scholar] [CrossRef]
- Desalegn, A.A.; Iszatt, N.; Stigum, H.; Jensen, T.K.; Eggesbø, M. A case-cohort study of perinatal exposure to potential endocrine disrupters and the risk of cryptorchidism in the Norwegian HUMIS study. Environ. Int. 2021, 157, 106815. [Google Scholar] [CrossRef]
- Brulport, A.; Le Corre, L.; Maquart, G.; Barbet, V.; Dastugue, A.; Severin, I.; Vaiman, D.; Chagnon, M.-C. Multigenerational study of the obesogen effects of bisphenol S after a perinatal exposure in C57BL6/J mice fed a high fat diet. Environ. Poll. 2021, 270, 116243. [Google Scholar] [CrossRef]
- Meyer, J.; Hartman, J.; Mello, D. Mitochondrial toxicity. Toxicol. Sci. 2018, 162, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Roubicek, D.A.; de Souza-Pinto, N.C. Mitochondria and mitochondrial DNA as relevant targets for environmental contaminants. Toxicology 2017, 391, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Kochi, C.; Ahmad, S.; Salim, S. The effects of air pollution toxicants on the mitochondria. In Mitochondrial Intoxication; de Oliveira, M.R., Ed.; Academic Press: Cambridge, MA, USA, 2023; pp. 147–166. [Google Scholar] [CrossRef]
- Wang, M.; Tan, G.; Eljaszewicz, A.; Meng, Y.; Wawrzyniak, P.; Acharya, S.; Altunbulakli, C.; Westermann, P.; Dreher, A.; Yan, L.; et al. Laundry detergents and detergent residue after rinsing directly disrupt tight junction barrier integrity in human bronchial epithelial cells. J. Allergy Clin. Immunol. 2019, 143, 1892–1903. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Detergent | |||||
|---|---|---|---|---|---|
| Chemical Name | Type | Toxicity In Vivo LD50 | Reference | ||
| Fish mg L−1 | Rat p.o. mg kg−1 | ||||
| Genapol-X-080 † | Polyethyleneglycol monalkyl ether | nonionic | 5 | >2000 | [44] |
| Triton X-100 † | Polyethyleneglycol-p-is-octylphenyl ether | nonionic | 5 | 500 | [44] |
| TWEEN 80 † | Polyethylene glycol sorbitan mono oleate | nonionic | Not known | >25,000 | [44] |
| SDS † | Sodium dodecyl sulfate | anionic | 30 | 1288 | [44] |
| Biocides | |||||
| Chemical name | Biological activity | Toxicity in vivo LD50 | |||
| Fish mg L−1 | Rat p.o. mg kg−1 | ||||
| Triclosan † | Triclorooxy-phenylether | Mitochondrial toxin | 0.5 | 3700 | [9,44] |
| DDAC * | Didecyl- dimethyl- ammonium- chloride | Mitochondrial toxin | 0.5 | 238 | [44] |
| Fungal metabolites | |||||
| Chemical name | Biological activity | Toxicity in vivo LD50 Rat p.o. mg kg−1 | Ref. | ||
| Alamethicin † | Peptaibol | Cation channel-forming ionophore | 80 | [45] | |
| Citrinin † | Polyketide | Not known | 105 | ||
| Ophiobolin A ** | Sesterterpene | Not known | 238 | [46] | |
| Chaetoglobosin A † | Cytochalasin | Inhibitor of glucose transport | >400 | [47] | |
| Emodin † | Antraquinone | Mitochondrial toxin | 1000 | [48] | |
| Enniatin B † | Depsipeptide | K+ channel-forming inophore | 350 | [49] | |
| Moniliformin † | Dione | Mitochondrial toxin | 50 | [50] | |
| Ochratoxin A † | Isocoumarin | Not known | 20 | [51] | |
| Sterigmatocystin † | Polyketide | Inhibitor of translation | 120 | [52] | |
| T-toxin † | Trichothecene | Inhibitor of translation | 2.7 | [4] | |
| Roridin † | Trichothecene | Inhibitor of translation | 55 | [4] | |
| Verrucarin † | Trichothecene | Inhibitor of translation | 5.1 | [4] | |
| Valinomycin † | Depsipeptide | Mitochondrial toxin | 4 | [10] | |
| Toxic Response of Water Extract of Thawed Fruiting Bodies in Test Cell Solution % | Toxins According to the Literature | ||||||
|---|---|---|---|---|---|---|---|
| Ascomycetes | Basidiomycetes | In Vivo * | Ex Vivo BSMI ** | In Vitro ICP ** PK-15 | Known Toxins | Ref. | |
| Poisonous−Classified [19] Mushrooms (n = 8) | |||||||
| x | Amanita muscaria | + | (+) ≤1 | (−) 3 | Muscimol, ibotenic acid | [12] | |
| x | Amanita porphyria | + | (+) ≤1 | (−) 3 | |||
| x | Amanita virosa | + | (+) ≤1 | (+) 1 | Amatoxins | [17] | |
| x | Galerina marginata | + | (−) >1 | (+) 0.6 | Amatoxins | [21] | |
| x | Gyromitra esculenta | + | (+) ≤1 | n.a. | Gyromitrin | [18] | |
| x | Inocybe geophylla | + | (+) ≤1 | (+) 1 | Muscarine | [13] | |
| x | Lactarius helvus | + | (+) ≤1 | (+) 0.2 | Sesquiterpenes | [19] | |
| x | Paxillus involutus | + | (+) ≤1 | (−) 3 | [14] | ||
| Non−Edible [19] Mushrooms (n = 16) | |||||||
| x | Calocera viscosa | (+) | (−) >1 | (−) 5 | |||
| x | Clitocybe fragrans | n.a. | (+) ≤1 | (−) 5 | |||
| x | Clitocybe vibecinna | n.a. | (+) ≤1 | (−) 5 | |||
| x | Cortinarius armillatus | (+) | (+) ≤1 | (−) 3 | Orellanine | [15] | |
| x | Cortinarius collinitus | (+) | (−) >1 | (−) 5 | |||
| x | Cortinarius crassus | (+) | (−) >1 | (−) 5 | |||
| x | Cortinarius traganus | (+) | (−) >1 | (−) 5 | |||
| x | Hygrocybe conica | (+) | (+) ≤1 | (−) >5 | |||
| x | Hygrophorus erubescens | (+) | (+) ≤1 | (−) >5 | |||
| x | Leotia lubrica | (+) | (−) >1 | (+) 0.6 | [11] | ||
| x | Megacollybia platyphylla | (+) | (+) ≤1 | (+) 0.04 | |||
| x | Phaeolepiota aurea | (+) | (−) >1 | (+) 1 | |||
| x | Pholiota lenta | (+) | (−) >1 | (+) 1 | |||
| x | Pholiota squarrosa | (+) | (−) >1 | (+) 1 | Squarrosidine Pinillidine | [16] | |
| x | Tapinella atrotomentosa | (+) | (+) ≤1 | (+) 0.2 | |||
| x | Tricholoma equestre | (+) | (−) >1 | (−) >5 | |||
| Edible [19] Mushrooms (n = 18) | |||||||
| x | Albatrellus ovinus | − | (−) >1 | (+) 0.6 | |||
| x | Agaricus bisporus | − | (−) >1 | (−) 3 | |||
| x | Agaricus sylvicola | − | (−) >1 | (−) >5 | |||
| x | Amanita fulva | − | (−) >1 | (−) >5 | |||
| x | Ampulloclitocybe clavipes | − | (−) >1 | (−) 3 | |||
| x | Cantharellus cibarius | − | (−) >1 | (−) 3 | |||
| x | Cortinarius caperatus | − | (−) >1 | (−) >5 | |||
| x | Clitocybe odora | − | (+) ≤1 | (−) 3 | |||
| x | Craterellus cornucopioides | − | (−) >1 | (−) 3 | |||
| x | Craterellus tubaeformis | − | (−) >1 | (−) 5 | |||
| x | Hydnum rufescens | − | (−) >1 | (−) 3 | |||
| x | Kuehneromyces mutabilis | − | (−) >1 | (−) 3 | |||
| x | Leccinum scabrum | − | (−) >1 | (+) 1 | |||
| x | Lentinula edodes | − | (−) >1 | (−) >5 | |||
| x | Lycoperdon perlatum | − | (−) >1 | (−) 5 | |||
| x | Morchella elata | − | (−) >1 | n.a. | |||
| x | Phlegmacium triumphans | − | (−) >1 | (−) 3 | |||
| x | Suillus variegatus | − | (−) >1 | (−) >5 | |||
| Edible after Specific Pretreatments [19] (n = 4) *** | |||||||
| x | Lactarius rufus | − | (−) >1 | (−) >5 | |||
| x | Lactarius turpis | − | (−) >1 | (−) 5 | |||
| x | Lactarius vietus | − | (−) >1 | (−) >5 | |||
| x | Leccinum versipelle | − | (+) ≤1 | (−) 3 | |||
| Negative control | H2O | (−) > 2% | (−) > 5% | ||||
| Isolate Identifier | Toxicity after 2 d of Exposure | ||||||
|---|---|---|---|---|---|---|---|
| Species | Culture Collection Code | Lab Code | Identified Toxins | BSMI 2 | ICP 2 | Origin | Ref. |
| Acremonium exuviarum | DSM 21752 1 FBCC2543 2 | BMB4 | Acrebol | + | − | Russia | [6] |
| Acrostalagmus luteoalbus | SZMC 26545 3 | POB8 | Melinacidins | + | + | Finland | [62] |
| Aspergillus calidoustus | SZMC 22623 | MH4 | Ophiobolins | + | + | Finland | [52] |
| Aspergillus flavus | SZMC 24476 | 7D/SKK = 7D | Not identified 5 | − | + | Finland | [24] |
| Aspergillus fumigatus | SZMC 28215 | AE1 | Not identified 5 | − | + | Finland | [24] |
| Aspergillus niger | SZMC 27930 | Asp21 | Not identified 5 | − | + | Finland | [24] |
| Aspergillus versicolor | FBCC 2549 | SL/3 | Sterigmatocystin | − | + | Finland | [53] |
| Aspergillus westerdijkiae | FBCC 2553 | PP2 | Avrainvillamide Stephacidin B Ochratoxin | + | + | Finland | [24] [51] |
| Chaetomium globosum | SZMC 26534 | MTAV35 | Chaetoglobosin A+C | + | + | Finland | [47] |
| Chaetomium cochliodes | SZMC 24452 | OT7 | Chaetomin | − | + | Finland | [47] |
| Cladosporium sp. | n.a. | C11 | Not identified 5 | − | + | Finland | This study |
| Epicoccum sp. | FBCC 2565 | EMI | Not identified 5 | − | + | Finland | This study |
| Aspergillus pseudoglaucus | SZMC 27933 | 8/SL | Not identified 5 | + | + | Finland | This study |
| Penicillium chrysogenum | SZMC 22627 | RUK2/3 | Meleagrin | − | + | Finland | [59] |
| Penicillium expansum | SZMC 26543 FBCC2596 CBS 145620 4 | RcP61 = P61 | Communesin Chaetoglobosins | + | + | Finland | [59] |
| Penicillium glabrum | n.a. | PG21 | Not identified 5 | − | + | Finland | This study |
| Paecilomyces variotii | FBCC 2550 | Paec2 | Viriditoxin | + | − | Denmark | [24] |
| Paecilomyces sp. | FBCC2628 | ST32 | Not identified 5 | − | + | Finland | [24] |
| Stachybotrys chartarum | n.a. | RT | Satratoxin G | − | + | Finland | [4] |
| Trichoderma atroviride | SZMC 12750 | H1/226 | Trichorzianines | + | + | Finland | [45] |
| T. longibrachiatum | SZMC Thg | Thg | Trilongins | + | + | Finland | [45,54] |
| Negative control (% v/v) | Methanol | −(2%) | −(5%) | ||||
| − | − | Finland | |||||
| EC50 μg mL–1 | |||||
|---|---|---|---|---|---|
| In Vivo 3 | Ex Vivo BSMI 4 | In Vitro ICP 4 | Mitochondrial Toxicity 5 | ||
| 1–3d | 2d | 1d | Ref. | ||
| Mitochondrial Toxins (LD50 rat p.o. <1000 mg kg –1 Classified as Toxic in vivo +.) | |||||
| Acrebol 1 | ? | (+) 0.1 | (−) >10 | (+) 0.1 | [6] |
| Enniatin B 2 | + | (+) 0.5 | (−) 15 | (+) 2 | [49] |
| Moniliformin 2 | + | (+) 2 | (−) >20 | (+) 2 | [50] |
| Emodin 2 | ? | (+) 4 | (−) 45 | (+) 3 | [48] |
| Toxins Inhibiting the Glucose Transport | |||||
| Chaetoglobosin A 2 | + | (+) <1 | (+) 3 | ND | [47] |
| Alamethicin 2 | + | (+) 0.5 | (+) 6 | ND | [45] |
| Trilongins 1 | ? | (+) 0.4 | (+) 5 | ND | [45] |
| Trichorzianines 1 | ? | (+) 0.5 | (+) 5 | ND | [45] |
| Toxins with Undefined Function | |||||
| Ophiobolin | + | (+) 0.3 | (+) 0.1 | [46] | |
| Avrainvillamide 1 | ? | (+) 0.2 | (+) 0.2 | ND | [51] |
| Stephacidin B 1 | ? | (+) 0.3 | (++) 0.3 | ND | [51] |
| Citrinin 2 | + | (−) 20 | (+) 10 | [51] | |
| Toxins Inhibiting Translation | |||||
| Sterigmatocystin 2 | + | (−)>20 | (+) 0.1 | ND | [52] |
| T –2 toxin 2 | + | (−) 0.5 | (+) 0.0023 | [4] | |
| Satratoxin G 1 | ? | (−) 5 | (+) 0.0009 | [4] | |
| Verrucarin A 2 | + | (−) 5 | (+) 0.0006 | [4] | |
| Roridin A 2 | + | (−) >1 | (+) 0.0007 | [4] | |
| Mycotoxins Used as Positive Controls Calculated as the Mean of 10 Measurements | |||||
| Positive control for ICP assays (PK –15 and FFL cells) | |||||
| Citrinin | + | ≥20 | 13 (±1.6) | This study | |
| Positive control for the BSMI assay | |||||
| Enniatin B | + | 0.6 (±0.14) | 18 (±7.7) | This study | |
| Toxins without Response in the Bioassays Calculated from 20 Measurements | |||||
| Ochratoxin A 2 | + | (−) >50 | (−) >50 | – | [51] |
| Xenobiotic Chemicals, LD50 Fish Inhaled <10 μg mL –1 = Toxic in vivo Marked as +. | |||||
| DDAC | + | (+) 1 | (+) 0.5 | (+) 1 | [44] |
| Genapol X 080 | + | (+) 3 | (−) 30 | (+) 5 | [44] |
| Tween 80 | − | (+) 10 | (−) 500 | [44] | |
| Triton X –100 | + | (+) 1 | (−) 30 | [44] | |
| SDS | + | (+) 10 | (−) 120 | [44] | |
| Triclosan | + | (+) 1 | 20 (−) | 5 | [9] |
| Positive control for BSMI and ICP assay (PK –15 and FFL cells), calculated as the mean of 11 measurements | |||||
| Triclosan | + | 1 (±0.6) | 13 (±4.2) | This study | |
| Negative control % (v/v) | |||||
| Methanol | (−) 1% | (−) 10% | This study | ||
| Ethanol | (−) 1% | (−) 10% | This study | ||
| Toxicity | ||||
|---|---|---|---|---|
| Reported Health Complaints In Vivo | Ex Vivo BSMI | In Vitro ICP | Mitochondrial Toxicity 1 | |
| Wet Buildings Containing Visible Mold Growth (n = 12) | ||||
| A | + | (−) >50 | (−) >100 | − |
| B | + | (−) >50 | (−) >100 | − |
| C | + | (−) >50 | (−) >100 | − |
| D | + | (+) <25 | (+) <50 | (+) |
| E | + | (−) >50 | (−) >100 | − |
| F | + | (+) <25 | (−) >100 | (+) |
| G | + | (−) >50 | (−) >100 | |
| H | + | (−) >50 | (−) >100 | − |
| I | + | (+) <25 | (+) <50 | (+) |
| J | + | (+) <25 | (−) >100 | |
| K | + | (−) >50 | (−) >100 | |
| L | + | (+) <25 | (−) >100 | |
| Dry, mold−free buildings (n = 14) | ||||
| 1 | − | (+) <25 | (+) <50 | (+) |
| 2 | − | (+) <25 | (+) <50 | (+) |
| 3 | − | (+) <25 | (−) >100 | (+) |
| 4 | − | (+) <25 | (+) <50 | (+) |
| 5 | − | (+) <25 | (−) >100 | (+) |
| 6 | − | (−) >50 | (−) >100 | (−) |
| 7 | − | (−) >50 | (−) >100 | (−) |
| 8 | − | (+) <25 | (+) <50 | (+) |
| 9 | − | (+) <25 | (−) >100 | (+) |
| 10 | − | (+) <25 | (−) >100 | (−) |
| 11 | − | (−) >50 | (−) >100 | (−) |
| 12 | − | (−) >50 | (−) >100 | (−) |
| 13 | − | (−) >50 | (−) >100 | (−) |
| 14 | − | (−) >50 | (−) >100 | (−) |
| Positive controls | ||||
| Dusts from spoiled feed and bedding 3 (n = 6) | + | (+) <25 | (+) <50 | (+) |
| Hay dust spiked with valinomycin 1000 μg g−1 | (+) <0.1 | >5 | (+) | |
| Negative controls | ||||
| Hay dust (good quality hay) (n = 10) 2 | − | (−) >50 | (−) >100 | (−) |
| Methanol % (v/v) | + | (−) <1% | (−) <10% | (+) |
| Proportion of Positive Responses in Total Positive Test Responses Obtained in the Two Bioassays (%) | |||||
|---|---|---|---|---|---|
| Total ICP- Positive | Total BSMI-Positive | ICP- + BSMI- Positive | Only ICP- Positive | Only BSMI-Positive | |
| Mushrooms (n = 46) | 26 | 35 | 11 | 15 | 22 |
| Indoor molds (n = 21) | 90 | 48 | 38 | 52 | 10 |
| Fungal metabolites (n = 18) | 72 | 67 | 44 | 28 | 22 |
| Xenobiotic chemicals (n = 6) | 33 | 100 | 33 | none | 83 |
| Settled dusts (n = 23) | 23 | 50 | 26 | none | 27 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hintikka, T.; Andersson, M.A.; Lundell, T.; Marik, T.; Kredics, L.; Mikkola, R.; Andersson, M.C.; Kurnitski, J.; Salonen, H. Toxicity Screening of Fungal Extracts and Metabolites, Xenobiotic Chemicals, and Indoor Dusts with In Vitro and Ex Vivo Bioassay Methods. Pathogens 2024, 13, 217. https://doi.org/10.3390/pathogens13030217
Hintikka T, Andersson MA, Lundell T, Marik T, Kredics L, Mikkola R, Andersson MC, Kurnitski J, Salonen H. Toxicity Screening of Fungal Extracts and Metabolites, Xenobiotic Chemicals, and Indoor Dusts with In Vitro and Ex Vivo Bioassay Methods. Pathogens. 2024; 13(3):217. https://doi.org/10.3390/pathogens13030217
Chicago/Turabian StyleHintikka, Tuomas, Maria A. Andersson, Taina Lundell, Tamás Marik, László Kredics, Raimo Mikkola, Magnus C. Andersson, Jarek Kurnitski, and Heidi Salonen. 2024. "Toxicity Screening of Fungal Extracts and Metabolites, Xenobiotic Chemicals, and Indoor Dusts with In Vitro and Ex Vivo Bioassay Methods" Pathogens 13, no. 3: 217. https://doi.org/10.3390/pathogens13030217
APA StyleHintikka, T., Andersson, M. A., Lundell, T., Marik, T., Kredics, L., Mikkola, R., Andersson, M. C., Kurnitski, J., & Salonen, H. (2024). Toxicity Screening of Fungal Extracts and Metabolites, Xenobiotic Chemicals, and Indoor Dusts with In Vitro and Ex Vivo Bioassay Methods. Pathogens, 13(3), 217. https://doi.org/10.3390/pathogens13030217