

Molecular Identification of Piroplasmids in Ticks from Infested Small Ruminants in Konya Province, Turkey

 ,
,  ,
,  ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Study Area
2.3. Tick Collection and Identification
2.4. DNA Extraction
2.5. Molecular Identification of Tick-Borne Piroplasmids
2.6. Cloning and Sequencing Analysis
2.7. Phylogenetic and Statistical Analysis
2.8. Accession Numbers

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Piroplasmid | Target Gene | Primer Sequence (5′ to 3′) | Size (bp) | Annealing Temperature (°C) | References |
|---|---|---|---|---|---|
| Babesia/Theileria spp. | 18S rRNA | GTCTTGTAATTGGAATGATGG TAGTTTATGGTTAGGACTACG | ~500 | 55 | [39] |
| B. naoakii | AMA-1 | TGGCGCCGACTTCCTGGAGCCCATCTCCAA AGCTGGGGCCCTCCTTCGATGAACCGTCGG | 371 | 64 | [41] |
| B. ovis | 18S rRNA | TGGGCAGGACCTTGGTTCTTCT CCGCGTAGCGCCGGCTAAATA | 549 | 62 | [10] |
| T. lestoquardi | Merozoite surface antigen (Tlms) | GTGCCGCAAGTGAGTCA GGACTGATGAGAAGACGATGAG | 730 | 54 | [42] |
| T. luwenshuni | 18S rRNA | GGTAGGGTATTGGCCTACTGA TCATCCGGATAATACAAG | 389 | 57 | [43] |
| T. ovis | 18S rRNA | TCGAGACCTTCGGGT TCCGGACATTGTAAAACAAA | 520 | 53 | [44] |
| T. uilenbergi | 18S rRNA | GGTAGGGTATTGGCCTACCGG ACACTCGGAAAATGCAAGCA | 388 | 55 | [43] |
3. Results
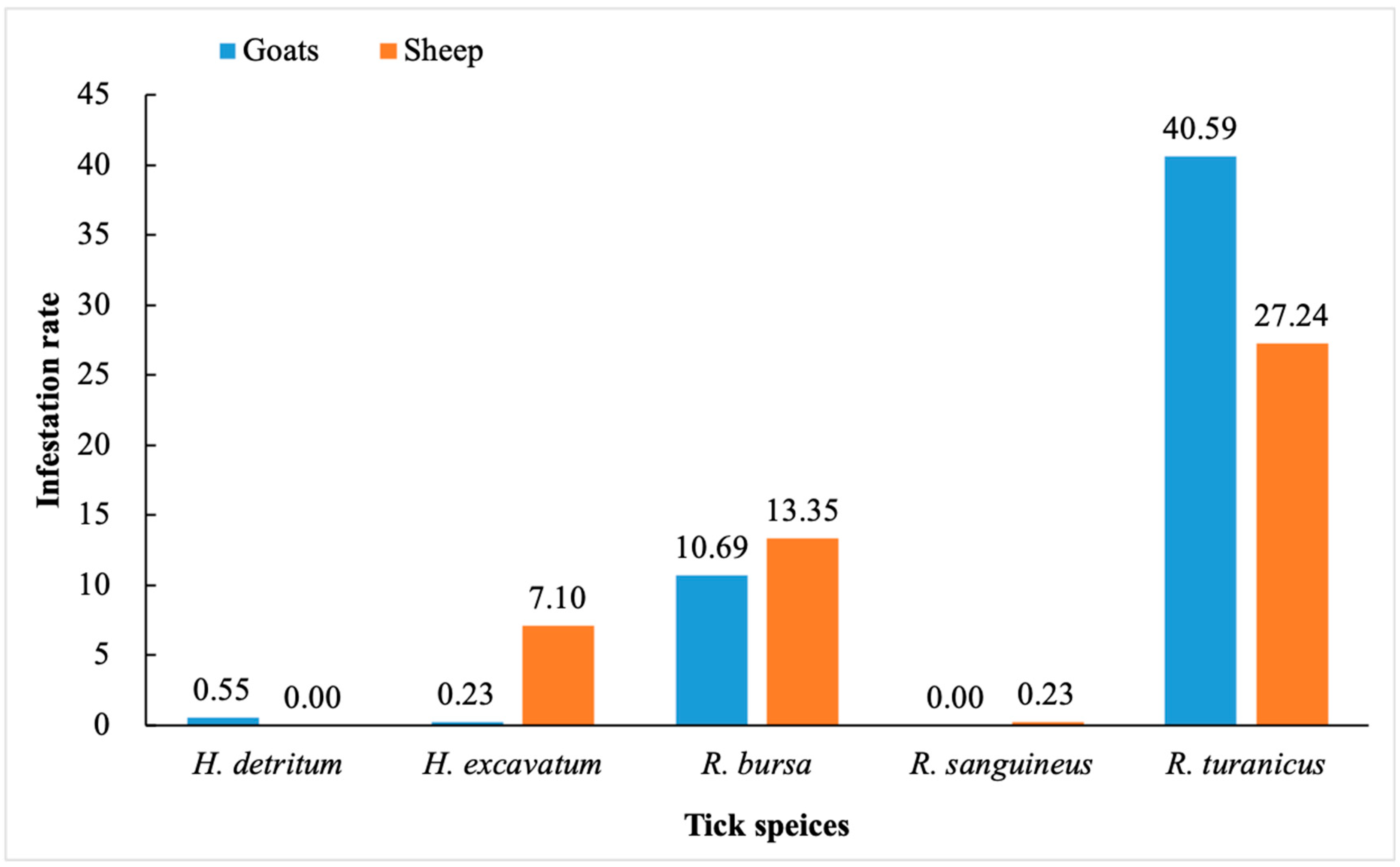
3.1. Tick Species Identification and Infestation Rate
3.2. Detection of Piroplasms in Ticks
3.3. Phylogenetic Analyses of B. ovis and T. ovis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Uilenberg, G. International collaborative research: Significance of tick-borne hemoparasitic diseases to world animal health. Vet. Parasitol. 1995, 57, 19–41. [Google Scholar] [CrossRef]
- Minjauw, B.; McLeod, A. Tick-borne diseases and poverty: The impact of ticks and tick-borne diseases on the livelihoods of small-scale and marginal livestock owners in India and Eastern and Southern Africa. In Tick-Borne Diseases and Poverty: The Impact of Ticks and Tick-Borne Diseases on the Livelihoods of Small-Scale and Marginal Livestock Owners in India and Eastern and Southern Africa; DFID Animal Health Programme, Centre for Tropical Veterinary Medicine: Edinburgh, UK, 2003. [Google Scholar]
- McLeod, R.; Kristjanson, P. Economic Impact of Ticks and Tick-Borne Diseases to Livestock in Africa, Asia and Australia; International Livestock Research Institute: Nairobi, Kenya, 1999. [Google Scholar]
- Ceylan, O.; Xuan, X.; Sevinc, F. Primary tick-borne protozoan and rickettsial infections of animals in Turkey. Pathogens 2021, 10, 231. [Google Scholar] [CrossRef] [PubMed]
- Minami, T.; Fujinaga, T.; Furuya, K.; Ishihara, T. Clinico-hematologic and serological comparison of Japanese and Russian strains of Theileria sergenti. Natl. Inst. Anim. Health Q. 1980, 20, 44–52. [Google Scholar]
- Bock, R.; Jackson, L.; de Vos, A.; Jorgensen, W. Babesiosis of cattle. Parasitology 2004, 129, S247–S269. [Google Scholar] [CrossRef] [PubMed]
- Sivakumar, T.; Tuvshintulga, B.; Otgonsuren, D.; Batmagnai, E.; Ahedor, B.; Kothalawala, H.; Vimalakumar, S.C.; Silva, S.S.P.; Yamagishi, J.; Yokoyama, N. Phylogenetic analyses of the mitochondrial, plastid, and nuclear genes of Babesia sp. Mymensingh and its naming as Babesia naoakii n. sp. Parasit. Vectors 2022, 15, 299. [Google Scholar] [CrossRef] [PubMed]
- Bilgic, H.B.; Bakırcı, S.; Kose, O.; Unlu, A.H.; Hacılarlıoglu, S.; Eren, H.; Weir, W.; Karagenc, T. Prevalence of tick-borne haemoparasites in small ruminants in Turkey and diagnostic sensitivity of single-PCR and RLB. Parasit. Vectors 2017, 10, 211. [Google Scholar] [CrossRef] [PubMed]
- Morel, P.C.; Uilenberg, G. The nomenclature of some Theileria species (Sporozoa, Babesioidea) of domestic ruminants (Author’s Transl). Rev. Elev. Med. Vet. Pays. Trop. 1981, 34, 139–143. [Google Scholar]
- Aktaş, M.; Altay, K.; Dumanli, N. Development of a polymerase chain reaction method for diagnosis of Babesia ovis infection in sheep and goats. Vet. Parasitol. 2005, 133, 277–281. [Google Scholar] [CrossRef]
- Liu, A.H.; Yin, H.; Guan, G.Q.; Schnittger, L.; Liu, Z.J.; Ma, M.L.; Dang, Z.S.; Liu, J.L.; Ren, Q.Y.; Bai, Q.; et al. At least two genetically distinct large Babesia species infective to sheep and goats in China. Vet. Parasitol. 2007, 147, 246–251. [Google Scholar] [CrossRef]
- Köseoğlu, A.E.; Can, H.; Güvendi, M.; Erkunt Alak, S.; Kandemir, Ç.; Taşkın, T.; Demir, S.; Akgül, G.; Değirmenci Döşkaya, A.; Karakavuk, M.; et al. Molecular investigation of bacterial and protozoal pathogens in ticks collected from different hosts in Turkey. Parasit. Vectors 2021, 14, 270. [Google Scholar] [CrossRef]
- De la Fuente, J.; Estrada-Pena, A.; Venzal, J.M.; Kocan, K.M.; Sonenshine, D.E. Overview: Ticks as vectors of pathogens that cause disease in humans and animals. Front. Biosci. 2008, 13, 6938–6946. [Google Scholar] [CrossRef] [PubMed]
- Nava, S.; Guglielmone, A.A.; Mangold, A.J. An overview of systematics and evolution of ticks. Front. Biosci. 2009, 14, 2857–2877. [Google Scholar] [CrossRef] [PubMed]
- Reuben Kaufman, W. Ticks: Physiological aspects with implications for pathogen transmission. Ticks Tick Borne Dis. 2010, 1, 11–22. [Google Scholar] [CrossRef]
- Guo, H.; Adjou Moumouni, P.F.; Thekisoe, O.; Gao, Y.; Liu, M.; Li, J.; Galon, E.M.; Efstratiou, A.; Wang, G.; Jirapattharasate, C.; et al. Genetic characterization of tick-borne pathogens in ticks infesting cattle and sheep from three South African provinces. Ticks Tick Borne Dis. 2019, 10, 875–882. [Google Scholar] [CrossRef] [PubMed]
- Teng, Z.; Shi, Y.; Zhao, N.; Zhang, X.; Jin, X.; He, J.; Xu, B.; Qin, T. Molecular detection of tick-borne bacterial and protozoan pathogens in Haemaphysalis longicornis (Acari: Ixodidae) ticks from free-ranging domestic sheep in Hebei Province, China. Pathogens 2023, 12, 763. [Google Scholar] [CrossRef] [PubMed]
- Guglielmone, A.A.; Robbins, R.G.; Apanaskevich, D.A.; Petney, T.N.; Estrada-peña, A.; Horak, I.G.; Shao, R.; Barker, S.C. The Argasidae, Ixodidae and Nuttalliellidae (Acari: Ixodida) of the world: A list of valid species names. Zootaxa 2010, 2528, 1–28. [Google Scholar] [CrossRef]
- Randolph, S.E.; Green, R.M.; Hoodless, A.N.; Peacey, M.F. An empirical quantitative framework for the seasonal population dynamics of the tick Ixodes ricinus. Int. J. Parasitol. 2002, 32, 979–989. [Google Scholar] [CrossRef]
- Guglielmone, A.A. The seasonal occurrence of Amblyomma triguttatum triguttatum Koch (Acari, Ixodidae). Acarologia 1994, 35, 107–113. [Google Scholar]
- Abunna, F.; Kasasa, D.; Shelima, B.; Megersa, B.; Regassa, A.; Amenu, K. Survey of tick infestation in small ruminants of Miesso district, West Harergie, Oromia Region, Ethiopia. Trop Anim. Health Prod. 2009, 41, 969–972. [Google Scholar] [CrossRef]
- Sevinc, F.; Xuan, X. Major tick-borne parasitic diseases of animals: A frame of references in Turkey. Eurasian J. Vet. Sci. 2015, 31, 132. [Google Scholar] [CrossRef]
- Turkish Statistical Institute. Available online: http://www.turkstat.gov.tr/ (accessed on 9 February 2023).
- Bursali, A.; Keskin, A.; Tekin, S. A review of the ticks (Acari: Ixodida) of Turkey: Species diversity, hosts and geographical distribution. Exp. Appl. Acarol. 2012, 57, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Ji, S.; Ceylan, O.; Ma, Z.; Galon, E.M.; Zafar, I.; Li, H.; Hasegawa, Y.; Sevinc, M.; Masatani, T.; Iguchi, A.; et al. Protozoan and rickettsial pathogens in ticks collected from infested cattle from Turkey. Pathogens 2022, 11, 500. [Google Scholar] [CrossRef] [PubMed]
- Aktas, M. A survey of ixodid tick species and molecular identification of tick-borne pathogens. Vet. Parasitol. 2014, 200, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Aktas, M.; Altay, K.; Dumanli, N.; Kalkan, A. Molecular detection and identification of Ehrlichia and Anaplasma species in ixodid ticks. Parasitol. Res. 2009, 104, 1243–1248. [Google Scholar] [CrossRef] [PubMed]
- Aktas, M.; Altay, K.; Ozubek, S.; Dumanli, N. A survey of ixodid ticks feeding on cattle and prevalence of tick-borne pathogens in the Black Sea Region of Turkey. Vet. Parasitol. 2012, 187, 567–571. [Google Scholar] [CrossRef]
- Aktas, M.; Altay, K.; Dumanli, N. PCR-based detection of Theileria ovis in Rhipicephalus bursa adult ticks. Vet. Parasitol. 2006, 140, 259–263. [Google Scholar] [CrossRef]
- Aktas, M.; Ozübek, S.; Ipek, D.N.S. Molecular investigations of Hepatozoon species in dogs and developmental stages of Rhipicephalus sanguineus. Parasitol. Res. 2013, 112, 2381–2385. [Google Scholar] [CrossRef]
- Altay, K.; Aktas, M.; Dumanli, N. Detection of Babesia ovis by PCR in Rhipicephalus bursa collected from naturally infested sheep and goats. Res. Vet. Sci. 2008, 85, 116–119. [Google Scholar] [CrossRef]
- Aydin, M.; Aktas, M.; Dumanli, N. Tick infestations on sheep and goats in the Black Sea Region of Türkiye. Kafkas Univ. Vet. Fak. Derg. 2012, 18, A17–A22. [Google Scholar] [CrossRef]
- Ozubek, S.; Aktas, M. Molecular and parasitological survey of ovine piroplasmosis, including the first report of Theileria annulata (Apicomplexa: Theileridae) in sheep and goats from Turkey. J. Med. Entomol. 2017, 54, 212–220. [Google Scholar] [CrossRef]
- Benedicto, B.; Ceylan, O.; Moumouni, P.F.A.; Lee, S.-H.; Tumwebaze, M.A.; Li, J.; Galon, E.M.; Liu, M.; Li, Y.; Ji, S.; et al. Molecular detection and assessment of risk factors for tick-borne diseases in sheep and goats from Turkey. Acta Parasitol. 2020, 65, 723–732. [Google Scholar] [CrossRef] [PubMed]
- Ceylan, O.; Byamukama, B.; Ceylan, C.; Galon, E.M.; Liu, M.; Masatani, T.; Xuan, X.; Sevinc, F. Tick-borne hemoparasites of sheep: A molecular research in Turkey. Pathogens 2021, 10, 162. [Google Scholar] [CrossRef] [PubMed]
- Sevinc, F.; Zhou, M.; Cao, S.; Ceylan, O.; Aydin, M.F.; Sevinc, M.; Xuan, X. Haemoparasitic agents associated with ovine babesiosis: A possible negative interaction between Babesia ovis and Theileria ovis. Vet. Parasitol. 2018, 252, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Horak, I.G.; Heyne, H.; Williams, R.; Gallivan, G.J.; Spickett, A.M.; Bezuidenhout, J.D.; Estrada-Peña, A. The Ixodid Ticks (Acari: Ixodidae) of Southern Africa; Springer International Publishing: Cham, Switzerland, 2018; ISBN 978-3-319-70640-5. [Google Scholar]
- Adjou Moumouni, P.F.; Terkawi, M.A.; Jirapattharasate, C.; Cao, S.; Liu, M.; Nakao, R.; Umemiya-Shirafuji, R.; Yokoyama, N.; Sugimoto, C.; Fujisaki, K.; et al. Molecular detection of spotted fever group rickettsiae in Amblyomma variegatum ticks from Benin. Ticks Tick Borne Dis. 2016, 7, 828–833. [Google Scholar] [CrossRef]
- Casati, S.; Sager, H.; Gern, L.; Piffaretti, J.-C. Presence of potentially pathogenic Babesia sp. for Human in Ixodes ricinus in Switzerland. Ann. Agric. Environ. Med. 2006, 13, 65–70. [Google Scholar]
- McLure, A.; O’Neill, B.; Mayfield, H.; Lau, C.; McPherson, B. PoolTestR: An R package for estimating prevalence and regression modelling for molecular xenomonitoring and other applications with pooled samples. Environ. Model. Softw. 2021, 145, 105158. [Google Scholar] [CrossRef]
- Sivakumar, T.; Tuvshintulga, B.; Zhyldyz, A.; Kothalawala, H.; Yapa, P.R.; Kanagaratnam, R.; Vimalakumar, S.C.; Abeysekera, T.S.; Weerasingha, A.S.; Yamagishi, J.; et al. Genetic analysis of Babesia isolates from cattle with clinical babesiosis in Sri Lanka. J. Clin. Microbiol. 2018, 56, 10–1128. [Google Scholar] [CrossRef]
- Kirvar, E.; Ilhan, T.; Katzer, F.; Wilkie, G.; Hooshmand-Rad, P.; Brown, D. Detection of Theileria lestoquardi (hirci) in ticks, sheep, and goats using the polymerase chain reaction. Ann. N. Y. Acad. Sci. 1998, 849, 52–62. [Google Scholar] [CrossRef]
- Yin, H.; Liu, Z.; Guan, G.; Liu, A.; Ma, M.; Ren, Q.; Luo, J. Detection and differentiation of Theileria luwenshuni and T. uilenbergi infection in small ruminants by PCR. Transbound. Emerg. Dis. 2008, 55, 233–237. [Google Scholar] [CrossRef]
- Altay, K.; Dumanli, N.; Holman, P.J.; Aktas, M. Detection of Theileria ovis in naturally infected sheep by nested PCR. Vet. Parasitol. 2005, 127, 99–104. [Google Scholar] [CrossRef]
- Inci, A.; Yildirim, A.; Duzlu, O.; Doganay, M.; Aksoy, S. Tick-borne diseases in Turkey: A review based on one health perspective. PLoS Negl. Trop Dis. 2016, 10, e0005021. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Peña, A.; Ayllón, N.; de la Fuente, J. Impact of climate trends on tick-borne pathogen transmission. Front. Physiol. 2012, 3, 64. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Z.; Mu, L.M.; Zhang, K.; Yang, M.H.; Zhang, L.; Du, J.Y.; Liu, Z.Q.; Li, Y.X.; Lu, W.H.; Chen, C.F.; et al. A broad-range survey of ticks from livestock in Northern Xinjiang: Changes in tick distribution and the isolation of Borrelia burgdorferi sensu stricto. Parasit. Vectors 2015, 8, 449. [Google Scholar] [CrossRef] [PubMed]
- Ceylan, O.; Uslu, A.; Ceylan, C.; Sevinc, F. Predominancy of Rhipicephalus turanicus in tick-infested sheep from Turkey: A large-scale survey. Pak. Vet. J. 2021, 41, 2074–7764. [Google Scholar] [CrossRef]
- Uilenberg, G. Babesia—A historical overview. Vet. Parasitol. 2006, 138, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Bai, Q.; Liu, G.; Liu, D.; Ren, J.; Li, X. Isolation and preliminary characterization of a large Babesia sp. from sheep and goats in the Eastern part of Gansu Province, China. Parasitol. Res. 2002, 88, S16–S21. [Google Scholar] [CrossRef]
- Sevinc, F.; Sevinc, M.; Ekici, O.D.; Yildiz, R.; Isik, N.; Aydogdu, U. Babesia ovis infections: Detailed clinical and laboratory observations in the pre-and post-treatment periods of 97 field cases. Vet. Parasitol. 2013, 191, 35–43. [Google Scholar] [CrossRef]
- Ceylan, O.; Sevinc, F. Endemic instability of ovine babesiosis in Turkey: A country-wide sero-epidemiological study. Vet. Parasitol. 2020, 278, 109034. [Google Scholar] [CrossRef]
- Altay, K.; Dumanli, N.; Aktas, M. A study on ovine tick-borne hemoprotozoan parasites (Theileria and Babesia) in the East Black Sea Region of Turkey. Parasitol. Res. 2012, 111, 149–153. [Google Scholar] [CrossRef]
- Zhou, M.; Cao, S.; Sevinc, F.; Sevinc, M.; Ceylan, O.; Ekici, S.; Jirapattharasate, C.; Moumouni, P.F.A.; Liu, M.; Wang, G.; et al. Molecular detection and genetic characterization of Babesia, Theileria and Anaplasma amongst apparently healthy sheep and goats in the Central Region of Turkey. Ticks Tick Borne Dis. 2017, 8, 246–252. [Google Scholar] [CrossRef]
- Aydin, M.F.; Aktas, M.; Dumanli, N. Molecular identification of Theileria and Babesia in ticks collected from sheep and goats in the Black Sea Region of Turkey. Parasitol. Res. 2015, 114, 65–69. [Google Scholar] [CrossRef]
- Friedhoff, K. Tick-borne diseases of sheep and goats caused by Babesia, Theileria or Anaplasma spp. Parasitologia 1997, 39, 99–109. [Google Scholar]
- Ozubek, S.; Aktas, M. Molecular evidence for a novel species of Babesia in unfed Rhipicephalus sanguineus sensu lato (Acari: Ixodidae). J. Med. Entomol. 2018, 55, 1271–1276. [Google Scholar] [CrossRef] [PubMed]
- Hamid, P.H.; Cahyadi, M.; Wardhana, A.H.; Sawitri, D.H.; Setya, N.N.R.; Insyariati, T.; Kurnianto, H.; Hermosilla, C.R. First autochthonous report on cattle Babesia naoakii in Central Java, Indonesia, and identification of Haemaphysalis bispinosa ticks in the investigated area. Pathogens 2022, 12, 59. [Google Scholar] [CrossRef] [PubMed]
- Futse, J.E.; Ueti, M.W.; Knowles, D.P.; Palmer, G.H. Transmission of Anaplasma marginale by Boophilus microplus: Retention of vector competence in the absence of vector-pathogen interaction. J. Clin. Microbiol. 2003, 41, 3829–3834. [Google Scholar] [CrossRef] [PubMed]
- Ringo, A.E.; Aboge, G.O.; Adjou Moumouni, P.F.; Hun Lee, S.; Jirapattharasate, C.; Liu, M.; Gao, Y.; Guo, H.; Zheng, W.; Efstratiou, A.; et al. Molecular detection and genetic characterisation of pathogenic Theileria, Anaplasma and Ehrlichia species among apparently healthy sheep in Central and Western Kenya. Onderstepoort J. Vet. Res. 2019, 86, e1–e8. [Google Scholar] [CrossRef]
- Aktaş, M.; Altay, K.; Dumanli, N. Survey of Theileria parasites of sheep in eastern Turkey using polymerase chain reaction. Small Rumin. Res. 2005, 60, 289–293. [Google Scholar] [CrossRef]
- Ahmed, J.; Yin, H.; Bakheit, M.; Liu, Z.; Mehlhorn, H.; Seitzer, U. Small ruminant theileriosis. Prog. Parasitol. 2011, 2, 135–153. [Google Scholar] [CrossRef]
- Büscher, G.; Friedhoff, K.T.; El-Allawy, T.A.A. Quantitative Description of the development of Babesia ovis in Rhipicephalus bursa (hemolymph, ovary, eggs). Parasitol. Res. 1988, 74, 331–339. [Google Scholar] [CrossRef]
- Neitz, W.O. The experimental transmission of Theileria ovis by Rhipicephalus evertsi mimeticus and R. bursa. Onderstepoort J. Vet. Res. 1972, 39, 83–86. [Google Scholar]



| District | Tick Species (No. of Tick/No. of Tick Pool) | B. ovis (No. of Tick/No. of Tick Pool) | T. ovis (No. of Tick/No. of Tick Pool) |
|---|---|---|---|
| Kadınhanı | R. turanicus (547/89) | - | 12/89 (2.33% a) CI b (1.26–3.88) |
| Karatay | R. bursa (3/1) | 1/1 | - |
| R. sanguineus (1/1) | 1/1 | - | |
| Sarayönü | R. bursa (127/40) | 7/40 (6.24%) CI (2.72–11.80) | - |
| R. sanguineus (39/12) | - | - | |
| Selçuklu | H. detritum (7/7) | - | - |
| H. excavatum (94/60) | - | - | |
| R. bursa (178/77) | 21/77 (13.50%) CI (8.73–19.60) | 1/77 (0.56%) CI (0.03–2.45) | |
| R. sanguineus (2/2) | - | - | |
| R. turanicus (283/68) | 1/68 (0.36%) CI (0.02–1.59) | 5/68 (1.90%) CI (0.69–4.04) | |
| Total | R. bursa (308/118) | 29/118 (10.80%) CI (7.43–14.90) | 1/118 (0.33%) CI (0.02–1.42) |
| R. turanicus (869/169) | 1/169 (0.12%) CI (0.01–0.51) | 17/169 (2.08%) CI (1.25–3.22) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Z.; Ceylan, O.; Galon, E.M.; Mohanta, U.K.; Ji, S.; Li, H.; Do, T.T.; Umemiya-Shirafuji, R.; El-Sayed, S.A.E.-S.; Zafar, I.; et al. Molecular Identification of Piroplasmids in Ticks from Infested Small Ruminants in Konya Province, Turkey. Pathogens 2023, 12, 1123. https://doi.org/10.3390/pathogens12091123
Ma Z, Ceylan O, Galon EM, Mohanta UK, Ji S, Li H, Do TT, Umemiya-Shirafuji R, El-Sayed SAE-S, Zafar I, et al. Molecular Identification of Piroplasmids in Ticks from Infested Small Ruminants in Konya Province, Turkey. Pathogens. 2023; 12(9):1123. https://doi.org/10.3390/pathogens12091123
Chicago/Turabian StyleMa, Zhuowei, Onur Ceylan, Eloiza May Galon, Uday Kumar Mohanta, Shengwei Ji, Hang Li, Thanh Thom Do, Rika Umemiya-Shirafuji, Shimaa Abd El-Salam El-Sayed, Iqra Zafar, and et al. 2023. "Molecular Identification of Piroplasmids in Ticks from Infested Small Ruminants in Konya Province, Turkey" Pathogens 12, no. 9: 1123. https://doi.org/10.3390/pathogens12091123
APA StyleMa, Z., Ceylan, O., Galon, E. M., Mohanta, U. K., Ji, S., Li, H., Do, T. T., Umemiya-Shirafuji, R., El-Sayed, S. A. E.-S., Zafar, I., Liu, M., Sevinc, F., & Xuan, X. (2023). Molecular Identification of Piroplasmids in Ticks from Infested Small Ruminants in Konya Province, Turkey. Pathogens, 12(9), 1123. https://doi.org/10.3390/pathogens12091123