Abstract
Vesicular stomatitis virus (VSV) is an emergent virus affecting livestock in the US. Previously, using a recombinant VSV carrying the M51R mutation in the matrix protein (rNJ0612NME6-M51R), we evaluated the pathogenesis of this virus in pigs. Our results indicated that rNJ0612NME6-M51R represented an attenuated phenotype in in-vivo and in ex-vivo in pig macrophages, resembling certain clinical features observed in field VSV isolates. In order to gain more insight into the molecular basis leading to the attenuation of rNJ0612NME6-M51R in pigs, we conducted a microarray analysis to assess the gene expression profiles of primary porcine macrophages infected with rNJ0612NME6-M51R compared to its parental virus (rNJ0612NME6). Our results showed an overall higher gene expression in macrophages infected with rNJ0612NME6-M51R. Specifically, we observed that the pathways related with immune cytokine signaling and interferon (IFN)-related responses (including activation, signaling, induction, and antiviral mechanisms) were the ones comprising most of the relevant genes identified during this study. Collectively, the results presented herein highlight the relevance of type I interferon during the pathogenesis of VSV in pigs. The information generated from this study may represent a framework for future studies intended to understand the molecular bases of the pathogenesis of field strains in livestock.
1. Introduction
Vesicular stomatitis (VS) is an arboviral disease that affects livestock [1,2]. The disease is caused by the vesicular stomatitis virus (VSV), which is a negative-sense single-stranded RNA virus, and a prototype of the Vesiculovirus genus in the Rhabdoviridae family [3]. This genus encompasses a total of 19 species, among them, Vesiculovirus New Jersey (named as vesicular stomatitis New Jersey virus, or VSNJV) and Vesiculovirus Indiana (vesicular stomatitis Indiana, or VSIV) are responsible for the majority of clinical cases of VS in livestock in the Americas [4]. The genome of VSV encodes five structural proteins: nucleoprotein, phosphoprotein, matrix protein, glycoprotein, and a large polymerase. The matrix protein is one of the most-studied VSV proteins, and is known for its function as a key regulator of the innate immune response [5,6,7].
While advances in our understanding of VSV biology have led to the development of recombinant VSVs with oncolytic properties [8], and as a viral-vectored vaccine platform [9,10], there are still important aspects of the virus that remain poorly understood. Understanding the molecular mechanisms underlying VSV pathogenesis is critical for grasping the extraordinary capacity of the virus to cause cyclic emergence and produce large epidemic outbreaks in the USA [11,12,13,14], making it highly relevant for the livestock industry.
VSV infection in livestock arises through biting insects that carry the virus [15], or by direct contact with an infected animal [16]. The infection causes a disease that is characterized by the development of vesicular lesions in the oral mucosa, tongue, teats, and coronary bands of affected animals [17]. Given their high susceptibility to VSV infection, pigs, which are natural hosts of the virus, have been widely used as an experimental model for studying VSV pathogenesis [16,18,19,20,21,22,23]. The pig model can also be utilized to predict the virulence of VSV (field strains or recombinant viruses) by assessing its ability to induce secondary vesicular lesions in pigs after an initial inoculation via scarification in the snout or direct animal contact [16,23], along with monitoring typical parameters of virulence such as the development of fever and RNAemia [16]. Based on these parameters, it has been typically assumed that VSNJV is more virulent than VSIV due to differences in viral transmission via direct contact and the development of secondary vesicular lesions serving as the main markers of virulence [24]. However, a recent study has shown that certain VSIV strains are comparable in virulence to the VSNJV strains [19]. This suggests that specific genetic determinants among VSIV strains may mediate virulence. Furthermore, a pathogenesis study comparing the virulence between the epidemic strain (NJ0612NME6) and its closest endemic relative, NJ0608VCB, showed that epidemic strains of VSNJV exhibit a more virulent phenotype than endemic ones [16]. Interestingly, the increased virulence observed in NJ0612NME6 was correlated not only with an increased number of vesicular lesions, but also with decreased levels of type I interferon (IFN) in the infected pigs. This observation highlighted the ability for some VSV field strains to suppress the type I IFN response, and represents a potential marker of virulence in pigs.
Recently, we genetically engineered the highly virulent epidemic strain NJ0612NME6 to carry a M51R mutation in the M protein (rNJ0612NME6-M51R) [23]. This mutation has been demonstrated to disrupt the virus’s ability to block the nuclear–cytoplasmic transport of mRNA, which is required for type I IFN production in infected cells [5,25,26]. Interestingly, proof-of-concept studies conducted in swine resulted in an attenuated phenotype. The infection produced vesicular lesions at the inoculation site, but the capacity to produce secondary vesicular lesions and other clinical signs of virulence was decreased (Figure 1). The decreased replication capacity of rNJ0612NME6-M51R in pigs was found to be correlated with reduced viral replication in cultured primary porcine macrophages [23], suggesting an important role for these cells in VSV pathogenesis. Our recent study provides additional evidence for the critical role of porcine macrophages in VSV pathogenesis. Specifically, we identified a subset of differentially expressed genes (DEGs) involved in various immune pathways between primary swine macrophages infected with the field isolate and mock infection in vitro, shedding light on the molecular mechanisms underlying VSV infection [27].
Figure 1.
Phenotypic characteristics displayed by rNJ0612NME6-M51R during its infection in pigs. Contrasting clinical scores were observed between pigs infected by direct inoculation (IN) and contact exposure (CO) with rNJ0612NME6 and rNJ0612NME6-M51R, indicating the reduced ability of rNJ0612NME6-M51R to produce the development of secondary vesicular lesions after infection, a situation that resembles some clinical outcomes produced by VSIV strains. The attenuated phenotype showed by rNJ0612NME6-M51R was also correlated with the absence of fever and RNAemia in IN and CO pigs infected with this virus. The presence of viral RNA in the spleen of pigs infected with rNJ0612NME6-M51R indicates the ability of this virus to produce systemic infection. However, in both groups no infectious virus was recovered from post-mortem tissues. Interestingly. The lack of correlation between the presence of RNA in spleen samples from pigs infected with M51R, may be explained based on the decreased number of vesicular lesions produced in these pigs. The correlation between the intensity of the RNAemia and the number of vesicular lesion in infected animals has been previously reported [16]. The relevance of RNAemia in the pathogenesis of VSV, is currently under investigation. Based on these results, we are proposing the use of rNJ0612NME6-M51R as a potential model to obtain more insight into the molecular basis of the pathogenies of VSV in pigs. Data from this figure were obtained from Velazquez-Salinas et al. [23]. This figure was created using BioRender.com, under the academic license number: QX25JBPWHS.
In this study, we aimed to compare host mRNA expression profiles in primary porcine macrophages infected with either the highly virulent strain (WT) or the attenuated strain (M51R) of VSV. Our objective was to identify DEGs and associated biological pathways that may provide insights into the molecular basis mediating the virulence of VSV in pigs. The analysis of our study unveiled several immune-related pathways that were significantly over-represented and predominantly linked with type I interferon (IFN) responses. These findings deepen our understanding of VSV pathogenesis, and could facilitate future in vitro studies to characterize VSV field strains.
2. Materials and Methods
2.1. Cells and Viruses
Primary swine macrophages were prepared from defibrinated blood, as previously described [28]. The field isolate virus, rNJ0612NME6, and recombinant virus, rNJ0612NME6-M51R, were obtained from the high titer viral stocks produced in Vero cells during a previous study [23]. Both viruses were derived from our infectious clone system [29]. In the case of rNJ0612NME6-M51R, mutation at M51R in the matrix protein was engineered via site-directed mutagenesis, as previously described [23].
2.2. RNA Extraction
Infections with rNJ0612NME6 and rNJ0612NME6-M51R were carried out on primary swine porcine macrophages using three independent biological replicates collected from three pigs. Six-well plates containing 1 × 107 cells per well were infected at an MOI of 10 TCID50 with each virus, and incubated for five hours at 37 °C under 5% of CO2. The infections were conducted in triplicate. The mock-infected samples were incubated in the same conditions with macrophage media in triplicate. The incubation time was determined based on previous studies performed on primary porcine macrophages [22,23,27]. It is supported by the ability of VSV to induce high levels of gene expression in primary porcine macrophages infected with a high MOI, avoiding the presence of the cytopathic effect, a condition that decreases the RNA yields. After incubation, the total RNA was extracted using the RNeasy mini kit (Qiagen LLC-USA, Germantown, MD, USA cat# 74004) according to the manufacturer’s instructions. RNA quality was assessed using the Agilent 2100 bioanalyzer (Santa Clara, CA, USA), using an RNA nanochip. The quantification of RNA was conducted on a Nanodrop 1000 (Thermo Scientific, Rockwood, TN, USA).
2.3. ELISA
A commercial enzyme-linked immunosorbent assay (ELISA) (Abcam porcine IFN-alpha ELISA kit, Cat #ab273213) was used to compare the abilities of rNJ0612NME6 and rNJ0612NME6-M51R to induce the secretion of porcine type I IFN (IFNα) in primary swine porcine macrophages. For this purpose, six-well plates containing 1 × 107 cells per well were infected at an MOI of 10 TCID50, with each virus and incubated for five hours at 37 °C under 5% of CO2. Supernatants were collected and evaluated via ELISA, following the manufacturer’s protocol. The experiments were conducted in three biological replicates.
2.4. DNA Microarray Analysis
Microarray analyses were conducted using a previously described porcine microarray [27,30]. The porcine microarrays were custom designed and provided by Agilent Technologies. The RNA samples were labeled with Cy3 and Cy5 individually, using a low-input RNA labeling kit (Agilent Technologies cat# Part # 5190-2306). The Cy5-labeled RNA samples were co-hybridized with the corresponding Cy3-labeled sample (e.g., VSV and mock-infected) using a dye-swap design. Array slides were scanned using a GenePix 4000B scanner (Molecular Devices) with the GenePix Pro 6.0 software (Molecular Devices) at 5 μM resolution. The procedure and analysis were conducted according to manufacturers’ instructions.
The LIMMA package included in the R software was used to conduct data normalization, background correction, and statistical analysis [31]. Log2-fold changes in signal intensity were used to identify differentially expressed genes between the VSV and mock-infected samples. The Benjamini and Hochberg method was used to adjust p-values that were ultimately expressed as a false discovery rate (FDR). In this sense, genes showing FDR values of ≤0.05 and an expression difference of ≥50% were considered to be significant differentially expressed genes (DEGs) between the VSV-infected and mock-infected samples. A validation with qPCR of some of the DEGs reported in this study is provided in Supplementary File S1. This experiment was conducted using three biological replicates, following a methodology previously described [23].
2.5. Hierarchical Cluster Analysis
Hierarchical cluster analysis (HCA) was used to categorize the correlation of selected DEGs between rNJ0612NME6 and rNJ0612NME6-M51R. HCA uses a matrix of N × M dimensions, where N is the number of DEGs and M is the number of viruses used in the analysis. The overall data were robustly standardized, and clusters were defined by the complete linkage method. Once the number of clusters was defined, an analysis of variance (ANOVA) supported by Tukey’s honest significance test (a = 0.05) was conducted to identify significant clusters in terms of the differential in the levels of gene expressions between rNJ0612NME6 and rNJ0612NME6-M51R. The analyses were conducted using the predictive analytic software JMP® Pro version 16.0.0.
2.6. Pathway Analysis
Pig gene sequences of microarray probes were mapped to human genes using BLAST and/or manually annotated with genetic information displayed on the UCSC Genome Browser website. Human gene Entrez-ID numbers associated with DEG were used in the pathway analysis using the database for annotation, visualization, and integrated discovery (DAVID), v6.8 updated 2021 version [32]. The biological pathways were identified using the reactome pathway analysis [33], considering an FDR of ≤0.05 as the significant threshold, and the human database as a model for the analyses.
2.7. Euler Diagram
An Euler diagram was used to represent the relationship among genes of multiple identified pathways. This diagram is frequently used for the analysis of microarray experiments [34], and highlights relevant genes shared by different pathways reflecting potential functional associations between them. These associations are represented by closed circles that split the plane into divided but connected subcategories [35]. The analysis was conducted using the R package eulerr.
2.8. Biological Inference
The biological functions of the identified DEGs in the identified in over-represented pathways associated with the immune response were inferred based on scientific publications literature obtained from PubMed (https://pubmed.ncbi.nlm.nih.gov/ accessed on 1 January 2023). The biological inferences were based on (i) the immunological functions of the DEGs, (ii) the signal intensities of gene expression levels based on microarray-averaged signal intensities, and (iii) magnitudes (fold) of the differential expressions, assuming that higher mean signal intensities and therefore larger differentially expressed genes play a more significant bigger biological role in the gene groups. Genes with no significant differential expression, but that are known to play important roles in the biological pathways associated with the significant DEGs, but without differential expression themselves, were used as supporting evidence. Genes that were down- or up-regulated in the VSV-infected samples compared to the mock-infected samples were expressed as negative and positive values (fold), respectively. In this study, the genes that were differentially expressed between infected and mock-infected macrophages were used to further infer the molecular mechanisms of VSV pathogenesis and immune evasion.
3. Results
3.1. Clustering Analysis of DEGs
A total of 4024 DEGs (up- or down-regulated) between macrophages infected with either rNJ0612NME6 or rNJ0612NME6-M51R and the mock-infected cells were detected. HCA was used to group these DEGs into 20 clusters (Figure 2A), with cluster numbers 2 and 15 comprising 70.6% of the total DEGs. In these two clusters, the mean differential expressions between the two infected samples were only 0.77- and 0.43-fold, respectively, indicating that most of the identified genes were similarly expressed (Figure 2B). In contrast, a statistically significant difference between the two infected samples was found in six clusters (4, 9, 10, 11, 13 and 17) identified by ANOVA, which was further confirmed by the Tukey’s honest significance test (a = 0.05). These clusters displayed the highest differential expression means between the two infected samples compared to the other clusters. These six clusters consisted of a total of 106 genes (2.63% of the DEGs), with the differential expression means ranging from 6.13- and 30.19-fold (Figure 2B). In addition, another group of relevant clusters (1, 3, 5, 12, 14, 16, 20) comprising a total of 718 DEGs was identified, with differential mean gene expressions ranging from 2.20- to 3.98-fold between the two infected samples. Given their significant differential expression profiles, these two groups of clusters were further analyzed using pathway analysis. The remaining clusters containing a mean differential expression value lower than 2.20-fold were excluded from the identification pathway analysis (clusters 2, 6, 7, 8, 15, 18, 19).
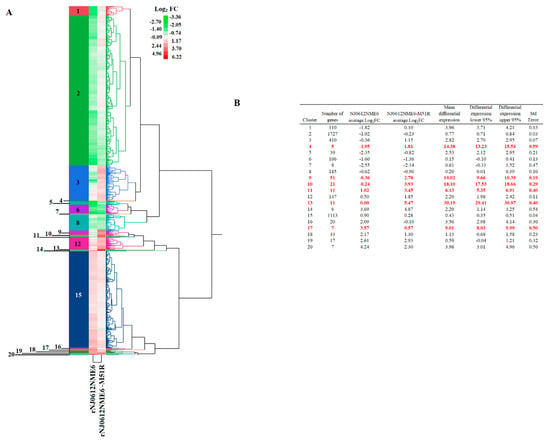
Figure 2.
Hierarchical cluster analysis (HCA) of differentially expressed transcripts. (A) HCA was conducted to identify relevant groups of genes differentially expressed in cell cultures of primary swine macrophages after infection with either rNJ0612NME6 or rNJ0612NME6-M51R. Levels of expression are displayed in terms of fold change (FC) Log2. Negative and positive values reflect genes that were down- or up-regulated, respectively. (B) Summary of the statistics displayed by different clusters. Numbers in red represent mean differential expressions that were significantly different from the rest of the clusters. Calculations were determined with ANOVA and supported by Tukey’s honest significance test.
3.2. DEG Pathway Analysis
The DAVID functional annotation tool was used to analyze the 13 selected gene clusters (1, 3–5, 9–14, 16, 17, 20), in order to identify the biological pathways that were significantly affected by differential expression. Our analysis revealed 19 pathways that were identified using the reactome pathway classification analysis with an FDR of <0.05 (Figure 3). All of the pathways were immune-related, with 166 out of the 824 DEGs containing multiple DEGs redundantly involved in different immunological pathways. Among these pathways, 15 contained 105 DEGs, and were statistically significant in terms of the differential mean expression between rNJ0612NME6- and the rNJ0612NME6-M51R-infected samples, as determined by unpaired t-tests (p < 0.05) (Figure 3, highlighted in grey). Fourteen pathways were associated with specific immune responses. Among them, the immune cytokine signaling pathway was found to be the most significantly affected, followed by eight pathways involved in IFN-related responses including activation, signaling, induction, and antiviral mechanisms (Figure 4). IFN-related pathways comprised a total of 56 genes accounting for 53% of the DEGs. The other five pathways were associated with MHC antigen processing and presentation or regulation of cell death (Figure 4). Interestingly, the Euler diagram showed 18 genes that were differentially upregulated among the different biological pathways identified in this study. Of relevance, the intracellular nucleic acid sensors DDX58 and IFIH1 (also known as RIG-I and MDA5, respectively) may play a central role in mediating the induction of type I IFN (alpha/beta) pathways, and consequently the up-regulation of other biological pathways (Figure 5). Our analysis suggests that the activation of viral RNA sensors during VSV infection may be a critical step in the control of this virus in a porcine model. However, given the functional redundancy among sensors, adaptors, and effectors of the IFN signaling pathway, defining a single functional network of genes responding to VSV infection may be challenging. A more detailed explanation about the multiple DEGs involved in different pathways identified in these analyses is presented in the following sections.
Figure 3.
Reactome pathway analysis. Reactome pathway analysis was conducted on a total 824 genes using the DAVID functional annotation tool. Selection of pathways was carried out based on FDR (p ≤ 0.05) criteria. Pathways highlighted in grey represent those with a statistically significant differential gene expression (p < 0.05) between rNJ0612NME6 and rNJ0612NME6-M51R. Details about the levels of expression of specific genes within the relevant pathways are presented in Figure 4.
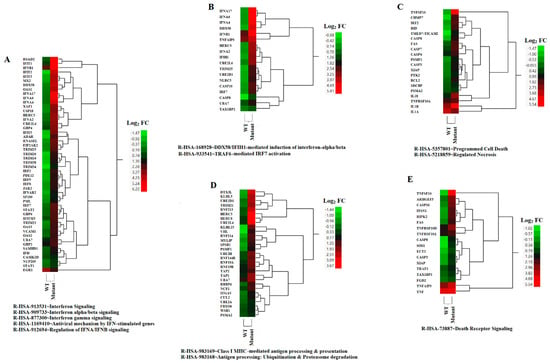
Figure 4.
HCA of relevant biological pathways differentially expressed due to infection by rNJ0612NME6 (WT) or rNJ0612NME6-M51R (mutant). Differential levels of expressions within genes associated with the identified biological/cellular pathways are represented (A–E). Values are expressed in terms of fold change (FC) Log2. The spectrum of colors ranges from green to red and reflect down- and up-regulation, respectively.
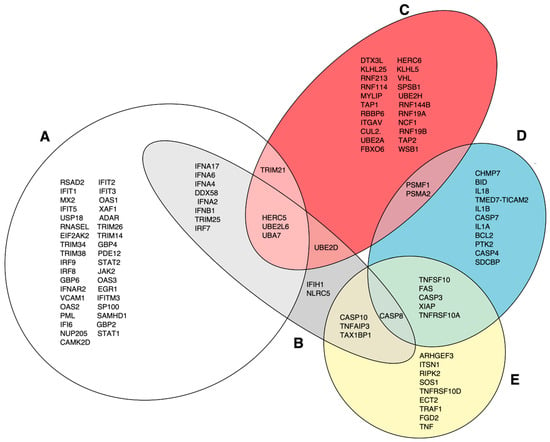
Figure 5.
Euler diagram analysis assessing the relationship among different pathways identified in this study. (A) represents genes in the following pathways: IFN signaling, IFN alpha/beta signaling, IFN gamma signaling, antiviral mechanisms by IFN-stimulated genes (ISGs), and regulation of IFNA/IFNB signaling. (B) DDX58/IFIH1-mediated induction of IFN alpha/beta. (C) Class I MHC-mediated antigen and presentation and antigen processing: ubiquitination and proteosome degradation. (D) Programed cell death and regulated necrosis. (E) Death receptor signaling.
3.3. DEGs Associated with IFN Responses
Macrophages infected with the M51R mutant virus expressed significantly higher levels of type I (IFNA, IFNB, IFND, and IFNW) and type III (IFNL) IFNs, but not type II IFN (IFNG) (Figure 6A). Among these IFNs, IFNA genes (including 2, 4, 6, and 17) were the most differentially expressed, followed by IFNL and IFND, where increased expression was observed in macrophages infected with the mutant virus compared to the WT virus-infected cells. The mutant-infected macrophages also expressed significantly higher levels of three IFN receptors (IFNAR1, IFNAR2, and IFNGR1), one IFN signaling transducer (JAK2), and three transcription factors (IRF9, STAT1, and STAT2) compared to the WT-infected cells (Figure 6B). These results suggest that the mutant-infected macrophages not only transcribed more type I and III IFNs, but also transduced IFN signals better than the WT-infected macrophages. This idea was strongly supported by the significantly up-regulated expression of many IFN-stimulated genes (ISGs) in the mutant-infected macrophages (Figure 4A). The results indicate that the M51R mutation in the M protein reduced the VSV’s ability to suppress IFN production and signaling. To corroborate with the results obtained by microarray analysis, we conducted type I IFN ELISA and showed that the type I IFN concentration was 3.77 times higher in the supernatants from cell culture infected with the mutant virus (1478.53 ± 86.89 pg/mL vs. 391.64 ± 73.62 pg/mL) than in those from WT virus infection.
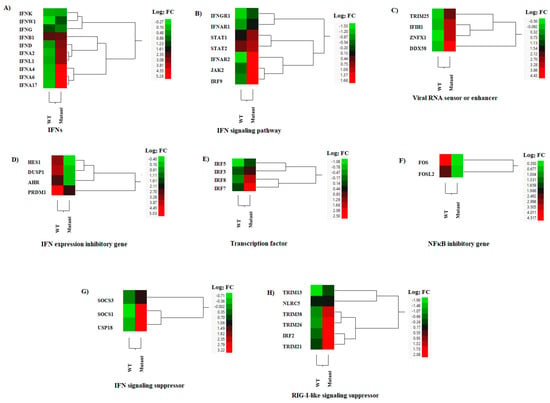
Figure 6.
Hierarchical cluster analysis (HCA) showing the differential gene expressions (DGEs) between rNJ0612NME6 (WT) or rNJ0612NME6-M51R (mutant) of genes associated with interferon (IFN) responses. These analysis included genes associated with; (A) Interferons (IFNs), (B) IFN signaling pathway, (C) Viral RNA sensing and enhancement of the IFN response, (D) Inhibition of IFN expression, (E) Transcription factors, (F) NFκΒ inhibition (G) IFN signaling suppression, and (H) Suppression of RIG-I-like sensors. HCA employs a color scheme to represent the extent of gene expression, with gradients ranging from green (suppressed or decreased) to red (increased). More information about the DGEs of these genes can be found in Supplementary Tables S1–S3.
Additionally, the expressions of three cytosolic pattern recognition receptors, IFIH1, RIG-I, and ZNFX1 [36,37], and an enhancer (TRIM25) [38], were significantly up-regulated in mutant-infected macrophages when compared with those infected with the WT virus (Figure 6C). In contrast, WT infection induced significantly higher expressions of genes that negatively regulate type I IFN expression, such as AHR [39], DUSP1 [40], HES1 [41], and PRDM1 [42], compared to the mutant-infected cells (Figure 6D). The expressions of these three genes were comparable between the mock-infected and mutant-infected macrophages (Supplementary File S2), indicating that the observed differences were caused by the WT infection. IRF3 and IRF7 are transcription factors that induce IFNB and IFNA transcription, respectively [43,44] (Figure 6E). FOS and FOSL2 are anti-inflammatory transcription factors that suppress NFκB activity [45], and may also suppress IFN expression. These genes were significantly down-regulated in the mutant virus-infected macrophages compared to the WT-infected cells (Figure 6F). Differential expression was observed for IRF7 but not for IRF3 (Figure 6E), which may explain the observed increased differential expression of IFNA genes compared to that of IFNB between the mutant- and WT-infected macrophages.
We also identified several DEGs that exhibit suppressive effects on IFN or RIG-I-like signaling in macrophages infected with the M51R mutant virus. These DEGs included three genes (SOCS1, SOCS3, and USP18) [46] (Figure 6G) involved in suppressing IFN signaling, and six genes (IRF2, NLRC5, TRIM13, TRIM 21, TRIM26, and TRIM38) [46] involved in suppressing RIG-I-like signaling, which were significantly up-regulated (Figure 6H).
3.4. DEGs of Interleukins, Chemokines, and Their Receptors
Several cytokines were up-regulated during the infection with both viruses (Figure 7A). Interestingly, ILIRN, known for its role to modulate inflammatory functions of cytokines including those stimulated by IL1A and IL1B [47], correlated with the clinical outcome linked to reduce disease severity in animals inoculated with this mutant virus. Furthermore, this suggests that the mutant virus may not induce an increase in body temperature, as was observed in the WT infection phenotype. Moreover, we found a reduction in CSF2 expression in the mutant-infected cells compared to WT-infected cells (Figure 7A). CSF2 is known to act as an anti-inflammatory/regulatory cytokine, promoting tolerogenic dendritic cell differentiation to enhance the numbers and function of regulatory T-cells [48]. The expression of IL15 (NK and cytotoxic T-cell-activating cytokine) [49] was not significantly different between both viruses. However, IL18 (Th1 response-enhancing cytokine) [50], IL27 (a T regulatory cell-promoting cytokine) [51], and TNFSF13B (a B-cell-activating TNF family member) [52] were significantly up-regulated in the mutant-infected macrophages compared to levels in WT-infected cells (Figure 7A). LIF, an anti-inflammatory cytokine [53] and a potent inhibitor of feed intake [54], was expressed at a significantly lower level in the mutant virus-infected cells compared to the WT-infected cells (Figure 7A). The expressions of IL1B (an endogenous pyrogen and proinflammatory cytokine) and IL10 (an immune suppressive cytokine) were not significantly different between the mutant- and WT-infected macrophages (Figure 7A).
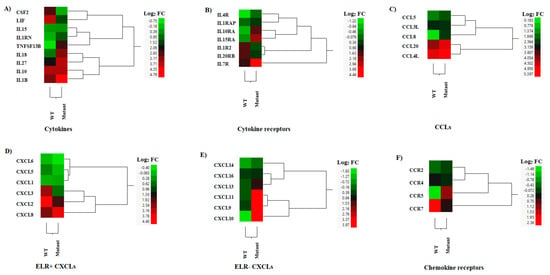
Figure 7.
Hierarchical cluster analysis (HCA) showing the differential gene expressions (DGEs) between rNJ0612NME6 (WT) or rNJ0612NME6-M51R (mutant) of genes associated with interleukins, chemokines, and their receptors. These analyses included genes implied with (A) Cytokine production, (B) Cytokine receptors, (C) C-C motif chemokine ligand genes (CCLs), (D) Pro-inflammatory cytokines containing a N-terminal glutamate, leucine, and arginine tripeptide motif (ELR+CXCLs), (E) ELR-CXCLs, and (F) Chemokine receptors. HCA employs a color scheme to represent the extent of gene expression, with gradients ranging from green (suppressed or decreased) to red (increased). More information about the DGEs of these genes can be found in Supplementary Table S5.
Our analysis identified eight interleukin receptors that were differentially expressed between the mutant- and WT-infected cells (Figure 7B). Notably, the expression of IL1R2 was down-regulated in mutant-infected cells compared to WT-infected cells, while IL1RAP expression was slightly up-regulated by the mutant infection. IL1R2 is a decoy receptor of IL1A and IL1B that interacts with IL1RAP to inhibit IL-1 signaling [55]. These results suggest that infection with the mutant virus may increase IL-1 signaling. The high-affinity receptor of IL15, IL15RA, can trans-present IL15 to potently stimulate NK and CD8+ Teff cells [56]. This IL15 receptor was significantly up-regulated by the mutant-infected cells. Among the other four up-regulated cytokine receptors, IL7R (stimulation of B-cell maturation, T and NK cell survival) and IL10RA (the receptor of the suppressive cytokine, IL-10) were up-regulated in the mutant-infected cells compared to the WT-infected cells (Figure 7B).
Different CCLs were expressed during the infection with both viruses (Figure 7C). Overall, higher levels of expression were seen in CCL20 and CCL4L, being slightly higher in the cells infected with the mutant virus (Figure 7C). Moreover, slightly higher levels of expression were observed in CCL5 during the infection with the mutant virus. Only the expression of CCL8 was significantly increased in comparison to the WT-infected cells (Figure 7C). CCL4 and CCL5 recruit macrophages and T cells [57]. CCL20 is a chemotactic factor for Th17 cells, and CCL8 promotes the Th2 response [57]. Regarding the overall expression of ELR+ (glutamic acid-leucine-arginine) CXCLs that recruit neutrophils and CD8+ effector T cells, the WT-infected cells expressed higher levels of all ELR+CXCLs than the mutant virus-infected cells (CXCL5, CXCL3, and CXCL2) (Figure 7D). The total expression of three ELR-CXCLs (CXCL9, CXCL10 and CXCL11) that recruit Th1, CD8+ T cells, and NK cells were up-regulated in mutant-infected cells compared to mock-infected cells (Figure 7E). Furthermore, CXCL13 was expressed slightly more in mutant-infected cells compared to WT-infected cells (Figure 7E).
The expressions of CCR5 (the receptor of CCL3, CCL4, and CCL5) and CCRL2 (expressed on neutrophils and primary monocytes) were significantly down-regulated by the WT infection (Figure 7F). Therefore, the expressions of CCR5 and CCLR2 were shown to be increased by the mutant-infected cells (Figure 7F). On the other hand, the expressions of CCR7 and CXCR4 were significantly induced by WT infection.
3.5. DEGs in MHC Antigen Presentation
Efficient MHC class I antigen presentation involves distinct biological pathways including ubiquitination, proteasomal degradation, and peptide loading assembly. In this study, the expression profiles of transcripts linked to ubiquitination (i.e., AREL1, RBBP6, UBA7, UBE2A, UBE2D1, UBE2H, UBE2B, UBE2K, UBE3A) and proteasomal degradation were up-regulated in cells infected with the VSV mutant when compared to mock infection (Figure 8A,B). Among the three peptide transporters identified, an increased expression was observed in TAP1 and TAP2 genes in cells infected with the mutant virus, while a higher expression was recorded for TAPBP during infection with the WT virus (Figure 8C).
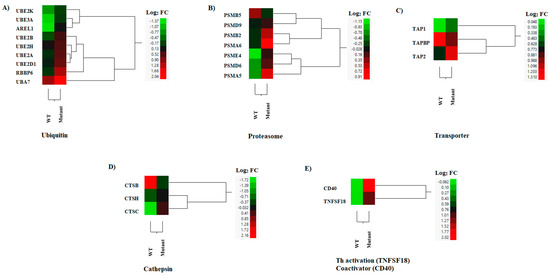
Figure 8.
Hierarchical cluster analysis (HCA) showing the differential gene expressions (DGEs) between rNJ0612NME6 (WT) or rNJ0612NME6-M51R (mutant) of genes associated with MHC antigen presentation. Class I antigen processing (A–C), class II antigen processing (D), and antigen presentation (E). HCA employs a color scheme to represent the extent of gene expression, with gradients ranging from green (suppressed or decreased) to red (increased). More information about the DGEs of these genes can be found in Supplementary Table S6.
In MHC class II antigen presentation, antigen acquisition and processing heavily rely on the endosomal/lysosomal compartments containing specific proteases such as cathepsins [58]. Two MHC class II-processing enzymes (CTSC and CTSH) were expressed at slightly higher levels, although only the increased expression of CTSC was statistically significant in the mutant-infected cells (Figure 8D). Conversely, higher expression levels were seen in the CTSB gene in cells infected with the WT virus (Figure 8D). Interestingly, TNFSF18 (a Th17-, Tfh-, and Th9 cell-activating TNF family member) [59] and CD40 (a T cell activation co-receptor of antigen presentation cells) [60] were significantly up-regulated by the mutant-infected macrophages when compared to WT-infected cells (Figure 8E). These results suggested that the mutant-infected cells could present MHC class I and II peptides more efficiently than the WT-infected cells.
3.6. DEGs in Apoptosis
VSV is well known for its capacity to induce apoptosis in infected cells [61]. Our results were consistent with previous publications indicating the increased and rapid ability of the VSV M51R mutation at the matrix protein to induce apoptosis in comparison with the WT protein [62]. Thirteen pro-apoptotic genes were expressed at higher levels by mutant-infected cells than the WT-infected cells (Figure 9A). The expressions of two death receptor (DR) ligands, TNFSF10 and TNFSF15, were higher in the mutant-infected cells (Figure 9B). In contrast, pro-apoptotic factor TNF was up-regulated in both infections, but not differentially expressed between the infections (Figure 9B). Three death receptors (TNFRSF1A, FAS, and TNFRSF10) were expressed at higher levels after mutant infection. Another TNF receptor, but not a death receptor, TNFRSF3/LTBR, was also up-regulated, whereas TNFRSF1B (an anti-apoptotic receptor) [63] was down-regulated. Three DR-signal-transducing genes (FADD, RIPK1, and TRAF2) were expressed at significantly higher levels in mutant-infected cells (Figure 9B), whereas the expressions of anti-apoptotic genes (REL and TNFRSF1B) in DR signaling pathways were down-regulated by mutant-infected cells compared to WT-infected cells (Figure 9B,C). These results suggest that macrophages infected with the mutant virus were able to express death receptor ligands at higher levels, and may be more sensitive to DR ligand-induced cell death or apoptosis.
Figure 9.
Hierarchical cluster analysis (HCA) showing the differential gene expressions (DGEs) between rNJ0612NME6 (WT) or rNJ0612NME6-M51R (mutant) of genes associated with apoptosis. These analyses included genes associated with; (A) Apoptosis, (B) Death receptor signaling and (C) Anti-apoptosis. HCA employs a color scheme to represent the extent of gene expression, with gradients ranging from green (suppressed or decreased) to red (increased). More information about the DGEs of these genes can be found in Supplementary Table S7.
3.7. Potential Relevance of the DGEs Observed in This Model
The effects of the M51R mutation in the VSV matrix protein inferred from differential gene expression are summarized in Table 1 The attenuation of M51R mutation in pigs is likely mediated by the up-regulated expressions of mainly type I and III IFN, IL1RN, and TNFSF10, and the down-regulated expression of LIF in the non-infected tissues. The up-regulated expressions of ELR+ CXCLs, IL1R2, death receptor, and their ligands could play additional roles in the control of inflammation and VSV infection of infected tissues. The genes induced by the mutant virus may also enhance MHC antigen processing and presentation as well as the immune response to induce early onset of protection via the innate antiviral activity of IFN, suggesting that the mutant virus could serve as a vector vaccine candidate. Additionally, the M51R mutation may also improve VSV oncolytic activity (Table 1).

Table 1.
The inferred effects of M51R mutation in VSV matrix protein on virus attenuation, vaccine vector, and cancer therapy of VSV based on genes differentially expressed between mutant- and WT-infected cells or genes induced by the mutant infection. Arrows ↑ and ↓ represent increased and decreased expression of specific genes respectively.
4. Discussion
The identification of molecular mechanisms associated with the virulence of VSV is critical to understand its pathogenesis in livestock. Currently, most of the knowledge in terms of its pathogeny comes from VSV mice models where macrophages, especially those residing in the lymph nodes, act as key players in the immune response against VSV [64,65,66,67,68]. In this regard, we used primary cultures of swine macrophages to evaluate immunological responses during VSV infection. In terms of host gene expression, VSV M protein inhibits the transcription of target genes [69] and nucleocytoplasmic transport [70]. Human IFNB gene is one of the genes that is targeted by the M protein [25]. The attenuation by mutations in the M protein have been well-characterized in mice [71]. We created the recombinant VSV with a similar mutation (rNJ0612NME6-M51R) as an experimental model for study in pigs. The rNJ0612NME6-M51R mutation was grown in ex vivo swine macrophages and displayed attenuated phenotypes in pigs [23].
Although rNJ0612NME6-M51R can induce large amounts of type I IFN in other primary cultures such as fetal kidney porcine cells (a mix of fibroblasts and epithelial cells), the growth of this virus was not compromised when compared with the parental strain. This observation is consistent with the ability of rNJ0612NME6-M51R to cause epithelial lesions upon intradermal inoculation in the snouts of pigs [23], highlighting the significance of macrophages in the pathogenesis of VSV in swine. In this study, we contrasted the genetic expression profiles between primary swine macrophages infected with the epidemic and highly virulent strains of VSNJV rNJ0612NME6 and the attenuated M51R mutant, in order to understand the mechanisms of attenuation.
We recently reported on genes induced by primary swine macrophages infected with the field isolate to infer VSV molecular pathogenesis [27]. In this study, our biological pathway analyses of DEGs identified several pathways associated with the type I IFN response (including activation, signaling, induction, and antiviral mechanisms), which was consistent with previous in vitro and in vivo experiments that demonstrated the susceptibility of VSV to the action of type I IFN [22,72,73,74]. Our findings indicate that the expressions of type I IFNs were significantly elevated in the mutant-infected cells compared to those in the WT-infected cells, confirming our expectations. Interestingly, we also observed a significant increase in the expressions of type III IFNs (IFNL), which has not been previously reported. These observations highlight the importance of further investigating the role of type III IFNs in the antiviral response to VSV infection.
In this study, the expression of IFNB in both mutant- and WT-infected cells was up-regulated. The studies of IFNB gene promoter have shown that activated ATF2-JUN, IRF3, and NFκB transcription factors induced IFNB expression [75]. These transcription factors were not differentially expressed between the M51R mutant and WT infection in this study. Instead, four IFN expression inhibitory genes (AHR, DUSP1, HES1, and PRDM1) were significantly up-regulated by WT infection. AHR down-regulates the IFNB production via up-regulating the expression of TIPARP to inhibit the activity of TBK1, a key signal transducer of IRF3 and NFκB signaling [39]. DUSP1 and HES1 are the inhibitors of pattern recognition receptor signaling pathways. DUSP1 is a negative regulator of mitogen-activated protein kinases (MAPK) that can inhibit IFNB expression by macrophages [40]. HES1 suppresses IFNB expression via attenuating signaling of poly(I:C), a synthetic mimic of viral double-stranded RNA, by inhibition of an adaptor molecule, WDFY1 [41]. PRDM1 is a transcription repressor that binds to the IFNB gene promoter [41]. Additionally, two other genes (FOS and FOSL2) were significantly up-regulated in WT-infected cells, but not in mutant-infected cells. These genes are anti-inflammatory-related transcription factors that suppress NFκB activity [45]. FOS and JUN form heterodimers called AP-1 [39], and a high expression of FOS could compete with ATF2 for dimerization, thereby suppressing IFNB transcription. The up-regulated expressions of these six genes (AHR, DUSP1, HES1, PRDM1, FOS, and FOSL2) in the WT- but not the mutant-infected cells suggest their role in suppressing IFNB expression by the M protein. In fact, it has been previously reported that the VSV M protein inhibits NFκB activation [76]. Our results suggest more detailed mechanisms of the M protein in suppressing IFNB expression.
IRF7 is the critical transcription factor of type I IFN expression induction, and the constitutive and induced expression of IRF7 in macrophages is primarily controlled by IFNB [44]. IRF7 expression was significantly up-regulated by the M51R mutant infection compared to the mock infection, whereas its expression between WT and the mock infection was comparable. Therefore, the suppressed production of type I IFNs by the VSV M protein was most likely mediated by suppression of IFNB expression via up-regulation of these six IFN inhibitory genes, including FOS and FOSL2. Additionally, our results revealed that the WT virus was capable of suppressing type I IFN signaling through down-regulation of IFNR2, IRF9, and STAT2 expression, which could subsequently limit the downstream effects of IFNB in inducing the expression of other type I IFNs. These differentially expressed genes provide insight into the mechanisms underlying the suppression of type I and possible type III IFN responses by WT-infected cells.
Given the potent antiviral activity of type I and III IFNs and our results, it is likely that the attenuation of VSV M51R in pigs is due to the reduction in the suppressive effects of the VSV M protein on IFNB expression and signaling. The expressions of IL1RN and TNFSF10 in mutant-infected cells were 22.7- and 80.5-fold higher, respectively, than those in WT-infected cells. IL1RN is the antagonist of IL-1 cytokines (potent pyrogens) [77]. The high induction of IL1RN in mutant-infected but not in WT-infected cells could be the main contribution to the lack of fever observed in the mutant-infected pigs [23]. Since IFN signaling induces IL1RN expression [78,79], the up-regulated IL1RN was likely due to increased expression of type I IFN. Likewise, human TNFSF10 induces cell death of virus-infected cells but not of uninfected cells, and its expression could also be induced by type I IFN signaling [80]. Therefore, up-regulated TNFSF10 in the mutant infection could only be another factor contributing to the attenuation.
Post-translational modifications (i.e., phosphorylation, ubiquitination, ISGylation) activate transcription factors that modulate IFN transcription. It is worth noting that substantial up-regulation of transcripts (UBA7, UBE2L6, HERC5) encoding cellular functions involved in conjugating the ubiquitin-like molecule ISG15 (ISGylation) to different protein substrates were featured in the Euler diagram. These results suggest that infection with the VSV mutant virus may promote cellular ISGylation to modulate immune responses in macrophages. In support of these observations, recent studies have demonstrated that protein ISGylation may regulate the inflammatory responses of macrophages during virus infection [81]. Further studies measuring total ISGylation are warranted to examine this role in the pathogenesis of VSV.
The activation of apoptosis by rNJ0612NME6-M51R using the extrinsic pathway was highly supported by our findings, identifying the relevance of the death receptor signaling pathway during our analysis according to [82]. TNF represents a key molecule to initiate the activation of the extrinsic pathway [83]. Interestingly, TNF was up-regulated by both the mutant and WT viruses at nearly the same level, by approximately 25-fold. TNF has been described for its essential role in promoting the survival of CD169+ macrophages during the infection of VSV in mice [84], suggesting that the ability of both viruses to induce TNF expression in macrophages was not responsible for the phenotypic differences seen in both viruses in pigs.
The advances in the understanding of the biology of VSV have led to the development of VSV recombinant viruses with oncolytic properties [8], and as a virally vectored vaccine platform [9,10]. In this sense, our results suggest that the M51R VSV mutant may be a better viral vaccine vector than VSV without the M51R mutation, based on the genes induced after infection; a summary of the differential gene expressions between mutant and WT infection is listed in Table 1. Firstly, the mutant infection induced production of a large amount of type I and III interferons, which can translate into early innate immune protection. Secondly, the ubiquitin–proteasome system is critical for MHC antigen processing [85]. Our results showed that the expressions of several ubiquitins were induced by the mutant infection, and the overall expressions of ubiquitin and proteasome protease genes were expressed at higher levels in the mutant-infected macrophages than in the WT-infected cells. Thirdly, several up-regulated genes, including IL18 [50], CCL4, CCL5, CCL8, CCL20 [57], CD40 [60], TNFSF13B [52], and TNFSF18 [59], may enhance MHC antigen presentation or the immune response except IL27, which mainly promotes the Treg response [51]. Likewise, the M51R VSV mutant may be a better oncolytic virus than the VSV without the M51R mutation. Type I IFN [86] and TNFSF10 [80] have been reported to have antitumor activity, and their expressions were significantly higher in the mutant-infected cells than the WT-infected cells.
5. Conclusions
Collectively, our results highlight the potential relevance of type I IFN-involved pathways in the pathogenesis of VSV in pigs. This assertion is supported by our previous experiment utilizing VSV-IFN beta-NIS, which demonstrated the impact of type IFN on the pathogenesis in pigs [22]. Our analysis suggested the importance of the DDX58/IFIHI-mediated induction of the interferon alpha/beta and TRAF6-mediated IRF7 activation pathways to explain the differences in pathogenesis between rNJ0612NME6 and rNJ0612NME6-M51R. In this sense, while previous studies have evaluated the induction of type I IFN by various isolates in primary cultures of chicken fibroblasts [87], it is important to emphasize the significance of conducting these experiments using a more physiologically relevant cell line to study the pathogenesis of VSV. Building upon the results presented in this study, we are currently investigating the capacity of an extensive and genetically diverse collection of VSV isolates, representative of multiple geographic regions in the Americas, in order to induce type I IFN in porcine macrophages.
The main goal of this study was to utilize rNJ0612NME6-M51R as proof-of-concept to unravel the molecular basis of VSV pathogenesis in pigs, with the ultimate aim of extending this understanding to other susceptible livestock species. Currently, there is limited characterization of VSV strains in terms of their virulence levels in pigs and other livestock susceptible species. Nevertheless, our findings demonstrate important parallels between the pathogenicity induced by rNJ0612NME6-M51R and some field isolates of VSIV in pigs [18,19,23,24]. Therefore, our study offers a platform for future research aimed at understanding the variations in virulence between VSNJV and VSIV. Finally, our study identified multiple genes implicated in the repression of type I IFN, shedding light on potential avenues for investigating alternative mechanisms of cytokine control. It is important to note that this study primarily serves as a descriptive analysis, and as such, additional experiments are warranted to validate and explore the hypothesis generated herein. Such investigations will contribute to a more comprehensive understanding of the complex interplay between gene regulation and type I IFN responses during VSV infection.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/pathogens12070896/s1, Figure S1: qPCR validation of some relevant genes; Table S1: Expression levels (EXP), false discovery rates (FDR) and fold differences (mutant vs. WT, M/W and mutant vs. mock infection, (M/N) of interferon related genes differentially expressed by infected macrophages; Table S2. Expression levels (EXP), false discovery rates (FDR) and fold differences (mutant vs. wildtype, M/W and mutant vs. mock infection, M/N) of differentially expressed genes in cytosolic pattern recognition receptor signaling pathways or with inhibitory effect on IFN expression; Table S3. Expression levels (EXP), false discovery rates (FDR) and fold differences (mutant vs. WT, M/W and mutant vs. mock infection, M/N) of differentially expressed genes with suppressive effects on IFN and RIG-I-like signaling; Table S4. Expression levels (EXP), false discovery rates (FDR) and fold differences (mutant vs. WT, M/W and mutant vs. mock infection, M/N) of cytokine, chemokine and the receptor genes differentially expressed by infected macrophages; Table S5. Expression levels (EXP), false discovery rates (FDR) and fold differences (mutant vs. WT, M/W and mutant vs. mock infection, M/N) of chemokine and receptor genes differentially expressed by infected macrophages; Table S6. Expression levels (EXP), false discovery rates (FDR) and fold differences (mutant vs. WT, M/W and mutant vs. mock infection, M/N) of chemokine and receptor genes differentially expressed by infected macrophages; Table S7. Expression levels (EXP), false discovery rates (FDR) and fold differences (mutant vs. WT, M/W and mutant vs. mock infection, M/N) of apoptosis related genes differentially expressed by infected macrophages.
Author Contributions
Conceptualization, L.V.-S., L.L.R. and J.J.Z.; methodology, L.V.-S. and J.J.Z.; validation, L.V.-S., S.C. and J.J.Z.; formal analysis, L.V.-S., F.V., S.Z. and J.J.Z.; investigation, L.V.-S., G.N.M., F.V., S.Z., S.C. and J.J.Z.; resources, L.L.R. and J.J.Z.; data curation, L.V.-S. and J.J.Z.; writing—original draft preparation, L.V.-S., G.N.M. and J.J.Z.; writing—review and editing, L.V.-S., G.N.M., F.V., S.C., J.J.Z. and L.L.R.; supervision, L.V.-S., J.J.Z. and L.L.R.; project administration, L.V.-S., J.J.Z. and L.L.R.; funding acquisition, L.L.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was performed under USDA Research Service CRIS Project 3022-32000-062-000-D.
Institutional Review Board Statement
All animal research was approved by The PIADC Institutional Animal Care and Use Committee of the US Departments of Agriculture and Homeland Security (Protocol Number: 247.01-21-R).
Informed Consent Statement
Not applicable.
Data Availability Statement
Raw data and the statistical results of the microarray analysis conducted in this study are available at the NCBI Gene Expression Omnibus (GEO) database on the following accession: ID GSE225798.
Acknowledgments
We would like to thank Manuel V. Borca and Elizabeth Ramirez-Medina from the Plum Island Animal Disease Center for kindly providing the cultures of primary swine macrophages used in this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rodriguez, L.L. Emergence and re-emergence of vesicular stomatitis in the United States. Virus Res. 2002, 85, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Velazquez-Salinas, L.; Zarate, S.; Eschbaumer, M.; Lobo, F.P.; Gladue, D.P.; Arzt, J.; Novella, I.S.; Rodriguez, L.L. Selective Factors Associated with the Evolution of Codon Usage in Natural Populations of Arboviruses. PLoS ONE 2016, 11, e0159943. [Google Scholar] [CrossRef] [PubMed]
- Dietzgen, R.G. Morphology, Genome Organization, Transcription and Replication of Rhabdoviruses; Kuzmin, R.G.D.a.I.V., Ed.; Caister Academic Press: Norfolk, UK, 2012; pp. 5–11. [Google Scholar]
- Pelzel-McCluskey, A.; Christensen, B.; Humphreys, J.; Bertram, M.; Keener, R.; Ewing, R.; Cohnstaedt, L.W.; Tell, R.; Peters, D.P.C.; Rodriguez, L. Review of Vesicular Stomatitis in the United States with Focus on 2019 and 2020 Outbreaks. Pathogens 2021, 10, 993. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.; McKenzie, M.O.; Puckett, S.; Hojnacki, M.; Poliquin, L.; Lyles, D.S. Ability of the matrix protein of vesicular stomatitis virus to suppress beta interferon gene expression is genetically correlated with the inhibition of host RNA and protein synthesis. J. Virol. 2003, 77, 4646–4657. [Google Scholar] [CrossRef]
- Hastie, E.; Cataldi, M.; Marriott, I.; Grdzelishvili, V.Z. Understanding and altering cell tropism of vesicular stomatitis virus. Virus Res. 2013, 176, 16–32. [Google Scholar] [CrossRef]
- Rajani, K.R.; Kneller, E.L.P.; McKenzie, M.O.; Horita, D.A.; Chou, J.W.; Lyles, D.S. Complexes of vesicular stomatitis virus matrix protein with host Rae1 and Nup98 involved in inhibition of host transcription. PLOS Pathog. 2012, 8, e1002929. [Google Scholar] [CrossRef]
- Leblanc, A.K.; Naik, S.; Galyon, G.D.; Jenks, N.; Steele, M.B.; Peng, K.-W.; Federspiel, M.J.; Donnell, R.L.; Russell, S.J. Safety studies on intravenous administration of oncolytic recombinant vesicular stomatitis virus in purpose-bred beagle dogs. Hum. Gene Ther. Clin. Dev. 2013, 24, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Matassov, D.; Marzi, A.; Latham, T.; Xu, R.; Ota-Setlik, A.; Feldmann, F.; Geisbert, J.B.; Mire, C.E.; Hamm, S.; Nowak, B.; et al. Vaccination with a Highly Attenuated Recombinant Vesicular Stomatitis Virus Vector Protects Against Challenge with a Lethal Dose of Ebola Virus. J. Infect. Dis. 2015, 212 (Suppl. S2), S443–S451. [Google Scholar] [CrossRef]
- Zhang, M.; Ge, J.; Li, X.; Chen, W.; Wang, X.; Wen, Z.; Bu, Z. Protective efficacy of a recombinant Newcastle disease virus expressing glycoprotein of vesicular stomatitis virus in mice. Virol. J. 2016, 13, 31. [Google Scholar] [CrossRef]
- O’donnell, V.K.; Pauszek, S.J.; Xu, L.; Moran, K.; Vierra, D.; Boston, T.; Dodd, K.A.; Faburay, B.; Barrette, R.W. Genome Sequences of Vesicular Stomatitis Indiana Viruses from the 2019 Outbreak in the Southwest United States. Microbiol. Resour. Announc. 2020, 9, e00894-20. [Google Scholar] [CrossRef]
- Rainwater-Lovett, K.; Pauszek, S.J.; Kelley, W.N.; Rodriguez, L.L. Molecular epidemiology of vesicular stomatitis New Jersey virus from the 2004–2005 US outbreak indicates a common origin with Mexican strains. J. Gen. Virol. 2007, 88, 2042–2051. [Google Scholar] [CrossRef]
- Rodriguez, L.L.; Bunch, T.A.; Fraire, M.; Llewellyn, Z.N. Re-emergence of vesicular stomatitis in the western United States is associated with distinct viral genetic lineages. Virology 2000, 271, 171–181. [Google Scholar] [CrossRef]
- Velazquez-Salinas, L.; Pauszek, S.J.; Zarate, S.; Basurto-Alcantara, F.J.; Verdugo-Rodriguez, A.; Perez, A.M.; Rodriguez, L.L. Phylogeographic characteristics of vesicular stomatitis New Jersey viruses circulating in Mexico from 2005 to 2011 and their relationship to epidemics in the United States. Virology 2014, 449, 17–24. [Google Scholar] [CrossRef]
- Mead, D.G.; Lovett, K.R.; Murphy, M.D.; Pauszek, S.J.; Smoliga, G.; Gray, E.W.; Noblet, R.; Overmyer, J.; Rodriguez, L.L. Experimental transmission of vesicular stomatitis New Jersey virus from Simulium vittatum to cattle: Clinical outcome is in-fluenced by site of insect feeding. J. Med. Entomol. 2009, 46, 866–872. [Google Scholar] [CrossRef]
- Velazquez-Salinas, L.; Pauszek, S.J.; Stenfeldt, C.; O’hearn, E.S.; Pacheco, J.; Borca, M.V.; Verdugo-Rodriguez, A.; Arzt, J.; Rodriguez, L.L. Increased Virulence of an Epidemic Strain of Vesicular Stomatitis Virus Is Associated With Interference of the Innate Response in Pigs. Front. Microbiol. 2018, 9, 1891. [Google Scholar] [CrossRef]
- Green, S.L. Vesicular Stomatitis in the Horse. Veter-Clin. North Am. Equine Pract. 1993, 9, 349–353. [Google Scholar] [CrossRef]
- Martinez, I.; Rodriguez, L.L.; Jimenez, C.; Pauszek, S.J.; Wertz, G.W. Vesicular stomatitis virus glycoprotein is a deter-minant of pathogenesis in swine, a natural host. J. Virol. 2003, 77, 8039–8047. [Google Scholar] [CrossRef]
- Morozov, I.; Davis, A.S.; Ellsworth, S.; Trujillo, J.D.; McDowell, C.; Shivanna, V.; Dasen, E.J.; Nichols, R.; Martin, B.K.; Monath, T.P.; et al. Comparative evaluation of pathogenicity of three isolates of vesicular stomatitis virus (Indiana serotype) in pigs. J. Gen. Virol. 2019, 100, 1478–1490. [Google Scholar] [CrossRef] [PubMed]
- Stallknecht, D.E.; Howerth, E.W.; Reeves, C.L.; Seal, B.S. Potential for contact and mechanical vector transmission of vesicular stomatitis virus New Jersey in pigs. Am. J. Veter-Res. 1999, 60, 43–48. [Google Scholar]
- Stallknecht, D.E.; Perzak, D.E.; Bauer, L.D.; Murphy, M.D.; Howerth, E.W. Contact transmission of vesicular stomatitis virus New Jersey in pigs. Am. J. Veter-Res. 2001, 62, 516–520. [Google Scholar] [CrossRef] [PubMed]
- Velazquez-Salinas, L.; Naik, S.; Pauszek, S.J.; Peng, K.-W.; Russell, S.J.; Rodriguez, L.L. Oncolytic Recombinant Vesicular Stomatitis Virus (VSV) Is Nonpathogenic and Nontransmissible in Pigs, a Natural Host of VSV. Hum. Gene Ther. Clin. Dev. 2017, 28, 108–115. [Google Scholar] [CrossRef]
- Velazquez-Salinas, L.; Pauszek, S.J.; Holinka, L.G.; Gladue, D.P.; Rekant, S.I.; Bishop, E.A.; Stenfeldt, C.; Verdugo-Rodriguez, A.; Borca, M.V.; Arzt, J.; et al. A Single Amino Acid Substitution in the Matrix Protein (M51R) of Vesicular Stomatitis New Jersey Virus Impairs Replication in Cultured Porcine Macrophages and Results in Significant Attenuation in Pigs. Front. Microbiol. 2020, 11, 1123. [Google Scholar] [CrossRef]
- Stallknecht, D.E.; Greer, J.B.; Murphy, M.D.; Mead, D.G.; Howerth, E.W. Effect of strain and serotype of vesicular stomatitis virus on viral shedding, vesicular lesion development, and contact transmission in pigs. Am. J. Veter-Res. 2004, 65, 1233–1239. [Google Scholar] [CrossRef]
- Ferran, M.C.; Lucas-Lenard, J.M. The vesicular stomatitis virus matrix protein inhibits transcription from the human beta interferon promoter. J. Virol. 1997, 71, 371–377. [Google Scholar] [CrossRef] [PubMed]
- Stojdl, D.F.; Lichty, B.D.; Tenoever, B.R.; Paterson, J.M.; Power, A.T.; Knowles, S.; Marius, R.; Reynard, J.; Poliquin, L.; Atkins, H.; et al. VSV strains with defects in their ability to shutdown innate immunity are potent systemic anti-cancer agents. Cancer Cell 2003, 4, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Velazquez-Salinas, L.; Canter, J.A.; Zhu, J.J.; Rodriguez, L.L. Molecular Pathogenesis and Immune Evasion of Vesicular Stomatitis New Jersey Virus Inferred from Genes Expression Changes in Infected Porcine Macrophages. Pathogens 2021, 10, 1134. [Google Scholar] [CrossRef] [PubMed]
- Borca, M.; Berggren, K.; Ramirez-Medina, E.; Vuono, E.; Gladue, D. CRISPR/Cas Gene Editing of a Large DNA Virus: African Swine Fever Virus. Bio-Protocol 2018, 8, e2978. [Google Scholar] [CrossRef] [PubMed]
- Velazquez-Salinas, L.; Pauszek, S.J.; Barrera, J.; Clark, B.; Borca, M.V.; Verdugo-Rodriguez, A.; Stenfeldt, C.; Arzt, J.; Rodriguez, L.L. Validation of a site-specific recombination cloning technique for the rapid development of a full-length cDNA clone of a virulent field strain of vesicular stomatitis New Jersey virus. J. Virol. Methods 2019, 265, 113–116. [Google Scholar] [CrossRef]
- Zhu, J.J.; Ramanathan, P.; Bishop, E.A.; O’donnell, V.; Gladue, D.P.; Borca, M.V. Mechanisms of African swine fever virus pathogenesis and immune evasion inferred from gene expression changes in infected swine macrophages. PLoS ONE 2019, 14, e0223955. [Google Scholar] [CrossRef]
- Smyth, G.; Gentleman, R.; Carey, V.; Dudoit, S.; Irizarry, R.; Huber, W. Limma: Linear Models for Microarray Data, Bioinformatics and Computational Biology Solutions Using R and Bioconductor; Springer: New York, NY, USA, 2005; pp. 397–420. [Google Scholar]
- Huangda, W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: Paths toward the comprehensive func-tional analysis of large gene lists. Nucleic. Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef]
- Fabregat, A.; Sidiropoulos, K.; Viteri, G.; Forner, O.; Marin-Garcia, P.; Arnau, V.; D’eustachio, P.; Stein, L.; Hermjakob, H. Reactome pathway analysis: A high-performance in-memory approach. BMC Bioinform. 2017, 18, 142. [Google Scholar] [CrossRef] [PubMed]
- Kestler, H.A.; Müller, A.; Kraus, J.M.; Buchholz, M.; Gress, T.M.; Liu, H.; Kane, D.W.; Zeeberg, B.R.; Weinstein, J.N. VennMaster: Area-proportional Euler diagrams for functional GO analysis of microarrays. BMC Bioinform. 2008, 9, 67. [Google Scholar] [CrossRef]
- Wybrow, M.; Rodgers, P.; Dib, F.K. Euler diagrams drawn with ellipses area-proportionally (Edeap). BMC Bioinform. 2021, 22, 214. [Google Scholar] [CrossRef] [PubMed]
- Rehwinkel, J.; Gack, M.U. RIG-I-like receptors: Their regulation and roles in RNA sensing. Nat. Rev. Immunol. 2020, 20, 537–551. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yuan, S.; Jia, X.; Ge, Y.; Ling, T.; Nie, M.; Lan, X.; Chen, S.; Xu, A. Mitochondria-localised ZNFX1 functions as a dsRNA sensor to initiate antiviral responses through MAVS. Nature 2019, 21, 1346–1356. [Google Scholar] [CrossRef]
- Choudhury, N.R.; Heikel, G.; Michlewski, G. TRIM25 and its emerging RNA-binding roles in antiviral defense. Wiley Interdiscip. Rev. RNA 2020, 11, e1588. [Google Scholar] [CrossRef]
- Yamada, T.; Horimoto, H.; Kameyama, T.; Hayakawa, S.; Yamato, H.; Dazai, M.; Takada, A.; Kida, H.; Bott, D.; Zhou, A.C.; et al. Constitutive aryl hydrocarbon receptor signaling constrains type I interferon–mediated antiviral innate defense. Nat. Immunol. 2016, 17, 687–694. [Google Scholar] [CrossRef]
- McGuire, V.A.; Rosner, D.; Ananieva, O.; Ross, E.A.; Elcombe, S.E.; Naqvi, S.; van den Bosch, M.M.W.; Monk, C.E.; Ruiz-Zorrilla Diez, T.; Clark, A.R.; et al. Beta Interferon Production Is Regulated by p38 Mitogen-Activated Protein Kinase in Macrophages via both MSK1/2- and Tristetraprolin-Dependent Pathways. Mol. Cell Biol. 2017, 37, e00454-16. [Google Scholar] [CrossRef]
- Ning, F.; Li, X.; Yu, L.; Zhang, B.; Zhao, Y.; Liu, Y.; Zhao, B.; Shang, Y.; Hu, X. Hes1 attenuates type I IFN responses via VEGF-C and WDFY1. J. Exp. Med. 2019, 216, 1396–1410. [Google Scholar] [CrossRef]
- Keller, A.D.; Maniatis, T. Identification and characterization of a novel repressor of beta-interferon gene expression. Genes Dev. 1991, 5, 868–879. [Google Scholar] [CrossRef]
- Marié, I.; Durbin, J.E.; Levy, D.E. Differential viral induction of distinct interferon-alpha genes by positive feedback through interferon regulatory factor-7. EMBO J. 1998, 17, 6660–6669. [Google Scholar] [CrossRef] [PubMed]
- Sin, W.X.; Yeong, J.P.; Lim, T.J.F.; Su, I.H.; Connolly, J.E.; Chin, K.C. IRF-7 Mediates Type I IFN Responses in Endo-toxin-Challenged Mice. Front. Immunol. 2020, 11, 640. [Google Scholar] [CrossRef] [PubMed]
- Ray, N.; Kuwahara, M.; Takada, Y.; Maruyama, K.; Kawaguchi, T.; Tsubone, H.; Ishikawa, H.; Matsuo, K. c-Fos sup-presses systemic inflammatory response to endotoxin. Int. Immunol. 2006, 18, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Arimoto, K.-I.; Miyauchi, S.; Stoner, S.A.; Fan, J.-B.; Zhang, D.-E. Negative regulation of type I IFN signaling. J. Leukoc. Biol. 2018, 103, 1099–1116. [Google Scholar] [CrossRef]
- Weber, A.; Wasiliew, P.; Kracht, M. Interleukin-1 (IL-1) Pathway. Sci. Signal. 2010, 3, cm1. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, P.; Budnick, I.; Singh, M.; Thiruppathi, M.; Alharshawi, K.; Elshabrawy, H.; Holterman, M.J.; Prabhakar, B.S. Dual Role of GM-CSF as a Pro-Inflammatory and a Regulatory Cytokine: Implications for Immune Therapy. J. Interf. Cytokine Res. 2015, 35, 585–599. [Google Scholar] [CrossRef] [PubMed]
- Perera, P.-Y.; Lichy, J.H.; Waldmann, T.A.; Perera, L.P. The role of interleukin-15 in inflammation and immune responses to infection: Implications for its therapeutic use. Microbes Infect. 2012, 14, 247–261. [Google Scholar] [CrossRef]
- Nakanishi, K. Unique Action of Interleukin-18 on T Cells and Other Immune Cells. Front. Immunol. 2018, 9, 763. [Google Scholar] [CrossRef]
- Yoshida, H.; Hunter, C.A. The Immunobiology of Interleukin-27. Annu. Rev. Immunol. 2015, 33, 417–443. [Google Scholar] [CrossRef]
- Smulski, C.R.; Eibel, H. BAFF and BAFF-Receptor in B Cell Selection and Survival. Front. Immunol. 2018, 9, 2285. [Google Scholar] [CrossRef]
- Weber, M.A.; Schnyder-Candrian, S.; Schnyder, B.; Quesniaux, V.; Poli, V.; Stewart, C.L.; Ryffel, B. Endogenous leu-kemia inhibitory factor attenuates endotoxin response. Lab. Investig. 2005, 85, 276–284. [Google Scholar] [CrossRef]
- Daniel, J.A.; Whitlock, B.K.; Marks, D.L.; Gard, J.A.; Sartin, J.L. Leukemia inhibitory factor as a mediator of lipopoly-saccharide effects on appetite and selected hormones and metabolites. J. Anim. Sci. 2016, 94, 2789–2797. [Google Scholar] [CrossRef]
- Lang, D.; Knop, J.; Wesche, H.; Raffetseder, U.; Kurrle, R.; Boraschi, D.; Martin, M.U. The The type II IL-1 receptor interacts with the IL-1 receptor accessory protein: A novel mechanism of regulation of IL-1 responsiveness. J. Immunol. 1998, 161, 6871–6877. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Luan, L.; Patil, N.K.; Sherwood, E.R. Immunobiology of the IL-15/IL-15Ralpha complex as an antitumor and antiviral agent. Cytokine Growth Factor Rev. 2017, 38, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Griffith, J.W.; Sokol, C.L.; Luster, A.D. Chemokines and chemokine receptors: Positioning cells for host defense and immunity. Annu. Rev. Immunol. 2014, 32, 659–702. [Google Scholar] [CrossRef]
- Shi, G.-P.; Villadangos, J.A.; Dranoff, G.; Small, C.; Gu, L.; Haley, K.J.; Riese, R.; Ploegh, H.L.; Chapman, H.A. Cathepsin S required for normal MHC class II peptide loading and germinal center development. Immunity 1999, 10, 197–206. [Google Scholar] [CrossRef]
- Tian, J.; Zhang, B.; Rui, K.; Wang, S. The Role of GITR/GITRL Interaction in Autoimmune Diseases. Front. Immunol. 2020, 11, 588682. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, B.; Thomas, R. CD40 and dendritic cell function. Crit. Rev. Immunol. 2003, 23, 83–107. [Google Scholar] [CrossRef]
- Gadaleta, P.; Perfetti, X.; Mersich, S.; Coulombié, F. Early activation of the mitochondrial apoptotic pathway in Vesicular Stomatitis virus-infected cells. Virus Res. 2005, 109, 65–69. [Google Scholar] [CrossRef]
- Gaddy, D.F.; Lyles, D.S. Vesicular stomatitis viruses expressing wild-type or mutant M proteins activate apoptosis through distinct pathways. J. Virol. 2005, 79, 4170–4179. [Google Scholar] [CrossRef]
- Al-Lamki, R.S.; Mayadas, T.N. TNF receptors: Signaling pathways and contribution to renal dysfunction. Kidney Int. 2015, 87, 281–296. [Google Scholar] [CrossRef]
- Iannacone, M.; Moseman, E.A.; Tonti, E.; Bosurgi, L.; Junt, T.; Henrickson, S.E.; Whelan, S.P.; Guidotti, L.G.; von Andrian, U.H. Subcapsular sinus macrophages prevent CNS invasion on peripheral infection with a neurotropic virus. Nature 2010, 465, 1079–1083. [Google Scholar] [CrossRef]
- Junt, T.; Moseman, E.A.; Iannacone, M.; Massberg, S.; Lang, P.A.; Boes, M.; Fink, K.; Henrickson, S.E.; Shayakhmetov, D.M.; Di Paolo, N.C.; et al. Subcapsular sinus macrophages in lymph nodes clear lymph-borne viruses and present them to antiviral B cells. Nature 2007, 450, 110–114. [Google Scholar] [CrossRef]
- Louie, D.A.P.; Liao, S. Lymph Node Subcapsular Sinus Macrophages as the Frontline of Lymphatic Immune Defense. Front. Immunol. 2019, 10, 347. [Google Scholar] [CrossRef] [PubMed]
- Moseman, E.A.; Iannacone, M.; Bosurgi, L.; Tonti, E.; Chevrier, N.; Tumanov, A.; Fu, Y.-X.; Hacohen, N.; von Andrian, U.H. B cell maintenance of subcapsular sinus macrophages protects against a fatal viral infection independent of adaptive immunity. Immunity 2012, 36, 415–426. [Google Scholar] [CrossRef] [PubMed]
- Solmaz, G.; Puttur, F.; Francozo, M.; Lindenberg, M.; Guderian, M.; Swallow, M.; Duhan, V.; Khairnar, V.; Kalinke, U.; Ludewig, B.; et al. TLR7 Controls VSV Replication in CD169+ SCS Macrophages and Associated Viral Neuroinvasion. Front. Immunol. 2019, 10, 466. [Google Scholar] [CrossRef] [PubMed]
- Black, B.L.; Lyles, D.S. Vesicular stomatitis virus matrix protein inhibits host cell-directed transcription of target genes in vivo. J. Virol. 1992, 66, 4058–4064. [Google Scholar] [CrossRef]
- Petersen, J.M.; Her, L.-S.; Varvel, V.; Lund, E.; Dahlberg, J.E. The matrix protein of vesicular stomatitis virus inhibits nucleocytoplasmic transport when it is in the nucleus and associated with nuclear pore complexes. Mol. Cell. Biol. 2000, 20, 8590–8601. [Google Scholar] [CrossRef]
- Jayakar, H.R.; Whitt, M.A. Identification of two additional translation products from the matrix (M) gene that contribute to vesicular stomatitis virus cytopathology. J. Virol. 2002, 76, 8011–8018. [Google Scholar] [CrossRef] [PubMed]
- Belkowski, L.S.; Sen, G.C. Inhibition of vesicular stomatitis viral mRNA synthesis by interferons. J. Virol. 1987, 61, 653–660. [Google Scholar] [CrossRef]
- Kueck, T.; Bloyet, L.-M.; Cassella, E.; Zang, T.; Schmidt, F.; Brusic, V.; Tekes, G.; Pornillos, O.; Whelan, S.P.J.; Bieniasz, P.D. Vesicular Stomatitis Virus Transcription Is Inhibited by TRIM69 in the Interferon-Induced Antiviral State. J. Virol. 2019, 93, e01372-19. [Google Scholar] [CrossRef] [PubMed]
- Repik, P.; Flamand, A.; Bishop, D.H.L. Effect of interferon upon the primary and secondary transcription of vesicular stomatitis and influenza viruses. J. Virol. 1974, 14, 1169–1178. [Google Scholar] [CrossRef]
- Honda, K.; Takaoka, A.; Taniguchi, T. Type I interferon [corrected] gene induction by the interferon regulatory factor family of transcription factors. Immunity 2006, 25, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Varble, A.J.; Ried, C.D.; Hammond, W.J.; Marquis, K.A.; Woodruff, M.C.; Ferran, M.C. The vesicular stomatitis virus matrix protein inhibits NF-kappaB activation in mouse L929 cells. Virology 2016, 499, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Cartmell, T.; Luheshi, G.N.; Hopkins, S.J.; Rothwell, N.J.; Poole, S. Role of endogenous interleukin-1 receptor antagonist in regulating fever induced by localised inflammation in the rat. J. Physiol. 2001, 531, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Tilg, H.; Mier, J.W.; Vogel, W.; Aulitzky, W.E.; Wiedermann, C.J.; Vannier, E.; Huber, C.; Dinarello, C.A. Induction of circulating IL-1 receptor antagonist by IFN treatment. J. Immunol. 1993, 150, 4687–4692. [Google Scholar] [CrossRef]
- Liu, J.S.H.; Amaral, T.D.; Brosnan, C.F.; Lee, S.C. IFNs are critical regulators of IL-1 receptor antagonist and IL-1 expression in human microglia. J. Immunol. 1998, 161, 1989–1996. [Google Scholar] [CrossRef]
- Tecchio, C.; Huber, V.; Scapini, P.; Calzetti, F.; Margotto, D.; Todeschini, G.; Pilla, L.; Martinelli, G.; Pizzolo, G.; Rivoltini, L.; et al. IFNalpha-stimulated neutrophils and monocytes release a soluble form of TNF-related apopto-sis-inducing ligand (TRAIL/Apo-2 ligand) displaying apoptotic activity on leukemic cells. Blood 2004, 103, 3837–3844. [Google Scholar] [CrossRef]
- Munnur, D.; Teo, Q.; Eggermont, D.; Lee, H.H.Y.; Thery, F.; Ho, J.; van Leur, S.W.; Ng, W.W.S.; Siu, L.Y.L.; Beling, A.; et al. Altered ISGylation drives ab-errant macrophage-dependent immune responses during SARS-CoV-2 infection. Nat. Immunol. 2021, 22, 1416–1427. [Google Scholar] [CrossRef]
- Xu, C.; Gamil, A.A.A.; Munang’andu, H.M.; Evensen, O. Apoptosis Induction by dsRNA-Dependent Protein Kinase R (PKR) in EPC Cells via Caspase 8 and 9 Pathways. Viruses 2018, 10, 526. [Google Scholar] [CrossRef]
- Cavalcante, G.C.; Schaan, A.P.; Cabral, G.F.; Santana-Da-Silva, M.N.; Pinto, P.; Vidal, A.F.; Ribeiro-Dos-Santos, A. A Cell’s Fate: An Overview of the Molecular Biology and Genetics of Apoptosis. Int. J. Mol. Sci. 2019, 20, 4133. [Google Scholar] [CrossRef] [PubMed]
- Shinde, P.V.; Xu, H.C.; Maney, S.K.; Kloetgen, A.; Namineni, S.; Zhuang, Y.; Honke, N.; Shaabani, N.; Bellora, N.; Doerrenberg, M.; et al. Tumor Necrosis Factor-Mediated Survival of CD169 + Cells Promotes Immune Activation during Vesicular Stomatitis Virus Infection. J. Virol. 2018, 92, e01637-17. [Google Scholar] [CrossRef] [PubMed]
- Loureiro, J.; Ploegh, H.L. Antigen presentation and the ubiquitin-proteasome system in host-pathogen interactions. Adv. Immunol. 2006, 92, 225–305. [Google Scholar]
- Si, W.; Liang, H.; Bugno, J.; Xu, Q.; Ding, X.; Yang, K.; Fu, Y.; Weichselbaum, R.R.; Zhao, X.; Wang, L. Lactobacillus rhamnosus GG induces cGAS/STING- dependent type I interferon and improves response to immune checkpoint blockade. Gut 2022, 71, 521–533. [Google Scholar] [CrossRef] [PubMed]
- Marcus, P.I.; Sekellick, M.J.; Nichol, S.T. Interferon induction by viruses. XXI. Vesicular stomatitis virus: Interferon in-ducibility as a phylogenetic marker. J. Interferon Res. 1992, 12, 297–305. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).