Abstract
The quantity of seafood imported and produced by domestic aquaculture farming has increased. Recently, it has been reported that multidrug-resistant (MDR) Salmonella Typhimurium may be associated with seafood. However, information is limited to the antimicrobial resistance, virulence properties, and genetic diversity of S. Typhimurium recovered from imported and domestic seafood. This study investigated the antimicrobial resistance, virulence properties, and genetic diversity of S. Typhimurium isolated from domestic and imported catfish, shrimp, and tilapia. A total of 127 isolates were tested for the presence of multidrug-resistance (MDR), virulence genes (invA, pagC, spvC, spvR), and genetic diversity using the Sensititre micro-broth dilution method, PCR, and pulsed-field gel electrophoresis (PFGE), respectively. All isolates were uniformly susceptible to six (amoxicillin/clavulanic acid, ceftiofur, ceftriaxone, imipenem, nitrofurantoin, and trimethoprim/sulfamethoxazole) of the 17 tested antimicrobials and genetically diverse. Fifty-three percent of the Salmonella isolates were resistant to at least one antimicrobial and 49% were multidrug resistant. Ninety-five percent of the isolates possessed the invA gene, 67% pagC, and 43% for both spvC, and spvR. The results suggest that S. Typhimurium recovered from seafood is frequently MDR, virulent, and have the ability to cause salmonellosis.
1. Introduction
Salmonella belongs to the family Enterobacteriaceae and includes more than 2500 serovars. The occurrence of Salmonella in seafood is mainly due to cross-contamination associated with zoonotic and/or human involvement [1,2]. Salmonellosis is a global issue and one of the leading causes of foodborne illness in the U.S. that causes an estimated ~1.4 million non-typhoid cases of salmonellosis and 1.8 million cases of Typhoid salmonellosis per year resulting in 26,500 hospitalization and 420 fatalities [3,4,5,6]. Salmonella-contaminated food including seafood was linked to non-typhoidal human salmonellosis [7,8,9,10]. Approximately 3–10% of salmonellosis cases result in bacteremia requiring treatment with antimicrobials [5]. Serovar, S. Typhimurium is the most common cause of illnesses in the United States and accounts for most human infections [3,4,5,6].
Antimicrobials are growth promoters and therapeutics used to prevent or treat a wide range of pathogens, especially bacteria. The use of antimicrobials in food-producing animals, including seafood, is a concern for human health. The overuse and/or misuse of antimicrobials in aquaculture increases the development of antimicrobial resistance [11]. The risk of antimicrobial resistance development in bacteria within the environment, and the potential for the occurrence and/or maintenance of zoonotic pathogens in an aquatic population are also of concern [11,12,13,14]. Antimicrobial resistance increases mortality rates due to food-borne illness [15]. Although resistance, in particular multidrug-resistant (MDR), appears to be most serious in certain serotypes, this situation may be shifting. In China, clinical S. Typhimurium isolates with resistance to ciprofloxacin, ceftriaxone, and azithromycin were reported [16]. Beshiru et al. [17] reported that S. Typhimurium isolated from ready-to-eat shrimp in Nigeria was resistant to 11 antimicrobials which belong to 7 antimicrobial groups. In Spain, S. Typhimurium isolated from different types of seafood was found resistant to at least 6 antimicrobials [18]. Increased surveillance is needed to track the global spread of antimicrobial-resistant phenotypes of Salmonella isolates of food, animal, and human origin.
During the recent decades, per capita seafood consumption has increased in the U.S. The quantity of seafood imported and produced by domestic aquaculture farming increased; to 19.2 pounds per annum in 2019 [19]. Approximately 97% of the fish and shellfish are imported from overseas [20]. Nearly 50% of U.S. seafood imports are produced by aquaculture, and frozen seafood accounts for virtually 75% of the gross imports. Recent studies indicate that MDR Salmonella may be associated with seafood, especially shrimp, salmon, catfish, and tilapia [12,17,21,22,23].
The outcome of a Salmonella infection depends on the status of the host and the bacterium. Gene expression of various virulence factors is required for infection. The invasion of Salmonella into host cells is triggered by the invA gene [24,25]. The survival of Salmonella within macrophages is encoded by the pagC virulence gene. The prolific growth of salmonellae in host reticuloendothelial tissues depends on the spvC gene. Salmonella has been recovered from food, clinical and environmental samples that possessed multiple virulence factors [17,24,25,26,27,28,29].
Pulsed-field gel electrophoresis (PFGE) is a technique used at the molecular level to distinguish, identify, and evaluate the genetic diversity/relatedness and fingerprint database among bacterial strains including Salmonella from environmental, food and clinical strains of known human-pathogenic significance [22,30,31,32,33]. PFGE has been used to assess genetic-relatedness among Salmonella serotypes recovered from humans, sick food animals, and a variety of seafood [22,25,32,34]. Antimicrobial resistance, virulence properties and genetic diversity data for Salmonella recovered from imported and domestic seafood are limited. The study objective was to investigate the antimicrobial resistance, virulence properties, and genetic diversity of Salmonella recovered from seafood obtained from four retail stores located on the eastern shore of Maryland.
2. Materials and Methods
2.1. Salmonella Isolates
One hundred twenty-seven confirmed S. Typhimurium var 5 previously isolated from 440 domestic and imported frozen shrimp (domestic: Pandalus jordani, Litopenaeus setiferus, and Crangon franciscorum; imported: Litopenaeus vannamei, Penaeus merguensis, and Metapenaeus spp.—total n = 156), catfish (domestic: Ictalurus punctatus; imported: Pangasius swai—total n = 142), and tilapia (domestic: Oreochromis aureus; imported: Oreochromis niloticus—total n = 142) were used in this study. Of them, 62 isolates were from domestic, and 65 isolates were from imported seafood. All samples were purchased from four retail stores on the Eastern Shore of Maryland [34]. One isolate from each positive sample was selected for characterization using antimicrobial susceptibility testing, virulence genes, and PFGE.
2.2. Antimicrobial Susceptibility Testing
Minimal Inhibitory Concentrations (MIC) tests for Salmonella isolates were determined by broth microdilution and interpreted according to the Clinical and Laboratory Standards Institute’s guidelines [35,36]. In brief, different microdilutions of tested antimicrobials were made in Mueller Hinton broth (Thermo Fisher Scientific, Wilmington, DE, USA) considering their breakpoints. Each Salmonella isolate was added to all antimicrobial microdilutions. The minimal concentration that inhibited the growth of the pathogen was determined and compared to the breakpoints of each antimicrobial to determine whether the pathogen was susceptible, intermediate resistant, or resistant to the antimicrobial. Seventeen antimicrobials were tested (Trek Diagnostic Systems Inc., Cleveland, OH, USA) (Table 1). All antimicrobials were chosen based on the type of bacteria, their clinical usage for Salmonella in both humans and animals, as well as aquaculture practices. Escherichia coli ATCC 25922 and Pseudomonas aeruginosa ATCC 27853 were used as control strains.

Table 1.
Tested antimicrobials, concentrations, and breakpoints.
2.3. Detection of Virulence Genes
Salmonella isolates were tested for virulence genes (invA, pagC, spvC, and spvR) by PCR using the methods previously described [24,37,38]. In brief, genomic DNA for each isolate was extracted using the InstaGene matrix according to the manufacturer’s instructions (Bio-Rad Laboratories, Hercules, CA, USA). The DNA concentrations of Salmonella were determined using NanoDrop Lite, according to the manufacturer’s instructions (Thermo Fisher Scientific, Wilmington, DE, USA). Primers were used for Salmonella characterization following the manufacturer (Invitrogen by Life Technologies, Carlsbad, CA, USA) instructions (Table 2).
Table 2.
Nucleotide sequences for all targeted genes for Salmonella species [25].
GoTaq® Green Master Mix (Promega Corporation, Madison, WI, USA) was used according to the manufacturer’s guidelines. Twenty-five microliter reaction volumes were used; in which 12.5 µL of GoTaq Master Mix (2X) was added to 0.5 µL upstream primer and the same amount of downstream primer of each gene, 3 µL of DNA template and 8.5 µL nuclease-free water. Samples were loaded into the cycler and cycling conditions were adjusted for each targeted virulence gene as the denaturation, annealing, extension temperature and time varied depending on the testing genes [39,40] (Table 3).
Table 3.
The PCR conditions for Salmonella genes.
Salmonella Typhimurium strain Lt-2x3324 containing a recombinant plasmid with invA [41], S. Typhimurium containing a recombinant plasmid with spvC and spvR, and S. Typhimurium SR 11 x3337 containing a recombinant plasmid with pagC [42] were used as the positive controls and E. coli DH5-α (Invitrogen, Carlsbad, CA, USA) was used as the negative control for all reactions. Electrophoresis was carried out in a 1% agarose gel to separate the PCR products and the gels were stained with ethidium bromide.
2.4. Pulsed-Field Gel Electrophoresis of Salmonella
Pulsed-field Gel Electrophoresis (PFGE) technique was used to determine the change in the genomic profiles and was evaluated using XbaI (Boehringer Mannheim, Indianapolis, IN, USA) according to the guidelines developed by the Centers for Disease Control and Prevention (CDC) The CHEF-DR III SYSTEM (Bio-Rad Laboratories, Hercules, CA, USA) was used to perform electrophoresis using 1% SeaKem Gold agarose gel in 0.5X Tris-borate-EDTA (TBE) buffer at 14 °C for 19 h. The conditions of electrophoresis were as follows: initial switch time value of 2.16 s, final switch time of 63.8 s at a gradient of 6 V/cm, and an included angle of 120°. After electrophoresis, the gel was stained with ethidium bromide solution (40 mg/mL) and then de-stained with deionized water. DNA bands were visualized with a UV transilluminator and a digital image of PFGE patterns was acquired. To analyze the data BioNumerics software (Version 7.6, Applied Math, Sint-Martens-Latem, Belgium) was used. The relatedness between the strains was determined using the Dice Coefficient of similarity, and strains were grouped by hierarchal clustering of inter-strain similarities based on the Unweighted Pair Group Method with Arithmetic Averages (UPGMA) [37].
2.5. Statistical Analysis
Descriptive summaries of the frequency of antimicrobial resistance and prevalence of virulence genes were determined, as well as the number and frequency of antimicrobial resistance profiles among isolates that were resistant to one or more antimicrobials. Statistical significance of differences in the frequency of antimicrobial resistance of isolates by source (domestic vs. import) was determined for each antimicrobial agent using Fisher’s exact test. Fisher’s exact test was also used to determine the significance of differences in the presence of virulence genes in isolates from different types of seafood. All statistical calculations were performed using R version 4.1.2 [43]. Differences were considered significant when the p-value was less than 0.05.
3. Results
3.1. Prevalence of the Antimicrobial Resistance Phenotypes
All isolates were evaluated for antimicrobial resistance phenotypes using 17 different antimicrobials (Table 1).
Table 4 showed the antimicrobial resistance phenotypes of Salmonella recovered from all domestic and imported seafood. All tested Salmonella isolates were uniformly susceptible to six (amoxicillin/clavulanic acid, ceftiofur, ceftriaxone, imipenem, nitrofurantoin, and trimethoprim/sulfamethoxazole) of the 17 tested antimicrobials. Fifty-three percent of the Salmonella isolates (n = 67) were resistant to at least one antimicrobial and 49% of them (n = 63) were multidrug resistant. Forty-seven percent (n = 60) of all Salmonella isolates (23 isolates were domestic and 37 were imported) were susceptible to at least one of eleven tested antimicrobials.
Table 4.
Antimicrobial resistance phenotypes of Salmonella Typhimurium recovered from domestic and imported seafood.
Isolates recovered from domestic seafood were less resistant to some antimicrobials compared to those recovered from imported seafood. Salmonella-resistant isolates show high resistance against tetracycline (TET) (domestic 35% and imported 40%), nalidixic acid (NAL) (domestic 23% and imported 38%), gentamicin (Gen) (23% for the domestic and 34% for imported) and ciprofloxacin (CIP) (domestic showed low resistance (8%) in contrast to the imported (32%)). Differences in resistance to Ciprofloxacin, Chloramphenicol, Ampicillin, and Cefoxitin among isolates recovered from domestic versus imported seafood were statistically significant (p < 0.05).
A significant number of isolates from all seafood (n = 63) were MDR. Table 5 showed the antimicrobial resistance profiles observed among Salmonella isolates recovered from all domestic and imported seafood, respectively. Twenty-four antimicrobial profiles were observed among all Salmonella isolates, 12 of them among the domestic and 11 among the imported isolates. The most frequent multidrug resistance (MDR) profiles for domestic isolates were amikacin, ampicillin, cefoxitin, doxycycline, gentamicin, kanamycin, nalidixic acid, and tetracycline (AMI-AMP-FOX-DOX-GEN-KAN-NAL-TET), 9.7% (n = 6) and cefoxitin, ciprofloxacin, gentamicin, nalidixic acid, and tetracycline (FOX-CIP-GEN-NAL-TET) (9.2%) (n = 6) among imported seafood isolates. The second most prominent profiles among domestic were ampicillin, nalidixic acid, and tetracycline (AMP-NAL-TET), and doxycycline and tetracycline (DOX-TET) 6.4% (n = 4) each. Among the imported isolates, cefoxitin, chloramphenicol, ciprofloxacin, doxycycline, kanamycin, and nalidixic acid (FOX-CH-CIP-DOX-KAN-NAL) profile was rated second (8%) (n = 5) while ampicillin, cefoxitin, gentamicin, and tetracycline (AMP-FOX-GEN-TET) profile was rated third (6.2%) (n = 4).
Table 5.
Antimicrobial resistance profiles for Salmonella Typhimurium recovered from all seafood.
The antimicrobial profiles of Salmonella recovered from shrimp were varied and displayed nine profiles. Domestic isolates (n = 21) possessed three profiles including 15 isolates that were susceptible to all antimicrobials while imported (n = 20) exhibited six profiles; 6 were susceptible to all antimicrobials. The predominant profile was ampicillin-cefoxitin-gentamicin-tetracycline (AMP-FOX-GEN-TET) exhibited by four isolated recovered from imported seafood. The antimicrobial profiles for Salmonella recovered from tilapia were varied and displayed six profiles. The predominant profile for the isolates recovered from imported tilapia was cefoxitin-ciprofloxacin-gentamicin-nalidixic acid-tetracycline (FOX-CIP-GEN-NAL-TET). The antimicrobial profiles for Salmonella recovered from catfish were varied and displayed 12 profiles. Domestic isolates (n = 26) possessed eight profiles while imported (n = 15) exhibited four. The most frequent profiles within the isolates recovered from domestic catfish were amikacin-ampicillin-cefoxitin-doxycycline-florfenicol-gentamicin-kanamycin-nalidixic acid-tetracycline (AMI-AMP-FOX-DOX-FFN-GEN-KAN-NAL-TET). No apparent differences were observed among antibiotic resistance profiles of isolates from different types of seafood.
3.2. Characterization of Salmonella Isolates Recovered from Seafood for the Presence or Absence of Virulence Genes
Table 6 showed the presence of virulence genes invA, pagC, spvC, and spvR. The amplicon’s sizes for invA, pagC, and spvC genes were 284 bp, 318 bp, 571 bp, and 310 bp, respectively. Overall, 95% of the isolates were positive for the invA gene (91% of the isolates recovered from domestic seafood, and 100% of the isolates were recovered from imported seafood). Sixty-seven percent of the isolates possessed pagC, and 43% for both spvC and spvR. With respect to the seafood type and source, the invA gene was detected in 90% of the isolates recovered from domestic shrimp and 100% of the isolates recovered from imported shrimp. It was also detected in 86% and 100% of the isolates recovered from domestic and imported catfish, respectively. The gene was also detected in all isolates (100%) recovered from tilapia of both sources.
Table 6.
Prevalence of Salmonella Typhimurium virulence genes among tested isolates sizes.
The pagC gene was detected in 67% (n = 85) of the isolates. The gene was detected in 54% (n = 35) and 81% (n = 50) of the domestic and imported isolates, respectively. Fifty-two percent of the isolates recovered from the domestic and 95% of the isolates recovered from imported shrimp possessed the pagC gene. The gene was also detected in 46% and 43% of the isolates recovered from domestic and imported catfish, respectively. On the other hand, it was detected in 69% and 89% of isolates recovered from both domestic and imported tilapia, respectively. Similar prevalence of spvC and spvR genes were observed in all domestic and imported seafood. Forty-three percent of all isolates had both genes. The two genes were detected in 29% (n = 6) and 50% of domestic and imported shrimp 36% (n = 10) and 57% (n = 8) of the domestic and imported catfish, respectively. Both genes were also detected in 63% (n = 10) and 39% (n = 11) of the domestic and imported tilapia, respectively (Table 6). Except for the pagC gene, no statistically significant differences in the frequency of detection of the gene targets by seafood type were identified. For the pagC gene, the lower frequency of detection in isolates from catfish versus shrimp and tilapia was statistically significant (p < 0.05). There was no apparent association between the presence of the tested virulence genes and antimicrobial resistance profiles of Salmonella isolates.
3.3. Molecular Characterization of Salmonella Using Pulsed-Field Gel Electrophoresis—PFGE
Pulsed-field Gel Electrophoresis (PFGE) was used to appraise the genetic relatedness among the strains of the same pathogen recovered from the three types of seafood (shrimp, catfish, and tilapia). Salmonella isolates were digested with the restriction enzyme XbaI. Digestion of DNA resulted in 12 to 20 bands and the molecular weights of bands ranged from 50 to 1000 kbp.
The PFGE profiles of all Salmonella recovered from domestic (n = 62) and imported (n = 65) seafood combined (shrimp, catfish, and tilapia,) revealed 18 banding patterns which have been grouped into 14 clusters (from A to N) with 73% pattern of similarity. The similarity index of clusters ranged from 73% to 74% and the number of isolates in clusters was 1–30 (Figure 1).


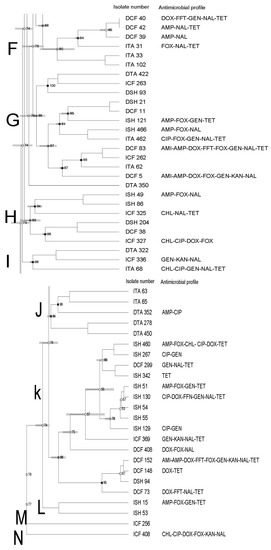
Figure 1.
Dendrogram of PFGE profiles of Salmonella Typhimurium isolated from domestic and imported seafood combined. The similarity index is indicated on the left axis. Abbreviations: DSH and ISH: domestic and imported shrimp, DCT and ICF: domestic and imported catfish and DTA and ITA: domestic imported tilapia. Antimicrobial key: Amikacin (AMI), ampicillin (AMP), cefoxitin (FOX), chloramphenicol (CHL), ciprofloxacin (CIP), ciprofloxacin (CIP), doxycycline (DOX), florfenicol (FFN), gentamicin (GEN), imipenem (IMI), kanamycin (KAN), nalidixic acid (NAL), tetracycline (TET). Letters A to N on the left represent pulsed-field gel electrophoresis clusters.
The PFGE profiles of all Salmonella (n = 62) recovered from domestic seafood (shrimp, catfish, and tilapia) revealed 18 banding patterns which have been grouped into four clusters (from A to D) with a 74% similarity index (Figure 2).

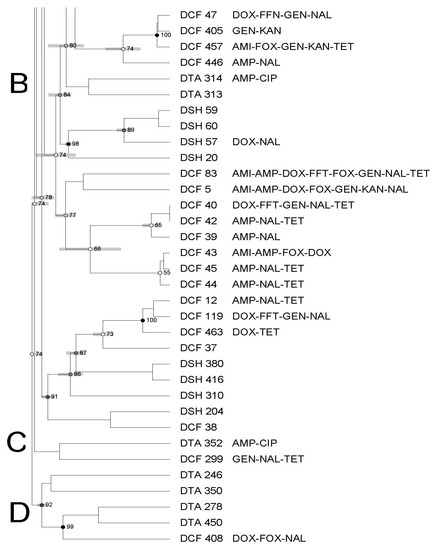
Figure 2.
Dendrogram of PFGE profiles of Salmonella Typhimurium isolated from all domestic seafood. The similarity index is indicated on the left axis. Abbreviations: DSH: domestic shrimp, DCT: domestic catfish and DTA: domestic tilapia. Antimicrobial’s key: amikacin (AMI), amoxycillin/clavulanic acid 2:1 (A.U.G2), ampicillin (AMP), ciprofloxacin (CIP), doxycycline (DOX), gentamicin (GEN), imipenem (IMI), nalidixic acid (NAL), nitrofurantoin (NIT), tetracycline (TET). Letters A to D on the left represent pulsed-field gel electrophoresis clusters.
The PFGE of all Salmonella (n = 65) recovered from all types of imported seafood (shrimp, catfish, and tilapia) revealed nine banding patterns which have been grouped into six clusters (from A to F) with an 82% similarity index (Figure 3).
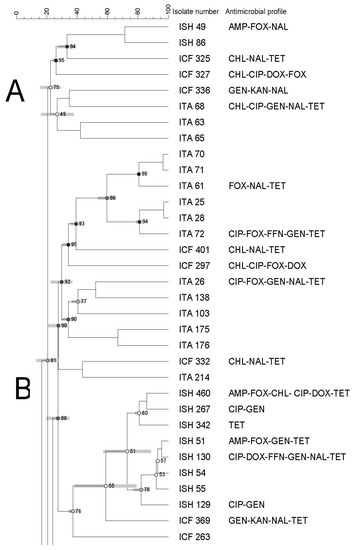
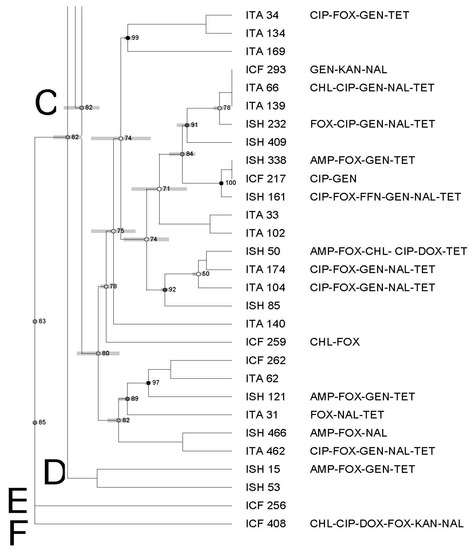
Figure 3.
Dendrogram of PFGE profiles of Salmonella Typhimurium isolated from all imported seafood. The similarity index is indicated on the left axis. Abbreviations: ISH: imported shrimp, ICT: imported catfish and ITA: imported tilapia. Antimicrobial’s key: amikacin (AMI), amoxycillin/clavulanic acid 2:1 (A.U.G2), ampicillin (AMP), ciprofloxacin (CIP), doxycycline (DOX), gentamicin (GEN), imipenem (IMI), nalidixic acid (NAL), nitrofurantoin (NIT), tetracycline (TET). Letters A to F on the left represent pulsed-field gel electrophoresis clusters.
Each cluster showed a different similarity index (74–92%), isolates (1–36 isolates), and antimicrobial resistance profiles. In addition, clusters contained isolates from different seafood types/sources (Figure 1, Figure 2 and Figure 3).
High genetic diversity was also observed, as evidenced by the dendrogram using both domestic and imported Salmonella isolates. Moreover, we did not observe any apparent association between antimicrobial resistance and PFGE profiles. There was also no association among PFGE profile, type of seafood, and country of origin through a few isolates displayed a tendency to cluster based on their source, type of seafood, and/or antimicrobial profiles. Although some isolates showed similar antimicrobial resistance profiles, they were genetically diverse.
4. Discussion
This study characterized 127 Salmonella Typhimurium isolates from imported and domestic catfish, shrimp, and tilapia for antimicrobial resistance profiles, virulence determinants, and genetic relatedness based on PFGE profiles. It is particularly notable that S. Typhimurium was the exclusive serovar isolated in this study in contrast to previous seafood surveys. Wild-caught domestic shrimp exhibited a lower incidence of resistance to individual antimicrobials and MDR than imported aquacultured shrimp consistent with previous studies (Table 7).
Table 7.
Comparison of antimicrobial resistance phenotypes of Salmonella Typhimurium recovered from domestic and imported seafood of our study to other studies.
Other studies key: Beshiru et al. [17] in Nigeria (#S1); Broughton and Walker, [44] in China (#S2), Das et al. [45] in Bangladesh (#S3); Karp et al. [46] in Taiwan (#S4), Obaidat and Salman [47] in Jordon (#S5), Wang et al. [48] in USA (#S6), Zhao et al. [49] in USA (#S7). N/A (not applicable): the drug was not used in the study. ND: not determined. Varies: other drugs used with variable resistance. T: Sensitive to all tested antibiotics. +: resistance was identified but the percentage was not determined in the study.
Most of the countries exporting seafood to the EU and the USA have adopted the same regulations as the country to which they export [50]. Differences in mandates between products for domestic consumption and the export market may impact the antimicrobial resistance profile of pathogens [51]. Predictably, resistance to tetracyclines (oxytetracycline is FDA approved) was greater than other antimicrobials that are not allowed for USA aquaculture. However, none of the isolates from either domestic or imported seafood were resistant to trimethoprim/sulfamethoxazole, which is FDA approved. Ciprofloxacin is in the fluoroquinolone class and is not FDA approved but is used in other countries [52]. Our results showed a lower resistance to ciprofloxacin among domestic isolates than imported isolates. All isolates in which ciprofloxacin was included in MDR were from imported seafood.
Antimicrobial profiles of Salmonella revealed resistance to the following antimicrobials: amikacin, ampicillin, cefoxitin, chloramphenicol, ciprofloxacin, doxycycline, florfenicol, gentamicin, kanamycin, nalidixic acid, and tetracycline. Salmonella isolates were susceptible to amoxicillin/clavulanic acid 2:1, ceftiofur, ceftriaxone, imipenem, and nitrofurantoin, as well as trimethoprim/sulfamethoxazole (Table 4). In comparison in a Bangladesh study on raw domestic shrimp from local farms, S. Typhimurium and S. enteritidis were isolated. S. Typhimurium was found to be resistant to amikacin, colistin, and erythromycin (Table 7). Moreover, it possessed intermediate resistance to ciprofloxacin, and kanamycin [45]. Beshiru et al. [17] in Nigeria stated that S. Typhimurium showed multidrug resistance to amoxicillin/clavulanic acid, ampicillin, doxycycline, and tetracycline. In addition, the bacterium showed resistance to amoxicillin, penicillin, and erythromycin (Table 7). The findings of this study contradicted the findings of Wang et al. [48] who stated that Salmonella isolates were susceptible to ampicillin, chloramphenicol, ciprofloxacin, gentamicin, and tetracycline (Table 7). Zhao et al. [49] demonstrated antimicrobial-resistant Salmonella serovars including Typhimurium recovered from imported seafood. They showed resistance to at least one antimicrobial: amoxicillin/clavulanic acid 2:1, ciprofloxacin, chloramphenicol, nalidixic acid, tetracycline, and trimethoprim/sulfamethoxazole. The later antimicrobial was found susceptible in our study (Table 7). Such difference may be due to the continual mutable paradigms of antimicrobials used in each country, the long-term and combined usage of antimicrobials, as well as the epidemiological surveillance objectives. Antimicrobial profiles were complex and constantly changing episodes in different environments.
As reported by Zhao et al. [49], the Food and Drug Administration (FDA) recovered 208 Salmonella isolates from >5000 samples of imported foods entering the USA in 2001, including aquatic foods such as lobster, catfish, shrimp, and many others. Different salmonellae were isolated and among them, S. Weltevreden (20%), S. Newport (6%), S. Lexington (5%), and S. Thompson (4%) were the foremost isolates. S. Typhimurium and other salmonellae were also isolated. The study of the antimicrobial resistance of these isolates reported that 11% were resistant to at least one antimicrobial and 3.4% were resistant to at least 3 antimicrobials [49] (Table 7). Obaidat and Salman [47] in their study in Jordon stated that 79% and 7% of Salmonella isolates recovered from imported seafood demonstrated antimicrobial resistance to at least single and multi-antimicrobials, respectively. It contradicted the findings of this study and showed high resistance toamoxicillin–clavulanic acid (52.2%), ampicillin (44.8%), and cephalothin (52.2%), as well as very low resistance to chloramphenicol (4.5%), ciprofloxacin (1.5%), gentamicin (6%), kanamycin (10.4%), nalidixic acid (4.5%), and tetracycline (9%). Obaidat and Salman’s [47] study exhibited very low resistance to trimethoprim/sulfamethoxazole (4.5%). Salmonella isolates of this study were susceptible to ceftriaxone (Table 7). The findings agreed with another study conducted in China which reported that Salmonella isolated from fish (perch, silver crab, and catfish) were resistant to ampicillin (20%), cotrimoxazole (20%), erythromycin (100%), gentamicin (20%), nalidixic acid (40%), nitrofurantoin (20%), penicillin (100%), streptomycin (20%), sulfonamide (40%), tetracycline (40%), and trimethoprim (20%). They showed intermediate resistance to ciprofloxacin (20%), chloramphenicol (20%), nalidixic acid (40%), and streptomycin (20%). The isolates were susceptible to cephalosporin, cefotaxime, ceftazidime, and neomycin [44] (Table 7). Karp et al. [46] determined a multidrug resistance Salmonella serotype Anatum isolated from U.S. clinical cases (travelers) and seafood from Asia (Taiwan) and U.S. Salmonella were resistant to amoxicillin/clavulanic acid, ampicillin, cefoxitin, chloramphenicol, streptomycin, sulfisoxazole, tetracycline, and trimethoprim/sulfamethoxazole (Table 7). In Nigeria, Beshiru et al. [17] reported multi-antimicrobial resistance for Salmonella Typhimurium; isolated from ready-to-eat shrimp to penicillin, erythromycin, ampicillin, amoxicillin, amoxicillin/clavulanic acid, and doxycycline. Several other investigators also have reported finding multidrug-resistant Salmonella isolates and these have been responsible for both human and animal salmonellosis worldwide [11,17,45,53]. These findings in addition to other studies including the current study confirmed the necessity for comprehensive worldwide cosmopolitan antimicrobial resistance surveillance (Table 7).
Salmonella Typhimurium recovered from domestic seafood showed low resistance (8%) to Ciprofloxacin (CIP) compared to imported seafood (32%). These results suggest the use/misuse of CIP or other fluoroquinolones in or near aquaculture facilities in the countries from which these seafood products were exported. Cabello [11] concluded that excessive use of antimicrobials as prophylactic or preventive medications could be a predisposing factor to antimicrobial resistance of bacteria recovered from seafood. Moreover, antimicrobial resistance may be transferred vertically among bacteria.
The virulence of Salmonella is associated with both chromosomal and plasmid-linked genes. This study investigated four major virulence genes: an invasion gene (invA) for epithelial cells invasion, a chromosomal virulence membrane gene (pagC) for survival within the macrophage, Salmonella plasmid virulence gene spvC (often named virA), and the transcriptional regulator, spvR, which is in an operon with the other three genes. The spv group potentiates the systemic spread of the pathogen and survival within the host cell [24].
The findings of this study reported the percentage of samples in which the different virulence genes invA (95%), pagC (67%), spvC (43%), and spvR (43%) could be detected. With the exception of pagC, the prevalence of these virulence determinants was not associated with seafood source or type (Table 6). No association between antimicrobial profiles of Salmonella isolates and seafood source or type is evident. All Salmonella isolates from fish were invA positive in an Indian study [25] and similar high prevalence results appeared universally [26,27,28,29] (Table 8). The findings above suggested that invA may be considered as a target gene to detect Salmonella irrespective of the other genes and all isolates recovered from food/environmental samples may not contain other virulence genes as clinical isolates.
Table 8.
Comparison of the prevalence of Salmonella Typhimurium virulence genes among tested isolates to other studies.
PFGE is a well-established epidemiological tool to assess diversity among Salmonella strains and suggests their relatedness regardless of their source (environment, seafood, and/or human clinical cases) [31,32,49]. Similar to previous investigations [33,34,52], the PFGE patterns of Salmonella in our study showed that Salmonella recovered from imported and domestic seafood are genetically diverse and did not show any concordance with source/type of seafood, antimicrobial resistance, or virulence properties.
The results of this study demonstrate a highly diverse population structure within the S. Typhimurium serovar in isolates from both domestic and imported seafood. Higher resistance levels to antimicrobials banned in the US such as CIP in imported seafood isolates suggest that their use is occurring around production environments in some export countries. Further studies to identify the genetic determinants linked to resistance patterns and their potential for horizontal transfer are warranted and whole genome sequencing (WGS) of the collection is currently under consideration.
Author Contributions
Conceptualization, S.M.E., J.B., A.D., M.J., F.H. and S.P.; Data curation, S.M.E., A.M.A., J.B. and L.V.D.S.; Formal analysis, S.M.E., A.M.A., J.B., A.D., A.J.P.-D., L.V.D.S. and S.P.; Funding acquisition, J.B., A.D., M.J. and S.P.; Investigation, S.M.E., A.M.A., A.J.P.-D. and S.P.; Methodology, S.M.E., A.M.A., A.J.P.-D. and S.P.; Project administration, S.P.; Resources, S.P.; Software, J.B.; Supervision, F.H. and S.P.; Validation, S.M.E. and S.P.; Visualization, S.M.E., A.M.A., J.B., A.D., M.J., L.V.D.S., F.H. and S.P.; Writing—original draft, S.M.E., A.M.A. and S.P.; Writing—review & editing, S.M.E., A.M.A., J.B., A.D., M.J., A.J.P.-D., L.V.D.S., F.H. and S.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by USDA Evans Allen grant # 2013-4-450510 and 2018-4-451641, and the US Department of Education Title III.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Upon request, data from this study will be made available by the corresponding author.
Acknowledgments
We would like to thank Joan Meredith and Mark Frana, for technical assistance and helpful suggestions.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Venkat, H.; Matthews, J.; Lumadao, P.; Caballero, B.; Collins, J.; Fowle, N.; Kellis, M.; Tewell, M.; White, S.; Hassan, R.; et al. Salmonella enterica serotype Javiana infections linked to a seafood restaurant in Maricopa county, Arizona, 2016. J. Food. Prot. 2018, 81, 1283–1292. [Google Scholar] [CrossRef]
- Kumar, R.; Datta, T.K.; Lalitha, K.V. Salmonella grows vigorously on seafood and expresses its virulence and stress genes at different temperature exposure. BMC Microbiol. 2015, 15, 254. [Google Scholar] [CrossRef]
- CDC. Salmonella, Salmonella Homepage. Available online: https://www.cdc.gov/salmonella/index.html (accessed on 6 February 2023).
- Iyer, A.; Kumosani, T.; Yousef, J.M.; Barbour, E.; Harakeh, S. Salmonella among humans of Asian countries. J. Life Sci. 2018, 15, 86–91. [Google Scholar]
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.-A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne illness acquired in the United States-major pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef]
- Lin, Y.; Fong, R.; Leo, J.; Kwan, W.; Ye, A.; Chan, P.; Chong, N.; Lim, G.; Octavia, S.; Lin, M. Distribution of Salmonella spp. along the food chain in Singapore, 2008–2016. Epidemiol. News Bull. 2019, 45, 44–54. [Google Scholar]
- Ferrari, R.G.; Rosario, D.K.A.; Cunha-Neto, A.; Mano, S.B.; Figueiredo, E.E.S.; Conte-Junior, C.A. Worldwide epidemiology of Salmonella serovars in animal-based foods: A Meta-analysis. Appl. Environ. Microbiol. 2019, 85, e00591-19. [Google Scholar] [CrossRef] [PubMed]
- Hassan, R.; Tecle, S.; Adcock, B.; Kellis, M.; Weiss, J.; Saupe, A.; Sorenson, A.; Klos, R.; Blankenship, J.; Blessington, T.; et al. Multistate outbreak of Salmonella Paratyphi B variant L(+) tartrate(+) and Salmonella Weltevreden infections linked to imported frozen raw tuna: USA, March-July 2015. Epidemiol. Infect. 2018, 146, 1461–1467. [Google Scholar] [CrossRef] [PubMed]
- Heinitz, M.L.; Ruble, R.D.; Wagner, D.E.; Tatini, S.R. Incidence of Salmonella in fish and seafood. J. Food. Prot. 2000, 63, 579–592. [Google Scholar] [CrossRef] [PubMed]
- CDC. Surveillance for Foodborne Disease Outbreaks, United States, 2015, Annual Report. Available online: https://www.cdc.gov/foodsafety/pdfs/2015foodborneoutbreaks_508.pdf (accessed on 23 May 2023).
- Cabello, F.C. Heavy use of prophylactic antibiotics in aquaculture: A growing problem for human and animal health and for the environment. Environ. Microbiol. 2006, 8, 1137–1144. [Google Scholar] [CrossRef]
- Elbashir, S.; Parveen, S.; Schwarz, J.; Rippen, T.; Jahncke, M.; DePaola, A. Seafood pathogens and information on antimicrobial resistance: A review. Food. Microbiol. 2018, 70, 85–93. [Google Scholar] [CrossRef]
- CDC. National Antimicrobial Resistance Monitoring System for Enteric Bacteria (NARMS): Human Isolates Surveillance Report for 2015 (Final Report). Available online: https://www.cdc.gov/narms/pdf/2015-NARMS-Annual-Report-cleared_508.pdf (accessed on 23 May 2023).
- CDC. Antibiotic Resistance Threats in the United States. 2013. Available online: https://www.cdc.gov/drugresistance/pdf/ar-threats-2013-508.pdf (accessed on 20 September 2022).
- Helms, M.; Vastrup, P.; Gerner-Smidt, P.; Mølbak, K. Excess mortality associated with antimicrobial drug-resistant Salmonella Typhimurium. Emerg. Infect. Dis. 2002, 8, 490–495. [Google Scholar] [CrossRef] [PubMed]
- Wong, M.H.Y.; Yan, M.; Chan, E.W.C.; Biao, K.; Chen, S. Emergence of clinical Salmonella enterica serovar Typhimurium isolates with concurrent resistance to ciprofloxacin, ceftriaxone, and azithromycin. Antimicrob. Agents Chemother. 2014, 58, 3752–3756. [Google Scholar] [CrossRef]
- Beshiru, A.; Igbinosa, I.H.; Igbinosa, E.O. Prevalence of antimicrobial resistance and virulence gene elements of Salmonella serovars from Ready-to-Eat (RTE) shrimps. Front. Microbiol. 2019, 10, 01613. [Google Scholar] [CrossRef]
- Martinez-Urtaza, J.; Liebana, E.; Garcia-Migura, L.; Perez-Piñeiro, P.; Saco, M. Characterization of Salmonella enterica serovar typhimurium from marine environments in coastal waters of Galicia (Spain). Appl. Environ. Microbiol. 2004, 70, 4030–4034. [Google Scholar] [CrossRef]
- National Marine Fisheries Service. Fisheries of the United States. 2019. Available online: https://www.fisheries.noaa.gov/national/sustainable-fisheries/fisheries-united-states (accessed on 18 October 2021).
- Gould, L.H.; Kline, J.; Monahan, C.; Vierk, K. Outbreaks of disease associated with food imported into the United States, 1996–2014. Emerg. Infect. Dis. 2017, 23, 525–528. [Google Scholar] [CrossRef] [PubMed]
- Hounmanou, Y.M.G.; Dalsgaard, A.; Sopacua, T.F.; Uddin, G.M.N.; Leekitcharoenphon, P.; Hendriksen, R.S.; Olsen, J.E.; Larsen, M.H. Molecular characteristics and zoonotic potential of Salmonella Weltevreden from cultured shrimp and tilapia in Vietnam and China. Front. Microbiol. 2020, 11, 01985. [Google Scholar] [CrossRef] [PubMed]
- Budiati, T.; Rusul, G.; Wan-Abdullah, W.N.; Chuah, L.O.; Ahmad, R.; Thong, K.L. Geneticrelatedness of Salmonella serovars isolated from catfish (Clarias gariepinus) and tilapia (Tilapia mossambica) obtained from wet markets and ponds in Penang, Malaysia. J. Food. Prot. 2016, 79, 659–665. [Google Scholar] [CrossRef]
- Friesema, I.; de Jong, A.; Hofhuis, A.; Heck, M.; van den Kerkhof, H.; de Jonge, R.; Hameryck, D.; Nagel, K.; van Vilsteren, G.; van Beek, P.; et al. Large outbreak of Salmonella Thompson related to smoked salmon in the Netherlands, August to December 2012. Euro Surveill. 2014, 19, 20918. [Google Scholar] [CrossRef]
- Chaudhary, J.H.; Nayak, J.B.; Brahmbhatt, M.N.; Makwana, P.P. Virulence genes detection of Salmonella serovars isolated from pork and slaughterhouse environment in Ahmedabad, Gujarat. Vet. World 2015, 8, 121–124. [Google Scholar] [CrossRef]
- Tekale, A.; Savalia, C.; Kshirsagar, D.; Brahmbhatt, M.; Chatur, Y. Detection and virulence gene characterization of Salmonella isolates from fish by conventional and molecular methods. J. Vet. Public Health 2015, 13, 43–46. [Google Scholar]
- Kumar, P.; Agarwal, R.; Avinash, R.; Prasannavadhana, A.; Thomas, P.; Kumar, A.; Kataria, J.; Kerketta, P. Loop mediated isothermal amplification (LAMP) test for rapid detection of Salmonella in the chicken meat. J. Vet. Public Health 2014, 12, 7–12. [Google Scholar]
- Akiyama, T.; Khan, A.A.; Cheng, C.M.; Stefanova, R. Molecular characterization of Salmonella enterica serovar Saintpaul isolated from imported seafood, pepper, environmental and clinical samples. Food. Microbiol. 2011, 28, 1124–1128. [Google Scholar] [CrossRef] [PubMed]
- Bhatta, D.R.; Bangtrakulnonth, A.; Tishyadhigama, P.; Saroj, S.D.; Bandekar, J.R.; Hendriksen, R.S.; Kapadnis, B.P. Serotyping, PCR, phage-typing and antibiotic sensitivity testing of Salmonella serovars isolated from urban drinking water supply systems of Nepal. Lett. Appl. Microbiol. 2007, 44, 588–594. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, S.D.d.; Rodenbusch, C.R.; Michael, G.B.; Cardoso, M.I.R.; Canal, C.W.; Brandelli, A. Detection of virulence genes in Salmonella Enteritidis isolated from different sources. Braz. J. Microbiol. 2003, 34, 123–124. [Google Scholar] [CrossRef]
- Bhowmick, P.P.; Srikumar, S.; Devegowda, D.; Shekar, M.; Darshanee Ruwandeepika, H.A.; Karunasagar, I. Serotyping & molecular characterization for study of genetic diversity among seafood associated nontyphoidal Salmonella serovars. Indian J. Med. Res. 2012, 135, 371–381. [Google Scholar]
- Rodriguez-Castro, A.; Ansede-Bermejo, J.; Blanco-Abad, V.; Varela-Pet, J.; Garcia-Martin, O.; Martinez-Urtaza, J. Prevalence and genetic diversity of pathogenic populations of Vibrio parahaemolyticus in coastal waters of Galicia, Spain. Environ. Microbiol. Rep. 2010, 2, 58–66. [Google Scholar] [CrossRef]
- Zhao, S.; Qaiyumi, S.; Friedman, S.; Singh, R.; Foley, S.L.; White, D.G.; McDermott, P.F.; Donkar, T.; Bolin, C.; Munro, S.; et al. Characterization of Salmonella enterica serotype Newport isolated from humans and food animals. J. Clin. Microbiol. 2003, 41, 5366–5371. [Google Scholar] [CrossRef] [PubMed]
- Swaminathan, B.; Barrett, T.J.; Hunter, S.B.; Tauxe, R.V. PulseNet: The molecular subtyping network for foodborne bacterial disease surveillance, United States. Emerg. Infect. Dis. 2001, 7, 382–389. [Google Scholar] [CrossRef]
- Elbashir, S.; Jahncke, M.; DePaola, A.; Bowers, J.; Schwarz, J.; Punchihewage-Don, A.J.; Min, B.; Rippen, T.; Parveen, S. Prevalence and abundance of bacterial pathogens of concern in shrimp, catfish and tilapia obtained at retail stores in Maryland, USA. Pathogens 2023, 12, 187. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Methods for Dilutions Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically. In Approved Standard M7–A7, 7th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2006. [Google Scholar]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing; 20th Informational Supplement (June 2010 Update); CLSI Document M100-S20-U; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2010. [Google Scholar]
- Mohamed, T.; Zhao, S.; White, D.G.; Parveen, S. Molecular characterization of antibiotic resistant Salmonella Typhimurium and Salmonella Kentucky isolated from pre- and post-chill whole broilers carcasses. Food Microbiol. 2014, 38, 6–15. [Google Scholar] [CrossRef]
- Olah, P.A.; Sherwood, J.S.; Logue, C.M. Molecular analysis of Salmonella isolates recovered from processed Turkey carcasses. J. Food Prot. 2005, 68, 845–849. [Google Scholar] [CrossRef]
- Rahn, K.; De Grandis, S.A.; Clarke, R.C.; McEwen, S.A.; Galán, J.E.; Ginocchio, C.; Curtiss, R., 3rd; Gyles, C.L. Amplification of an invA gene sequence of Salmonella Typhimurium by polymerase chain reaction as a specific method of detection of Salmonella. Mol. Cell. Probes. 1992, 6, 271–279. [Google Scholar] [CrossRef]
- Lampel, K.A.; Orlandi, P.A.; Kornegay, L. Improved template preparation for PCR-based assays for detection of food-borne bacterial pathogens. Appl. Environ. Microbiol. 2000, 66, 4539–4542. [Google Scholar] [CrossRef]
- Galán, J.E.; Curtiss, R., 3rd. Cloning and molecular characterization of genes whose products allow Salmonella Typhimurium to penetrate tissue culture cells. Proc. Natl. Acad. Sci. USA 1989, 86, 6383–6387. [Google Scholar] [CrossRef] [PubMed]
- Gulig, P.A.; Danbara, H.; Guiney, D.G.; Lax, A.J.; Norel, F.; Rhen, M. Molecular analysis of spv virulence genes of the Salmonella virulence plasmids. Mol. Microbiol. 1993, 7, 825–830. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Broughton, E.I.; Walker, D.G. Prevalence of antibiotic-resistant Salmonella in fish in Guangdong, China. Foodborne Pathog. Dis. 2009, 6, 519–521. [Google Scholar] [CrossRef] [PubMed]
- Das, P.K.; Mandal, A.; Rahman, M.M.; Sarkar, S.L.; Jahid, I.K.; Hossain, M.A.; Alam, A.S.M.R.U.; Roy, P.C. Salmonella enterica serovar Typhimurium and Enteritidis isolated from raw shrimp in Bangladesh: An investigation based on molecular characteristics, survival, virulence, antibiotic resistance, and biofilm formation attributes. J. Food Qual. 2022, 2022, 3420364. [Google Scholar] [CrossRef]
- Karp, B.E.; Leeper, M.M.; Chen, J.C.; Tagg, K.A.; Francois Watkins, L.K.; Friedman, C.R. Multidrug-resistant Salmonella serotype Anatum in travelers and seafood from Asia, United States. Emerg. Infect. Dis. 2020, 26, 1030–1033. [Google Scholar] [CrossRef]
- Obaidat, M.M.; Bani Salman, A.E. Antimicrobial resistance percentages of Salmonella and Shigella in seafood imported to Jordan: Higher percentages and more diverse profiles in Shigella. J. Food. Prot. 2017, 80, 414–419. [Google Scholar] [CrossRef]
- Wang, F.; Jiang, L.; Yang, Q.; Han, F.; Chen, S.; Pu, S.; Vance, A.; Ge, B. Prevalence and antimicrobial susceptibility of major foodborne pathogens in imported seafood. J. Food Prot. 2011, 74, 1451–1461. [Google Scholar] [CrossRef]
- Zhao, S.; McDermott, P.F.; Friedman, S.; Qaiyumi, S.; Abbott, J.; Kiessling, C.; Ayers, S.; Singh, R.; Hubert, S.; Sofos, J.; et al. Characterization of antimicrobial-resistant Salmonella isolated from imported foods. J. Food Prot. 2006, 69, 500–507. [Google Scholar] [CrossRef] [PubMed]
- Braak, I.K.V. Opportunities to Improve Quality of Exports of Indian Seafood. Available online: https://www.rvo.nl/sites/default/files/2022-04/Opportunities%20to%20improve%20quality%20of%20exports%20of%20Indian%20Seafood.pdf (accessed on 8 May 2023).
- Henriksson, P.J.G.; Rico, A.; Troell, M.; Klinger, D.H.; Buschmann, A.H.; Saksida, S.; Chadag, M.V.; Zhang, W. Unpacking factors influencing antimicrobial use in global aquaculture and their implication for management: A review from a systems perspective. Sustain. Sci. 2018, 13, 1105–1120. [Google Scholar] [CrossRef]
- Guidi, L.R.; Santos, F.A.; Ribeiro, A.C.S.R.; Fernandes, C.; Silva, L.H.M.; Gloria, M.B.A. Quinolones and tetracyclines in aquaculture fish by a simple and rapid LC-MS/MS method. Food Chem. 2018, 245, 1232–1238. [Google Scholar] [CrossRef] [PubMed]
- Adesiji, Y.O.; Kogaluru Shivakumaraswamy, S.; Kumar Deekshit, V.; Shivani Kallappa, G.; Karunasagar, I. Molecular characterization of antimicrobial multi-drug resistance in non-typhoidal Salmonellae from chicken and clam in Mangalore, India. J. Biomed. Res. 2017, 32, 237–244. [Google Scholar] [CrossRef]
- Nolan, L.K.; Giddings, C.W.; Brown, J. The distribution of invA, pagC and spvC genes among Salmonella isolates from animals. Vet. Res. Commun. 1995, 19, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Soto, S.M.; Rodríguez, I.; Rodicio, M.R.; Vila, J.; Mendoza, M.C. Detection of virulence determinants in clinical strains of Salmonella enterica serovar Enteritidis and mapping on macrorestriction profiles. J. Med. Microbiol. 2006, 55, 365–373. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).