
Evaluation of Genetic Diversity and Virulence Potential of Legionella pneumophila Isolated from Water Supply Systems of Residential Buildings in Latvia

 , ,
, ,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Microbiological Testing of Legionella spp.
2.3. DNA Extraction from Legionella Isolates
2.4. Whole-Genome Sequencing
2.5. Data Analysis
3. Results
3.1. Prevalence of L. pneumophila in Residential Buildings
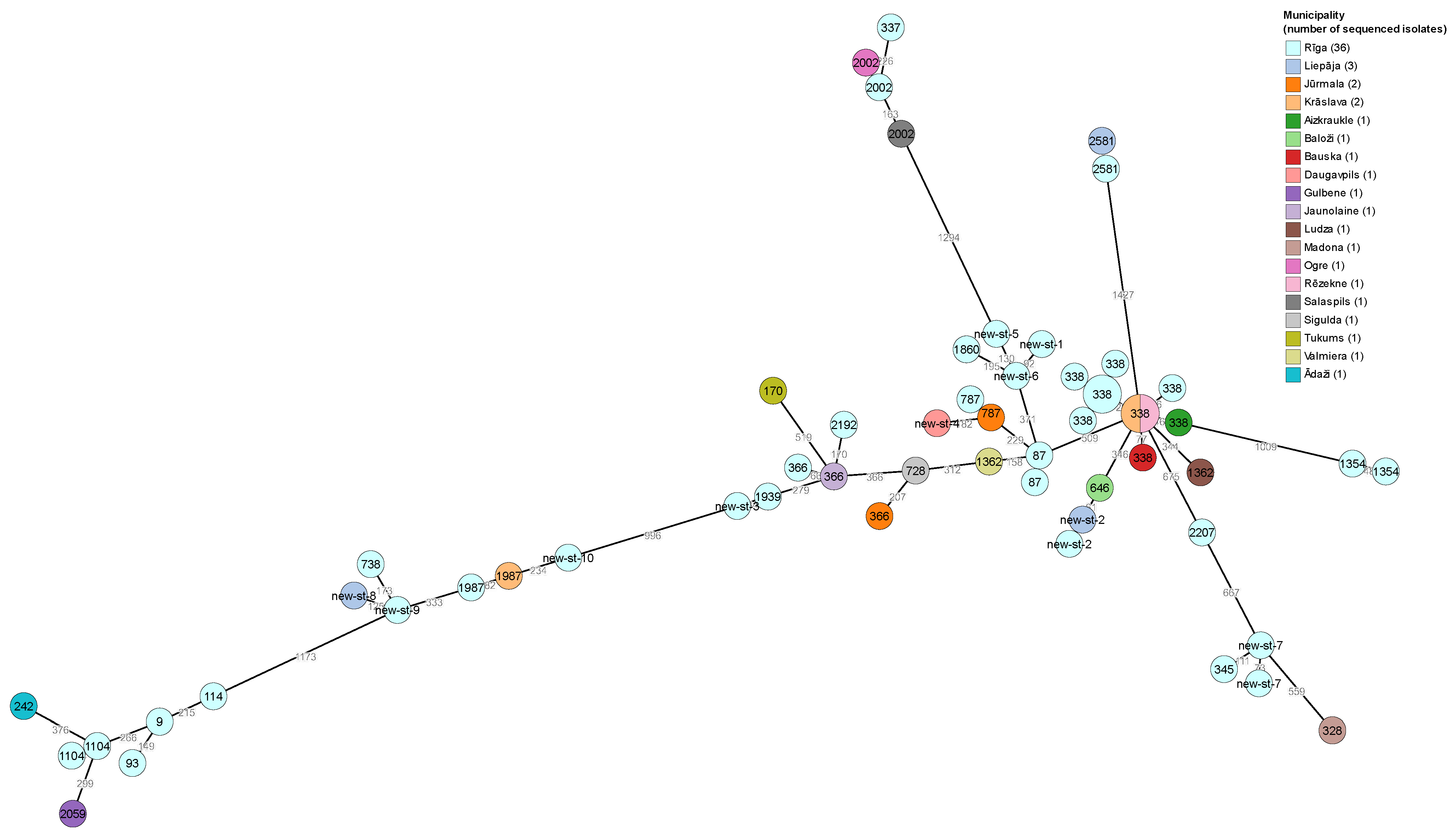
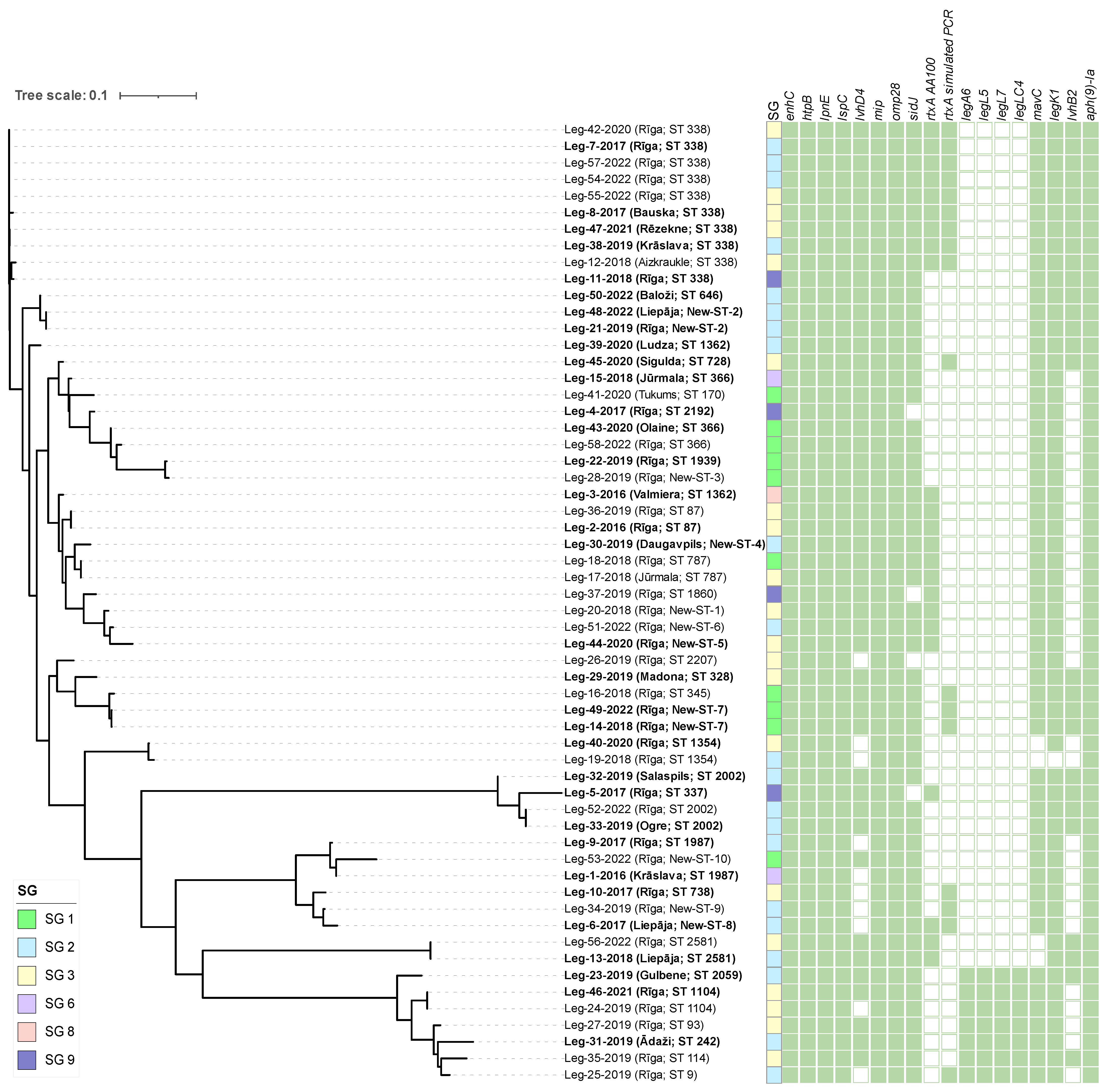
3.2. Whole-Genome Sequencing of L. pneumophila
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mondino, S.; Schmidt, S.; Rolando, M.; Escoll, P.; Gomez-Valero, L.; Buchrieser, C. Legionnaires’ Disease: State of the Art Knowledge of Pathogenesis Mechanisms of Legionella. Annu. Rev. Pathol. Mech. Dis. 2020, 15, 439–466. [Google Scholar] [CrossRef] [PubMed]
- Legionnaires’ Disease—Annual Epidemiological Report for 2020. Available online: https://www.ecdc.europa.eu/en/publications-data/legionnaires-disease-annual-epidemiological-report-2020 (accessed on 17 March 2023).
- Byrne, B.G.; McColm, S.; McElmurry, S.P.; Kilgore, P.E.; Sobeck, J.; Sadler, R.; Love, N.G.; Swanson, M.S. Prevalence of Infection-Competent Serogroup 6 Legionella pneumophila within Premise Plumbing in Southeast Michigan. mBio 2018, 9, e00016-18. [Google Scholar] [CrossRef] [PubMed]
- Yu, A.T.; Kamali, A.; Vugia, D.J. Legionella Epidemiologic and Environmental Risks. Curr. Epidemiol. Rep. 2019, 6, 310–320. [Google Scholar] [CrossRef]
- Kessler, M.A.; Osman, F.; Marx, J.; Pop-Vicas, A.; Safdar, N. Hospital-Acquired Legionella Pneumonia Outbreak at an Academic Medical Center: Lessons Learned. Am. J. Infect. Control 2021, 49, 1014–1020. [Google Scholar] [CrossRef]
- De Filippis, P.; Mozzetti, C.; Messina, A.; D’Alò, G.L. Prevalence of Legionella in Retirement Homes and Group Homes Water Distribution Systems. Sci. Total Environ. 2018, 643, 715–724. [Google Scholar] [CrossRef]
- Mentula, S.; Kääriäinen, S.; Jaakola, S.; Niittynen, M.; Airaksinen, P.; Koivula, I.; Lehtola, M.; Mauranen, E.; Mononen, I.; Savolainen, R.; et al. Tap Water as the Source of a Legionnaires’ Disease Outbreak Spread to Several Residential Buildings and One Hospital, Finland, 2020 to 2021. Eurosurveillance 2023, 28, 2200673. [Google Scholar] [CrossRef]
- Nisar, M.A.; Ross, K.E.; Brown, M.H.; Bentham, R.; Whiley, H. Water Stagnation and Flow Obstruction Reduces the Quality of Potable Water and Increases the Risk of Legionelloses. Front. Environ. Sci. 2020, 8, 611611. [Google Scholar] [CrossRef]
- Singh, R.; Chauhan, D.; Fogarty, A.; Rasheduzzaman, M.; Gurian, P.L. Practitioners’ Perspective on the Prevalent Water Quality Management Practices for Legionella Control in Large Buildings in the United States. Water 2022, 14, 663. [Google Scholar] [CrossRef]
- Kruse, E.-B.; Wehner, A.; Wisplinghoff, H. Prevalence and Distribution of Legionella spp. in Potable Water Systems in Germany, Risk Factors Associated with Contamination, and Effectiveness of Thermal Disinfection. Am. J. Infect. Control 2016, 44, 470–474. [Google Scholar] [CrossRef]
- Ge, Z.; Yuan, P.; Chen, L.; Chen, J.; Shen, D.; She, Z.; Lu, Y. New Global Insights on the Regulation of the Biphasic Life Cycle and Virulence via ClpP-Dependent Proteolysis in Legionella pneumophila. Mol. Cell. Proteom. 2022, 21, 100233. [Google Scholar] [CrossRef]
- Chauhan, D.; Shames, S.R. Pathogenicity and Virulence of Legionella: Intracellular Replication and Host Response. Virulence 2021, 12, 1122–1144. [Google Scholar] [CrossRef] [PubMed]
- Gattuso, G.; Rizzo, R.; Lavoro, A.; Spoto, V.; Porciello, G.; Montagnese, C.; Cinà, D.; Cosentino, A.; Lombardo, C.; Mezzatesta, M.L.; et al. Overview of the Clinical and Molecular Features of Legionella pneumophila: Focus on Novel Surveillance and Diagnostic Strategies. Antibiotics 2022, 11, 370. [Google Scholar] [CrossRef] [PubMed]
- Abu Khweek, A.; Amer, A.O. Factors Mediating Environmental Biofilm Formation by Legionella pneumophila. Front. Cell. Infect. Microbiol. 2018, 8, 38. [Google Scholar] [CrossRef]
- Scheithauer, L.; Karagöz, M.S.; Mayer, B.E.; Steinert, M. Protein Sociology of ProA, Mip and Other Secreted Virulence Factors at the Legionella pneumophila Surface. Front. Cell. Infect. Microbiol. 2023, 13, 1140688. [Google Scholar] [CrossRef]
- Yang, J.-L.; Li, D.; Zhan, X.-Y. Concept about the Virulence Factor of Legionella. Microorganisms 2022, 11, 74. [Google Scholar] [CrossRef]
- Centre of Disease Prevention and Control of Latvia. Epidemiological Reports|SPKC. 2023. Available online: https://www.spkc.gov.lv/lv/epidemiologijas-bileteni (accessed on 2 April 2023).
- ISO 19458:2006; Water Quality—Sampling for Microbiological Analysis. International Organization for Standardization: Geneva, Switzerland, 2006. Available online: https://www.iso.org/standard/33845.html (accessed on 15 November 2022).
- ISO 11731:2017; Water Quality—Enumeration of Legionella. International Organization for Standardization: Geneva, Switzerland, 2017. Available online: https://www.iso.org/standard/61782.html (accessed on 15 November 2022).
- Valciņa, O.; Pūle, D.; Ķibilds, J.; Lazdāne, A.; Trofimova, J.; Makarova, S.; Konvisers, G.; Ķimse, L.; Krūmiņa, A.; Bērziņš, A. Prevalence and Genetic Diversity of Legionella spp. In Hotel Water-Supply Systems in Latvia. Microorganisms 2023, 11, 596. [Google Scholar] [CrossRef] [PubMed]
- Prjibelski, A.D.; Puglia, G.D.; Antipov, D.; Bushmanova, E.; Giordano, D.; Mikheenko, A.; Vitale, D.; Lapidus, A. Extending RnaSPAdes Functionality for Hybrid Transcriptome Assembly. BMC Bioinform. 2020, 21, 302. [Google Scholar] [CrossRef] [PubMed]
- Gaia, V.; Fry, N.K.; Afshar, B.; Lück, P.C.; Meugnier, H.; Etienne, J.; Peduzzi, R.; Harrison, T.G. Consensus Sequence-Based Scheme for Epidemiological Typing of Clinical and Environmental Isolates of Legionella pneumophila. J. Clin. Microbiol. 2005, 43, 2047–2052. [Google Scholar] [CrossRef] [PubMed]
- Mentasti, M.; Underwood, A.; Lück, C.; Kozak-Muiznieks, N.A.; Harrison, T.G.; Fry, N.K. Extension of the Legionella pneumophila Sequence-Based Typing Scheme to Include Strains Carrying a Variant of the N-Acylneuraminate Cytidylyltransferase Gene. Clin. Microbiol. Infect. 2014, 20, O435–O441. [Google Scholar] [CrossRef][Green Version]
- Moran-Gilad, J.; Prior, K.; Yakunin, E.; Harrison, T.G.; Underwood, A.; Lazarovitch, T.; Valinsky, L.; Lück, C.; Krux, F.; Agmon, V.; et al. Design and Application of a Core Genome Multilocus Sequence Typing Scheme for Investigation of Legionnaires’ Disease Incidents. Eurosurveillance 2015, 20, 21186. [Google Scholar] [CrossRef] [PubMed]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.G.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid Large-Scale Prokaryote Pan Genome Analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef] [PubMed]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately Maximum-Likelihood Trees for Large Alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zheng, D.; Zhou, S.; Chen, L.; Yang, J. VFDB 2022: A General Classification Scheme for Bacterial Virulence Factors. Nucleic Acids Res. 2021, 50, D912–D917. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A Greedy Algorithm for Aligning DNA Sequences. J. Comput. Biol. 2000, 7, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Samrakandi, M.M.; Cirillo, S.L.G.; Ridenour, D.A.; Bermudez, L.E.; Cirillo, J.D. Genetic and Phenotypic Differences between Legionella pneumophila Strains. J. Clin. Microbiol. 2002, 40, 1352–1362. [Google Scholar] [CrossRef]
- Slater, G.; Birney, E. Automated Generation of Heuristics for Biological Sequence Comparison. BMC Bioinform. 2005, 6, 31. [Google Scholar] [CrossRef]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for Predictions of Phenotypes from Genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef]
- Liu, M.; Conover, G.M.; Isberg, R.R. Legionella pneumophila EnhC Is Required for Efficient Replication in Tumour Necrosis Factor α-Stimulated Macrophages. Cell. Microbiol. 2008, 10, 1906–1923. [Google Scholar] [CrossRef]
- Hoffman, P.C.; Garduno, R.A. Surface-Associated Heat Shock Proteins Of Legionella pneumophila and Helicobacter Pylori: Roles in Pathogenesis and Immunity. Infect. Dis. Obstet. Gynecol. 1999, 7, 58–63. [Google Scholar] [CrossRef]
- Helbig, J.H.; König, B.; Knospe, H.; Bubert, B.; Yu, C.; Lück, C.P.; Riboldi-Tunnicliffe, A.; Hilgenfeld, R.; Jacobs, E.; Hacker, J.; et al. The PPIase Active Site of Legionella pneumophila Mip Protein Is Involved in the Infection of Eukaryotic Host Cells. Biol. Chem. 2003, 384, 125–137. [Google Scholar] [CrossRef]
- Bellinger-Kawahara, C.; Horwitz, M.A. Complement Component C3 Fixes Selectively to the Major Outer Membrane Protein (MOMP) of Legionella pneumophila and Mediates Phagocytosis of Liposome-MOMP Complexes by Human Monocytes. J. Exp. Med. 1990, 172, 1201–1210. [Google Scholar] [CrossRef] [PubMed]
- Gan, N.; Nakayasu, E.S.; Hollenbeck, P.J.; Luo, Z.-Q. Legionella pneumophila Inhibits Immune Signalling via MavC-Mediated Transglutaminase-Induced Ubiquitination of UBE2N. Nat. Microbiol. 2018, 4, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Losick, V.P.; Haenssler, E.; Moy, M.-Y.; Isberg, R.R. LnaB: A Legionella pneumophila Activator of NF-ΚB. Cell. Microbiol. 2010, 12, 1083–1097. [Google Scholar] [CrossRef]
- Bhogaraju, S.; Bonn, F.; Mukherjee, R.; Adams, M.; Pfleiderer, M.M.; Galej, W.P.; Matkovic, V.; Lopez-Mosqueda, J.; Kalayil, S.; Shin, D.; et al. Inhibition of Bacterial Ubiquitin Ligases by SidJ–Calmodulin Catalysed Glutamylation. Nature 2019, 572, 382–386. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, P.; Lang, E.A.S.; Rasaputra, K.S.; Steinman, H.M. Implication of the VirD4 Coupling Protein of the Lvh Type 4 Secretion System in Virulence Phenotypes of Legionella pneumophila. J. Bacteriol. 2013, 195, 3468–3475. [Google Scholar] [CrossRef] [PubMed]
- Newton, H.J.; Browning, G.F.; Dao, J.; McAlister, A.D.; Sloan, J.; Cianciotto, N.P.; Hartland, E.L. Sel1 Repeat Protein LpnE Is a Legionella pneumophila Virulence Determinant That Influences Vacuolar Trafficking. Infect. Immun. 2007, 75, 5575–5585. [Google Scholar] [CrossRef]
- Rossier, O.; Starkenburg, S.R.; Cianciotto, N.P. Legionella pneumophila Type II Protein Secretion Promotes Virulence in the A/J Mouse Model of Legionnaires’ Disease Pneumonia. Infect. Immun. 2003, 72, 310–321. [Google Scholar] [CrossRef]
- Cirillo, S.L.G.; Bermudez, L.E.; El-Etr, S.H.; Duhamel, G.E.; Cirillo, J.D. Legionella pneumophila Entry Gene RtxA Is Involved in Virulence. Infect. Immun. 2001, 69, 508–517. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023; Available online: https://www.R-project.org/ (accessed on 6 April 2023).
- Letunic, I.; Bork, P. Interactive Tree of Life (ITOL) V5: An Online Tool for Phylogenetic Tree Display and Annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Zhou, Z.; Alikhan, N.-F.; Sergeant, M.J.; Luhmann, N.; Vaz, C.; Francisco, A.P.; Carriço, J.A.; Achtman, M. GrapeTree: Visualization of Core Genomic Relationships among 100,000 Bacterial Pathogens. Genome Res. 2018, 28, 1395–1404. [Google Scholar] [CrossRef]
- Dilger, T.; Melzl, H.; Gessner, A. Legionella Contamination in Warm Water Systems: A Species-Level Survey. Int. J. Hyg. Environ. Health 2018, 221, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Felice, A.; Franchi, M.; De Martin, S.; Vitacolonna, N.; Iacumin, L.; Civilini, M. Environmental Surveillance and Spatio-Temporal Analysis of Legionella spp. in a Region of Northeastern Italy (2002–2017). PLoS ONE 2019, 14, e0218687. [Google Scholar] [CrossRef] [PubMed]
- Gleason, J.A.; Conner, L.E.; Ross, K.M. Associations of Household Factors, Hot Water Temperature, and Chlorine Residual with Legionella Occurrence in Single-Family Homes in New Jersey. Sci. Total Environ. 2023, 870, 161984. [Google Scholar] [CrossRef]
- Valciņa; Pūle; Mališevs; Trofimova; Makarova; Konvisers; Bērziņš; Krūmiņa Co-Occurrence of Free-Living Amoeba and Legionella in Drinking Water Supply Systems. Medicina 2019, 55, 492. [CrossRef]
- Cabinet of Ministers of Latvia MK Nr. 906, 2010.28.09. Rules of Sanitary Maintenance of the Residential House. Available online: https://likumi.lv/ta/id/218830-dzivojamas-majas-sanitaras-apkopes-noteikumi (accessed on 11 May 2023).
- Valciņa, O.; Pūle, D.; Lucenko, I.; Krastiņa, D.; Šteingolde, Ž.; Krūmiņa, A.; Bērziņš, A. Legionella pneumophila Seropositivity-Associated Factors in Latvian Blood Donors. Int. J. Environ. Res. Public Health 2015, 13, 58. [Google Scholar] [CrossRef]
- Bešić, A.; Karakaš, S.; Obradović, Z.; Mušović, A.; Hrapović, E. Travel-Related Epidemiological Studies of Legionellosis in Federation of Bosnia and Herzegovina. Health Technol. 2021, 11, 971–979. [Google Scholar] [CrossRef]
- Raphael, B.H.; Huynh, T.; Brown, E.; Smith, J.C.; Ruberto, I.; Getsinger, L.; White, S.; Winchell, J.M. Culture of Clinical Specimens Reveals Extensive Diversity of Legionella pneumophila Strains in Arizona. mSphere 2019, 4, e00649-18. [Google Scholar] [CrossRef]
- Jiang, L.; Zhao, S.; Cai, X.; Mu, D.; Zhang, X.; Kang, J.; Zhao, L.; Chen, Y. Sequence-Based Typing of Clinical and Environmental Legionella pneumophila Isolates in Shenyang, China. Enferm. Infecc. Y Microbiol. Clin. (Engl. Ed.) 2021, 39, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Keše, D.; Obreza, A.; Rojko, T.; Kišek, T.C. Legionella pneumophila—Epidemiology and Characterization of Clinical Isolates, Slovenia, 2006–2020. Diagnostics 2021, 11, 1201. [Google Scholar] [CrossRef]
- Lévesque, S.; Lalancette, C.; Bernard, K.; Pacheco, A.L.; Dion, R.; Longtin, J.; Tremblay, C. Molecular Typing of Legionella pneumophila Isolates in the Province of Quebec from 2005 to 2015. PLoS ONE 2016, 11, e0163818. [Google Scholar] [CrossRef]
- Best, A.; Abu Kwaik, Y. Evolution of the Arsenal of Legionella pneumophila Effectors to Modulate Protist Hosts. mBio 2018, 9, e01313-18. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; O’Connor, T.J. Beyond Paralogs: The Multiple Layers of Redundancy in Bacterial Pathogenesis. Front. Cell. Infect. Microbiol. 2017, 7, 467. [Google Scholar] [CrossRef]
- Ridenour, D.A.; Cirillo, S.L.G.; Feng, S.; Samrakandi, M.M.; Cirillo, J.D. Identification of a Gene That Affects the Efficiency of Host Cell Infection by Legionella pneumophila in a Temperature-Dependent Fashion. Infect. Immun. 2003, 71, 6256–6263. [Google Scholar] [CrossRef]
- Sawczyn-Domańska, A. Detection of Legionella spp. And Occurrence of Virulence Genes: Lvh, RtxA and EnhC in Water Samples from Artificial Water Systems. Ann. Agric. Environ. Med. 2021, 28, 617–620. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.Z.; Liao, H.Y.; Luo, L.Z.; He, S.S.; Qin, T.; Zhou, H.J.; Li, H.X.; Chen, D.L.; Chen, J.P. An Investigation on the Molecular Characteristics and Intracellular Growth Ability among Environmental and Clinical Isolates of Legionella pneumophila in Sichuan Province, China. Biomed. Environ. Sci. 2019, 32, 520–530. [Google Scholar] [CrossRef] [PubMed]
- D’Auria, G.; Jiménez, N.; Peris-Bondia, F.; Pelaz, C.; Latorre, A.; Moya, A. Virulence Factor Rtx in Legionella pneumophila, Evidence Suggesting It Is a Modular Multifunctional Protein. BMC Genom. 2008, 9, 14. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| L. pneumophila Serogroup | No. of Isolates from Water Samples/ No. of Municipalities of Origin | No. of Selected Isolates/ No. of Municipalities of Origin (%) |
|---|---|---|
| SG 1 | 19/4 | 9/3 (47/75) |
| SG 2 | 108/14 | 22/11 (20/79) |
| SG 3 | 55/10 | 22/7 (40/70) |
| SG 6 | 4/2 | 2/2 (50/100) |
| SG 8 | 1/1 | 1/1 (100/100) |
| SG 9 | 4/1 | 4/1 (100/100) |
| Gene | Virulence Factor | Roles | |
|---|---|---|---|
| Bacterial surface structures | enhC | EnhC | Enhanced entry, trafficking of Legionella-containing vacuole [32] |
| htpB | Hsp60 | Attachment, modulation of invasion [33] | |
| mip | MIP | Penetration of the lung epithelial barrier [34] | |
| omp28 | MOMP | Mediating phagocytosis [35] | |
| T4SS effectors | mavC | MavC | Inhibiting host immunity [36] |
| legK1 | LegK1 | Activation of NF-kB [37] | |
| sidJ | SidJ | Calmodulin-activated glutamylase for SidE [38] | |
| lvhD4 | VirD4 | Coupling protein, reversing virulence defects [39] | |
| lpnE | LpnE | Entry into macrophages and epithelial cells; manipulate host cell trafficking [40] | |
| T2SS effectors | lsp | Lsp | Transport proteins from the periplasm to the extracellular space [41] |
| T1SS effectors | rtxA | RtxA | Ensures adherence and entry into host and enhances replication, cytotoxicity, and pore forming [42] |
| Buildings with Known LD Cases | Buildings without Known LD Cases | Total | ||||
|---|---|---|---|---|---|---|
| Samples Tested/Positive Samples (%) | Average Water Temperature, °C | Samples Tested/Positive Samples (%) | Average Water Temperature, °C | Samples Tested/Positive Samples (%) | Average Water Temperature, °C | |
| Cold water | 120/22 (18.3%) | 12.9 ± 0.4 | 44/19 (43.2%) | 15.6 ± 0.7 | 164/41 (25%) | 13.5 ± 0.3 |
| Hot water | 242/107 (44.2%) | 52.1 ± 0.4 | 86/49 (57.0%) | 45.8 ± 1.1 | 328/156 (47.6%) | 50.7 ± 0.4 |
| Total | 362/129 (35.6%) | — | 130/68 (52.3%) | — | 492/197 (40.0%) | — |
| Serogroup | No. of L. pneumophila Isolates (%) | ||
|---|---|---|---|
| Buildings with Known LD Cases | Buildings without Known LD Cases | Total | |
| SG 1 | 15 (11.6%) | 4 (6.0%) | 19 (9.7%) |
| SG 2 | 69 (53.5%) | 39 (58.2%) | 108 (55.1%) |
| SG 3 | 35 (27.1%) | 20 (29.9%) | 55 (28.1%) |
| SG 6 | 3 (2.3%) | 1 (1.5%) | 4 (2.0%) |
| SG 8 | 1 (0.8%) | — | 1 (0.5%) |
| SG 9 | 2 (1.6%) | 2 (3.0%) | 4 (2.0%) |
| SG 3, SG 2 | 3 (2.3%) | — | 3 (1.5%) |
| SG 3, SG 1 | — | 1 (1.5%) | 1 (0.5%) |
| SG 3, SG 9 | 1 (0.8%) | — | 1 (0.5%) |
| Serogroup | Levels of L. pneumophila Colonization (Min–Max (Average)), CFU/L | ||
|---|---|---|---|
| Buildings with Known LD Cases | Buildings without Known LD Cases | p-Value | |
| SG 1 | 50–4.0 × 103 (8.4 × 102) | 4.0 × 102–5.5 × 103 (2.6 × 103) | 0.056 |
| SG 2 | 50–1.3 × 103 (1.5 × 103) | 50–6.4 × 103 (2.1 × 103) | 0.158 |
| SG 3 | 50–1.7 × 104 (2.8 × 103) | 1.0 × 102–3.9 × 103 (8.2 × 102) | 0.033 |
| Total | 50–1.7 × 104 (2.0 × 103) | 50–6.4 × 103 (1.7 × 103) | 0.574 |
| Isolate Id | Year of Sampling | Municipality | SG | Linked with LD Cases | Allelic Profile | Sequence Type | Number of Observed Virulence Genes |
|---|---|---|---|---|---|---|---|
| Leg-1-2016 | 2016 | Krāslava | 6 | Yes | 7,6,17,3,50,11,9 | 1987 | 377 |
| Leg-2-2016 | 2016 | Rīga | 3 | Yes | 2,10,3,28,9,4,13 | 87 | 372 |
| Leg-3-2016 | 2016 | Valmiera | 8 | Yes | 2,10,3,28,9,4,207 | 1362 | 372 |
| Leg-4-2017 | 2017 | Rīga | 9 | Yes | 2,10,24,3,9,4,6 | 2192 | 370 |
| Leg-5-2017 | 2017 | Rīga | 9 | Yes | 10,22,7,28,16,18,6 | 337 | 358 |
| Leg-6-2017 | 2017 | Liepāja | 2 | Yes | 7,10,17,6,9,11,9 | New-ST-8 | 378 |
| Leg-7-2017 | 2017 | Rīga | 2 | Yes | 2,10,15,28,9,4,13 | 338 | 375 |
| Leg-8-2017 | 2017 | Bauska | 3 | Yes | 2,10,15,28,9,4,13 | 338 | 378 |
| Leg-9-2017 | 2017 | Rīga | 2 | Yes | 7,6,17,3,50,11,9 | 1987 | 378 |
| Leg-10-2017 | 2017 | Rīga | 3 | Yes | 7,10,17,28,17,11,9 | 738 | 376 |
| Leg-11-2018 | 2018 | Rīga | 9 | Yes | 2,10,15,28,9,4,13 | 338 | 378 |
| Leg-12-2018 | 2018 | Aizkraukle | 3 | No | 2,10,15,28,9,4,13 | 338 | 378 |
| Leg-13-2018 | 2018 | Liepāja | 2 | Yes | 2,32,20,38,34,35,219 | 2581 | 344 |
| Leg-14-2018 | 2018 | Rīga | 1 | Yes | 6,10,19,3,98,4, novel neuA allele | New-ST-7 | 374 |
| Leg-15-2018 | 2018 | Jūrmala | 6 | Yes | 2,10,3,3,9,4,6 | 366 | 374 |
| Leg-16-2018 | 2018 | Rīga | 1 | No | 6,10,19,3,19,4,11 | 345 | 374 |
| Leg-17-2018 | 2018 | Jūrmala | 3 | No | 2,10,1,3,9,4,3 | 787 | 372 |
| Leg-18-2018 | 2018 | Rīga | 1 | No | 2,10,1,3,9,4,3 | 787 | 372 |
| Leg-19-2018 | 2018 | Rīga | 2 | No | 2,10,24,28,4,4,207 | 1354 | 312 |
| Leg-20-2018 | 2018 | Rīga | 3 | No | 2,10,3,3,50,4,3 | New-ST-1 | 372 |
| Leg-21-2019 | 2019 | Rīga | 2 | Yes | 2,10,21,28,9,4,6 | New-ST-2 | 374 |
| Leg-22-2019 | 2019 | Rīga | 1 | Yes | 2,10,1,3,9,4,6 | 1939 | 374 |
| Leg-23-2019 | 2019 | Gulbene | 2 | Yes | 3,4,1,6,35,9,220 | 2059 | 415 |
| Leg-24-2019 | 2019 | Rīga | 3 | No | 3,13,1,28,14,9,13 | 1104 | 397 |
| Leg-25-2019 | 2019 | Rīga | 2 | No | 3,10,1,3,14,9,11 | 9 | 396 |
| Leg-26-2019 | 2019 | Rīga | 3 | No | 2,10,19,28,19,4,3 | 2207 | 360 |
| Leg-27-2019 | 2019 | Rīga | 3 | No | 3,10,1,28,14,9,13 | 93 | 415 |
| Leg-28-2019 | 2019 | Rīga | 1 | No | 2,10,17,3,9,4,6 | New-ST-3 | 373 |
| Leg-29-2019 | 2019 | Madona | 3 | Yes | 6,10,19,28,19,4,9 | 328 | 375 |
| Leg-30-2019 | 2019 | Daugavpils | 2 | Yes | 2,10,17,3,9,4,9 | New-ST-4 | 369 |
| Leg-31-2019 | 2019 | Ādaži | 2 | Yes | 3,10,1,28,1,9,3 | 242 | 415 |
| Leg-32-2019 | 2019 | Salaspils | 2 | Yes | 10,22,7,28,16,18,8 | 2002 | 363 |
| Leg-33-2019 | 2019 | Ogre | 2 | Yes | 10,22,7,28,16,18,8 | 2002 | 357 |
| Leg-34-2019 | 2019 | Rīga | 2 | No | 7,10,17,6,17,11,9 | New-ST-9 | 377 |
| Leg-35-2019 | 2019 | Rīga | 3 | No | 3,6,1,6,14,11,9 | 114 | 415 |
| Leg-36-2019 | 2019 | Rīga | 3 | No | 2,10,3,28,9,4,13 | 87 | 373 |
| Leg-37-2019 | 2019 | Rīga | 9 | No | 2,10,3,3,9,4,207 | 1860 | 371 |
| Leg-38-2019 | 2019 | Krāslava | 2 | Yes | 2,10,15,28,9,4,13 | 338 | 378 |
| Leg-39-2020 | 2020 | Ludza | 2 | Yes | 2,10,3,28,9,4,207 | 1362 | 376 |
| Leg-40-2020 | 2020 | Rīga | 3 | Yes | 2,10,24,28,4,4,207 | 1354 | 328 |
| Leg-41-2020 | 2020 | Tukums | 1 | Yes | 2,10,3,10,9,4,11 | 170 | 375 |
| Leg-42-2020 | 2020 | Rīga | 3 | No | 2,10,15,28,9,4,13 | 338 | 371 |
| Leg-43-2020 | 2020 | Olaine | 1 | Yes | 2,10,3,3,9,4,6 | 366 | 379 |
| Leg-44-2020 | 2020 | Rīga | 3 | Yes | 2,22,3,28,50,4,3 | New-ST-5 | 371 |
| Leg-45-2020 | 2020 | Sigulda | 3 | Yes | 2,10,3,28,9,4,3 | 728 | 373 |
| Leg-46-2021 | 2021 | Rīga | 3 | Yes | 3,13,1,28,14,9,13 | 1104 | 412 |
| Leg-47-2021 | 2021 | Rēzekne | 3 | Yes | 2,10,15,28,9,4,13 | 338 | 378 |
| Leg-48-2022 | 2022 | Liepāja | 2 | Yes | 2,10,21,28,9,4,6 | New-ST-2 | 372 |
| Leg-49-2022 | 2022 | Rīga | 1 | Yes | 6,10,19,3,17,4,11 | New-ST-7 | 374 |
| Leg-50-2022 | 2022 | Baloži | 2 | Yes | 2,10,21,28,9,4,13 | 646 | 375 |
| Leg-51-2022 | 2022 | Rīga | 2 | No | 2,10,3,3,50,4,6 | New-ST-6 | 372 |
| Leg-52-2022 | 2022 | Rīga | 2 | No | 10,22,7,28,16,18,8 | 2002 | 360 |
| Leg-53-2022 | 2022 | Rīga | 1 | No | 7,6,17,3,50,4,9 | New-ST-10 | 390 |
| Leg-54-2022 | 2022 | Rīga | 2 | No | 2,10,15,28,9,4,13 | 338 | 375 |
| Leg-55-2022 | 2022 | Rīga | 3 | No | 2,10,15,28,9,4,13 | 338 | 375 |
| Leg-56-2022 | 2022 | Rīga | 3 | No | 2,32,20,38,34,35,219 | 2581 | 344 |
| Leg-57-2022 | 2022 | Rīga | 2 | No | 2,10,15,28,9,4,13 | 338 | 375 |
| Leg-58-2022 | 2022 | Rīga | 1 | No | 2,10,3,3,9,4,6 | 366 | 375 |
| Virulence Gene | No. of Positive/No. of Sequenced Isolates | Relative Frequency, % | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Overall | Isolates from Buildings Linked to LD Cases N = 34 | Isolates from Buildings Not Linked to LD Cases N = 24 | SG 1 Isolates N = 10 | SG 2 Isolates N = 21 | SG 3 Isolates N = 20 | SG 6 Isolates N = 2 | SG 8 Isolates N = 1 | SG 9 Isolates N = 4 | ||
| enhC | 58/58 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| htpB | 58/58 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| omp28 | 58/58 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| mip | 58/58 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| mavC | 54/58 | 93 | 94 | 92 | 100 | 90 | 90 | 100 | 100 | 100 |
| legK1 | 57/58 | 98 | 100 | 96 | 100 | 95 | 100 | 100 | 100 | 100 |
| sidJ | 54/58 | 93 | 94 | 92 | 100 | 100 | 95 | 100 | 100 | 25 |
| lvhD4 | 48/58 | 83 | 85 | 79 | 100 | 76 | 80 | 50 | 100 | 100 |
| lpnE | 58/58 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| lspC | 58/58 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| rtxA AA100 | 23/58 | 40 | 32 | 50 | 10 | 38 | 55 | 0 | 100 | 50 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valciņa, O.; Pūle, D.; Ķibilds, J.; Labecka, L.; Terentjeva, M.; Krūmiņa, A.; Bērziņš, A. Evaluation of Genetic Diversity and Virulence Potential of Legionella pneumophila Isolated from Water Supply Systems of Residential Buildings in Latvia. Pathogens 2023, 12, 884. https://doi.org/10.3390/pathogens12070884
Valciņa O, Pūle D, Ķibilds J, Labecka L, Terentjeva M, Krūmiņa A, Bērziņš A. Evaluation of Genetic Diversity and Virulence Potential of Legionella pneumophila Isolated from Water Supply Systems of Residential Buildings in Latvia. Pathogens. 2023; 12(7):884. https://doi.org/10.3390/pathogens12070884
Chicago/Turabian StyleValciņa, Olga, Daina Pūle, Juris Ķibilds, Linda Labecka, Margarita Terentjeva, Angelika Krūmiņa, and Aivars Bērziņš. 2023. "Evaluation of Genetic Diversity and Virulence Potential of Legionella pneumophila Isolated from Water Supply Systems of Residential Buildings in Latvia" Pathogens 12, no. 7: 884. https://doi.org/10.3390/pathogens12070884
APA StyleValciņa, O., Pūle, D., Ķibilds, J., Labecka, L., Terentjeva, M., Krūmiņa, A., & Bērziņš, A. (2023). Evaluation of Genetic Diversity and Virulence Potential of Legionella pneumophila Isolated from Water Supply Systems of Residential Buildings in Latvia. Pathogens, 12(7), 884. https://doi.org/10.3390/pathogens12070884