
Clostridium perfringens—Opportunistic Foodborne Pathogen, Its Diversity and Epidemiological Significance
 , , and
, , and
Abstract
1. Introduction
2. Clostridium perfringens Organism and Its Toxins
- CPA (α-toxin) has enzyme activity able to degrade cell membranes by breaking down phosphatidylcholine and sphingomyelin and inhibiting the maturation of neutrophils as well as activating arachidonic acid metabolism. This then leads to vasoconstriction and platelet aggregation [30]. Consequently, the CPA toxin causes impairment of the innate immune response [31,32]. It is an extremely important factor in gas gangrene [5].
- ETX (ε-toxin) is the main aetiological factor of haemorrhagic enteritis and enterotoxaemia in sheep [34]. It becomes active after the action of enteric proteases and causes an increase in intestinal permeability. It is a cause of perivascular oedema with rapid cellular oedema. Moreover, it is able to accumulate in the kidneys and brain. The mechanism of the mentioned activity has not been elucidated until now [35,36].
- ITX (ι-toxin) is known as a binary toxin that is able to produce two distinct proteins: Ia and Ib. The mentioned Ib protein binds to a cell surface receptor and associates with Ia. The complex is endocytosed. The membrane channel created by Ib enables Ia to enter the cytosol and consequently causes depolymerisation of the actin cytoskeleton via ADP-ribosylation [37].
- CPE (enterotoxin) is also a pore-forming toxin able to bind to claudin receptors on the cell surface [38]. It is able to form a hexamer complex, playing an important role in calcium influx. The mentioned influx is dose-dependent and is one cause of activation of calpain and, consequently, cell death. The activity of CPE is the main cause of food-poisoning and non-foodborne diarrhoea [36].
- NetB (necrotic enteritis B-like toxin) is identified in avian necrotic enteritis, causing pore formation. NetB has been shown to have a 38% sequence similarity with CPB [39].

{kind=link}
| Toxin | Location | Biological Activities | Actions |
|---|---|---|---|
| CPA | Chromosomal | Necrotising, haemolytic, contraction of smooth muscle | Phospholipase C, activation of host cell signalling |
| CPB | Plasmid | Dermonecrosis, enterotoxic, oedema | Formation of pores |
| ETX | Plasmid | Dermonecrosis, oedema, contraction of smooth muscle | Formation of pores |
| ITX | Plasmid | Necrotising | ADP-ribosylating activities |
| PFO | Chromosomal | Necrotising | Formation of pores |
| CPE | Chromosomal/plasmid | Enterotoxic, erythema | Formation of pores |
| NetB | Plasmid | haemolytic | Formation of pores |
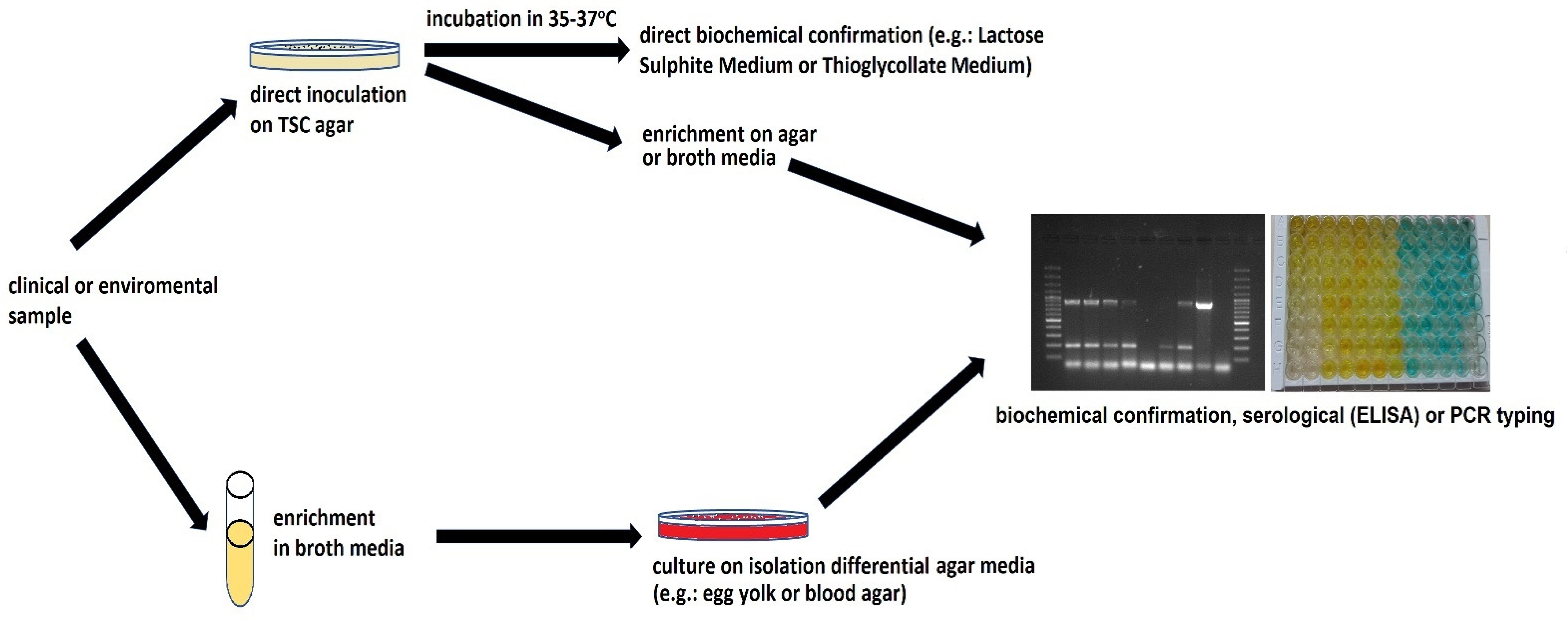
3. Clostridium perfringens—Epidemiology
4. Clostridium perfringens—Molecular Diversity
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lucey, B.P.; Hutchins, G.M.; William, H. William H. Welch, MD, and the Discovery of Bacillus welchii. Arch. Pathol. Lab. Med. 2004, 128, 1193–1195. [Google Scholar] [CrossRef] [PubMed]
- Welch, W.H.; Nutfall, G.H.F. A Gas-Producing Bacillus (Bacillus Aërogenes Capsulatus Nov.Spec.) Capable of Rapid Development in the Blood-Vessels after Death. Available online: https://wellcomecollection.org/works/upj2scka (accessed on 17 April 2023).
- Welch, W.H.; Flexner, S. Observations Concerning the Bacillus Aerogenes Capsulatus. J. Exp. Med. 1896, 1, 5–45. [Google Scholar] [CrossRef] [PubMed]
- Callender, G.R.; Coupal, J.F. Pathology of the Acute Respiratory Diseases, and of Gas Gangrene Following War Wounds; U.S. Government Printing Office: Washington, DC, USA, 1929.
- Rood, J.I.; Adams, V.; Lacey, J.; Lyras, D.; McClane, B.A.; Melville, S.B.; Moore, R.J.; Popoff, M.R.; Sarker, M.R.; Songer, J.G.; et al. Expansion of the Clostridium perfringens Toxin-Based Typing Scheme. Anaerobe 2018, 53, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Winslow, C.E.; Broadhurst, J.; Buchanan, R.E.; Krumwiede, C.; Rogers, L.A.; Smith, G.H. The Families and Genera of the Bacteria: Final Report of the Committee of the Society of American Bacteriologists on Characterization and Classification of Bacterial Types. J. Bacteriol. 1920, 5, 191–229. [Google Scholar] [CrossRef]
- Lugli, G.A.; Milani, C.; Mancabelli, L.; Turroni, F.; Ferrario, C.; Duranti, S.; van Sinderen, D.; Ventura, M. Ancient Bacteria of the Ötzi’s Microbiome: A Genomic Tale from the Copper Age. Microbiome 2017, 5, 5. [Google Scholar] [CrossRef]
- Jean, D.; Briolat, V.; Reysset, G. Oxidative Stress Response in Clostridium perfringens. Microbiol. Read. Engl. 2004, 150, 1649–1659. [Google Scholar] [CrossRef]
- Kiu, R.; Hall, L.J. An Update on the Human and Animal Enteric Pathogen Clostridium perfringens. Emerg. Microbes Infect. 2018, 7, 141. [Google Scholar] [CrossRef]
- Bosworth, T.J. On a New Type of Toxin Produced by Clostridium welchii. J. Comp. Pathol. Ther. 1943, 53, 245–255. [Google Scholar] [CrossRef]
- Shrestha, A.; Uzal, F.A.; McClane, B.A. Enterotoxic Clostridia: Clostridium perfringens Enteric Diseases. Microbiol. Spectr. 2018, 6. [Google Scholar] [CrossRef]
- Stiles, B.; Barth, G.; Barth, H.; Popoff, M. Clostridium perfringens Epsilon Toxin: A Malevolent Molecule for Animals and Man? Toxins 2013, 5, 2138–2160. [Google Scholar] [CrossRef]
- Selim, A.M.; Elhaig, M.M.; Zakaria, I.; Ali, A. Bacteriological and Molecular Studies of Clostridium perfringens Infections in Newly Born Calves. Trop. Anim. Health Prod. 2017, 49, 201–205. [Google Scholar] [CrossRef]
- Li, J.; McClane, B.A. Further Comparison of Temperature Effects on Growth and Survival of Clostridium perfringens Type A Isolates Carrying a Chromosomal or Plasmid-Borne Enterotoxin Gene. Appl. Environ. Microbiol. 2006, 72, 4561–4568. [Google Scholar] [CrossRef] [PubMed]
- Novak, J.S.; Juneja, V.K.; McClane, B.A. An Ultrastructural Comparison of Spores from Various Strains of Clostridium perfringens and Correlations with Heat Resistance Parameters. Int. J. Food Microbiol. 2003, 86, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Paredes-Sabja, D.; Sarker, M.R.; McClane, B.A. Further Characterization of Clostridium perfringens Small Acid Soluble Protein-4 (Ssp4) Properties and Expression. PLoS ONE 2009, 4, e6249. [Google Scholar] [CrossRef]
- Li, J.; McClane, B.A. A Novel Small Acid Soluble Protein Variant Is Important for Spore Resistance of Most Clostridium perfringens Food Poisoning Isolates. PLoS Pathog. 2008, 4, e1000056. [Google Scholar] [CrossRef] [PubMed]
- Dave, G.A. A Rapid Qualitative Assay for Detection of Clostridium perfringens in Canned Food Products. Acta Biochim. Pol. 2017, 64, 207–213. [Google Scholar] [CrossRef]
- Allaart, J.G.; van Asten, A.J.A.M.; Gröne, A. Predisposing Factors and Prevention of Clostridium perfringens-Associated Enteritis. Comp. Immunol. Microbiol. Infect. Dis. 2013, 36, 449–464. [Google Scholar] [CrossRef]
- Horwitz, W.; Latimer, G.W. Official Methods of Analysis of AOAC International, 18th ed.; AOAC International: Gaithersburg, MD, USA, 2010; ISBN 978-0-935584-80-6. [Google Scholar]
- Varga, J.J.; Nguyen, V.; O’Brien, D.K.; Rodgers, K.; Walker, R.A.; Melville, S.B. Type IV Pili-Dependent Gliding Motility in the Gram-Positive Pathogen Clostridium perfringens and Other Clostridia. Mol. Microbiol. 2006, 62, 680–694. [Google Scholar] [CrossRef]
- Fohler, S.; Klein, G.; Hoedemaker, M.; Scheu, T.; Seyboldt, C.; Campe, A.; Jensen, K.C.; Abdulmawjood, A. Diversity of Clostridium perfringens Toxin-Genotypes from Dairy Farms. BMC Microbiol. 2016, 16, 199. [Google Scholar] [CrossRef] [PubMed]
- Chukwu, E.E.; Nwaokorie, F.O.; Coker, A.O.; Avila-Campos, M.J.; Solis, R.L.; Llanco, L.A.; Ogunsola, F.T. Detection of Toxigenic Clostridium perfringens and Clostridium Botulinum from Food Sold in Lagos, Nigeria. Anaerobe 2016, 42, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Wimsatt, J.C.; Harmon, S.M.; Shah, D.B. Detection of Clostridium perfringens Enterotoxin in Stool Specimens and Culture Supernatants by Enzyme-Linked Immunosorbent Assay. Diagn. Microbiol. Infect. Dis. 1986, 4, 307–313. [Google Scholar] [CrossRef] [PubMed]
- McClane, B.A.; Snyder, J.T. Development and Preliminary Evaluation of a Slide Latex Agglutination Assay for Detection of Clostridium perfringens Type A Enterotoxin. J. Immunol. Methods 1987, 100, 131–136. [Google Scholar] [CrossRef]
- Harmon, S.M.; Kautter, D.A. Evaluation of a Reversed Passive Latex Agglutination Test Kit for Clostridium perfringens Enterotoxin. J. Food Prot. 1986, 49, 523–525. [Google Scholar] [CrossRef] [PubMed]
- Harmon, S.M.; Kautter, D.A. Improved Media for Sporulation and Enterotoxin Production by Clostridium perfringens. J. Food Prot. 1986, 49, 706–711. [Google Scholar] [CrossRef]
- Labbe, R.G.; Rey, D.K. Raffinose Increases Sporulation and Enterotoxin Production by Clostridium perfringens Type A. Appl. Environ. Microbiol. 1979, 37, 1196–1200. [Google Scholar] [CrossRef] [PubMed]
- Yao, P.; Annamaraju, P. Clostridium perfringens. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Titball, R.W.; Naylor, C.E.; Basak, A.K. The Clostridium perfringens Alpha-Toxin. Anaerobe 1999, 5, 51–64. [Google Scholar] [CrossRef]
- Takehara, M.; Takagishi, T.; Seike, S.; Ohtani, K.; Kobayashi, K.; Miyamoto, K.; Shimizu, T.; Nagahama, M. Clostridium perfringens α-Toxin Impairs Innate Immunity via Inhibition of Neutrophil Differentiation. Sci. Rep. 2016, 6, 28192. [Google Scholar] [CrossRef]
- Fernandez-Miyakawa, M.E.; Redondo, L.M. Role of Clostridium perfringens Alpha, Beta, Epsilon and Iota Toxins in Enterotoxemia of Monogastrics and Ruminants. In Microbial Toxins; Gopalakrishnakone, P., Stiles, B., Alape-Girón, A., Dubreuil, J.D., Mandal, M., Eds.; Toxinology; Springer: Dordrecht, The Netherlands, 2016; pp. 1–26. ISBN 978-94-007-6725-6. [Google Scholar]
- Nagahama, M.; Ochi, S.; Oda, M.; Miyamoto, K.; Takehara, M.; Kobayashi, K. Recent Insights into Clostridium perfringens Beta-Toxin. Toxins 2015, 7, 396–406. [Google Scholar] [CrossRef]
- Garcia, J.P.; Adams, V.; Beingesser, J.; Hughes, M.L.; Poon, R.; Lyras, D.; Hill, A.; McClane, B.A.; Rood, J.I.; Uzal, F.A. Epsilon Toxin Is Essential for the Virulence of Clostridium perfringens Type D Infection in Sheep, Goats, and Mice. Infect. Immun. 2013, 81, 2405–2414. [Google Scholar] [CrossRef]
- Popoff, M.R. Epsilon Toxin: A Fascinating Pore-Forming Toxin. FEBS J. 2011, 278, 4602–4615. [Google Scholar] [CrossRef]
- Navarro, M.A.; McClane, B.A.; Uzal, F.A. Mechanisms of Action and Cell Death Associated with Clostridium perfringens Toxins. Toxins 2018, 10, 212. [Google Scholar] [CrossRef] [PubMed]
- Takehara, M.; Takagishi, T.; Seike, S.; Oda, M.; Sakaguchi, Y.; Hisatsune, J.; Ochi, S.; Kobayashi, K.; Nagahama, M. Cellular Entry of Clostridium perfringens Iota-Toxin and Clostridium Botulinum C2 Toxin. Toxins 2017, 9, 247. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, A.; Uzal, F.A.; McClane, B.A. The Interaction of Clostridium perfringens Enterotoxin with Receptor Claudins. Anaerobe 2016, 41, 18–26. [Google Scholar] [CrossRef]
- Keyburn, A.L.; Boyce, J.D.; Vaz, P.; Bannam, T.L.; Ford, M.E.; Parker, D.; Di Rubbo, A.; Rood, J.I.; Moore, R.J. NetB, a New Toxin That Is Associated with Avian Necrotic Enteritis Caused by Clostridium perfringens. PLoS Pathog. 2008, 4, e26. [Google Scholar] [CrossRef] [PubMed]
- Awad, M.M.; Ellemor, D.M.; Boyd, R.L.; Emmins, J.J.; Rood, J.I. Synergistic Effects of Alpha-Toxin and Perfringolysin O in Clostridium perfringens-Mediated Gas Gangrene. Infect. Immun. 2001, 69, 7904–7910. [Google Scholar] [CrossRef]
- Li, J.; Adams, V.; Bannam, T.L.; Miyamoto, K.; Garcia, J.P.; Uzal, F.A.; Rood, J.I.; McClane, B.A. Toxin Plasmids of Clostridium perfringens. Microbiol. Mol. Biol. Rev. MMBR 2013, 77, 208–233. [Google Scholar] [CrossRef]
- McDonel, J.L. Clostridium perfringens Toxins (Type A, B, C, D, E). Pharmacol. Ther. 1980, 10, 617–655. [Google Scholar] [CrossRef]
- Yan, X.-X.; Porter, C.J.; Hardy, S.P.; Steer, D.; Smith, A.I.; Quinsey, N.S.; Hughes, V.; Cheung, J.K.; Keyburn, A.L.; Kaldhusdal, M.; et al. Structural and Functional Analysis of the Pore-Forming Toxin NetB from Clostridium perfringens. mBio 2013, 4, e00019-13. [Google Scholar] [CrossRef]
- Hatheway, C.L. Toxigenic Clostridia. Clin. Microbiol. Rev. 1990, 3, 66–98. [Google Scholar] [CrossRef]
- Awad, M.M.; Bryant, A.E.; Stevens, D.L.; Rood, J.I. Virulence Studies on Chromosomal Alpha-Toxin and Theta-Toxin Mutants Constructed by Allelic Exchange Provide Genetic Evidence for the Essential Role of Alpha-Toxin in Clostridium perfringens-Mediated Gas Gangrene. Mol. Microbiol. 1995, 15, 191–202. [Google Scholar] [CrossRef]
- Mehdizadeh Gohari, I.; Navarro, A.M.; Li, J.; Shrestha, A.; Uzal, F.; McClane, A.B. Pathogenicity and Virulence of Clostridium perfringens. Virulence 2021, 12, 723–753. [Google Scholar] [CrossRef] [PubMed]
- Stevens, D.L.; Aldape, M.J.; Bryant, A.E. Life-Threatening Clostridial Infections. Anaerobe 2012, 18, 254–259. [Google Scholar] [CrossRef] [PubMed]
- Buboltz, J.B.; Murphy-Lavoie, H.M. Gas Gangrene. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Shindo, Y.; Dobashi, Y.; Sakai, T.; Monma, C.; Miyatani, H.; Yoshida, Y. Epidemiological and Pathobiological Profiles of Clostridium perfringens Infections: Review of Consecutive Series of 33 Cases over a 13-Year Period. Int. J. Clin. Exp. Pathol. 2015, 8, 569–577. [Google Scholar] [PubMed]
- Bryant, A.E.; Stevens, D.L. Clostridial Myonecrosis: New Insights in Pathogenesis and Management. Curr. Infect. Dis. Rep. 2010, 12, 383–391. [Google Scholar] [CrossRef]
- Uzal, F.A. Diseases Produced by Clostridium perfringens Type A in Mammalian Species. In Clostridial Diseases of Animals; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2016; pp. 107–116. ISBN 978-1-118-72829-1. [Google Scholar]
- Mehdizadeh Gohari, I.; Parreira, V.R.; Nowell, V.J.; Nicholson, V.M.; Oliphant, K.; Prescott, J.F. A Novel Pore-Forming Toxin in Type A Clostridium perfringens Is Associated with Both Fatal Canine Hemorrhagic Gastroenteritis and Fatal Foal Necrotizing Enterocolitis. PLoS ONE 2015, 10, e0122684. [Google Scholar] [CrossRef]
- Munday, J.S.; Bentall, H.; Aberdein, D.; Navarro, M.; Uzal, F.A.; Brown, S. Death of a Neonatal Lamb Due to Clostridium perfringens Type B in New Zealand. N. Z. Vet. J. 2020, 68, 242–246. [Google Scholar] [CrossRef]
- Uzal, F.A.; Songer, J.G. Diagnosis of Clostridium perfringens Intestinal Infections in Sheep and Goats. J. Vet. Diagn. Investig. 2008, 20, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Uzal, F.A. Diagnosis of Clostridium perfringens Intestinal Infections in Sheep and Goats. Anaerobe 2004, 10, 135–143. [Google Scholar] [CrossRef]
- Wagley, S.; Bokori-Brown, M.; Morcrette, H.; Malaspina, A.; D’Arcy, C.; Gnanapavan, S.; Lewis, N.; Popoff, M.R.; Raciborska, D.; Nicholas, R.; et al. Evidence of Clostridium perfringens Epsilon Toxin Associated with Multiple Sclerosis. Mult. Scler. Houndmills Basingstoke Engl. 2019, 25, 653–660. [Google Scholar] [CrossRef]
- Ma, Y.; Sannino, D.; Linden, J.R.; Haigh, S.; Zhao, B.; Grigg, J.B.; Zumbo, P.; Dündar, F.; Butler, D.; Profaci, C.P.; et al. Epsilon Toxin–Producing Clostridium perfringens Colonize the Multiple Sclerosis Gut Microbiome Overcoming CNS Immune Privilege. Available online: https://www.jci.org/articles/view/163239/pdf (accessed on 8 May 2023).
- Diab, S.S.; Kinde, H.; Moore, J.; Shahriar, M.F.; Odani, J.; Anthenill, L.; Songer, G.; Uzal, F.A. Pathology of Clostridium perfringens Type C Enterotoxemia in Horses. Vet. Pathol. 2012, 49, 255–263. [Google Scholar] [CrossRef]
- Sayeed, S.; Uzal, F.A.; Fisher, D.J.; Saputo, J.; Vidal, J.E.; Chen, Y.; Gupta, P.; Rood, J.I.; McClane, B.A. Beta Toxin Is Essential for the Intestinal Virulence of Clostridium perfringens Type C Disease Isolate CN3685 in a Rabbit Ileal Loop Model. Mol. Microbiol. 2008, 67, 15–30. [Google Scholar] [CrossRef]
- Matsuda, T.; Okada, Y.; Inagi, E.; Tanabe, Y.; Shimizu, Y.; Nagashima, K.; Sakurai, J.; Nagahama, M.; Tanaka, S. Enteritis Necroticans “pigbel” in a Japanese Diabetic Adult. Pathol. Int. 2007, 57, 622–626. [Google Scholar] [CrossRef]
- Petrillo, T.M.; Beck-Sagué, C.M.; Songer, J.G.; Abramowsky, C.; Fortenberry, J.D.; Meacham, L.; Dean, A.G.; Lee, H.; Bueschel, D.M.; Nesheim, S.R. Enteritis Necroticans (Pigbel) in a Diabetic Child. N. Engl. J. Med. 2000, 342, 1250–1253. [Google Scholar] [CrossRef]
- Garcia, J.P.; Anderson, M.; Blanchard, P.; Mete, A.; Uzal, F.A. The Pathology of Enterotoxemia by Clostridium perfringens Type C in Calves. J. Vet. Diagn. Investig. 2013, 25, 438–442. [Google Scholar] [CrossRef]
- Kreft, B.; Dalhoff, K.; Sack, K. Necrotizing enterocolitis: A historical and current review. Med. Klin. Munich Ger. 2000, 95, 435–441. [Google Scholar] [CrossRef]
- Lawrence, G.; Cooke, R. Experimental Pigbel: The Production and Pathology of Necrotizing Enteritis Due to Clostridium welchii Type C in the Guinea-Pig. Br. J. Exp. Pathol. 1980, 61, 261–271. [Google Scholar]
- Kim, H.-Y.; Byun, J.-W.; Roh, I.-S.; Bae, Y.-C.; Lee, M.-H.; Kim, B.; Songer, J.G.; Jung, B.Y. First Isolation of Clostridium perfringens Type E from a Goat with Diarrhea. Anaerobe 2013, 22, 141–143. [Google Scholar] [CrossRef]
- Sarker, M.R.; Carman, R.J.; McClane, B.A. Inactivation of the Gene (Cpe) Encoding Clostridium perfringens Enterotoxin Eliminates the Ability of Two Cpe-Positive C. Perfringens Type A Human Gastrointestinal Disease Isolates to Affect Rabbit Ileal Loops. Mol. Microbiol. 1999, 33, 946–958. [Google Scholar] [CrossRef]
- Uzal, F.A.; Freedman, J.C.; Shrestha, A.; Theoret, J.R.; Garcia, J.; Awad, M.M.; Adams, V.; Moore, R.J.; Rood, J.I.; McClane, B.A. Towards an Understanding of the Role of Clostridium perfringens Toxins in Human and Animal Disease. Future Microbiol. 2014, 9, 361–377. [Google Scholar] [CrossRef]
- Collie, R.E.; McClane, B.A. Evidence That the Enterotoxin Gene Can Be Episomal in Clostridium perfringens Isolates Associated with Non-Food-Borne Human Gastrointestinal Diseases. J. Clin. Microbiol. 1998, 36, 30–36. [Google Scholar] [CrossRef]
- Lovland, A.; Kaldhusdal, M.; Redhead, K.; Skjerve, E.; Lillehaug, A. Maternal Vaccination against Subclinical Necrotic Enteritis in Broilers. Avian Pathol. 2004, 33, 81–90. [Google Scholar] [CrossRef]
- Cooper, K.K.; Songer, J.G.; Uzal, F.A. Diagnosing Clostridial Enteric Disease in Poultry. J. Vet. Diagn. Investig. 2013, 25, 314–327. [Google Scholar] [CrossRef]
- Beceiro, A.; Tomás, M.; Bou, G. Antimicrobial Resistance and Virulence: A Successful or Deleterious Association in the Bacterial World? Clin. Microbiol. Rev. 2013, 26, 185–230. [Google Scholar] [CrossRef] [PubMed]
- Wei, B.; Cha, S.-Y.; Zhang, J.-F.; Shang, K.; Park, H.-C.; Kang, J.; Lee, K.-J.; Kang, M.; Jang, H.-K. Antimicrobial Susceptibility and Association with Toxin Determinants in Clostridium perfringens Isolates from Chickens. Microorganisms 2020, 8, 1825. [Google Scholar] [CrossRef] [PubMed]
- Regassa, B.; Mohammed, M. Review on the Major Antimicrobial Resistance Bacterial Pathogen of Poultry. J. Dairy Vet. Sci. 2019, 12, 1–8. [Google Scholar] [CrossRef]
- The Risk of Development of Antimicrobial Resistance with the Use of Coccidiostats in Poultry Diets. VKM Rep. 2014.
- Fan, Y.-C.; Wang, C.-L.; Wang, C.; Chen, T.-C.; Chou, C.-H.; Tsai, H.-J. Incidence and Antimicrobial Susceptibility to Clostridium perfringens in Premarket Broilers in Taiwan. Avian Dis. 2016, 60, 444–449. [Google Scholar] [CrossRef]
- Slavić, Đ.; Boerlin, P.; Fabri, M.; Klotins, K.C.; Zoethout, J.K.; Weir, P.E.; Bateman, D. Antimicrobial Susceptibility of Clostridium perfringens Isolates of Bovine, Chicken, Porcine, and Turkey Origin from Ontario. Can. J. Vet. Res. 2011, 75, 89–97. [Google Scholar]
- Gulliver, E.; Adams, V.; Rossetto Marcelino, V.; Gould, J.; Rutten, E.; Powell, D.; Young, R.; D’Adamo, G.; Hemphill, J.; Solari, S.; et al. Extensive Genome Analysis Identifies Novel Plasmid Families in Clostridium perfringens. Microb. Genom. 2023, 9, 000995. [Google Scholar] [CrossRef]
- Abdel-Glil, M.Y.; Thomas, P.; Linde, J.; Busch, A.; Wieler, L.H.; Neubauer, H.; Seyboldt, C. Comparative in Silico Genome Analysis of Clostridium perfringens Unravels Stable Phylogroups with Different Genome Characteristics and Pathogenic Potential. Sci. Rep. 2021, 11, 6756. [Google Scholar] [CrossRef]
- Harris, S.R.; Robinson, C.; Steward, K.F.; Webb, K.S.; Paillot, R.; Parkhill, J.; Holden, M.T.G.; Waller, A.S. Genome Specialization and Decay of the Strangles Pathogen, Streptococcus equi, Is Driven by Persistent Infection. Genome Res. 2015, 25, 1360–1371. [Google Scholar] [CrossRef] [PubMed]

| Toxinotype | α-Toxin CPE | β-Toxins CPB | ε-Toxins ETX | ι-Toxins ITX | CPE Enterotoxin | NetB Necrotic Enteritis B-like Toxin | Diseases in Humans and Animals |
|---|---|---|---|---|---|---|---|
| Genes | cpa/plc | cpb | etx | iap/ibp/itx | cpe | netB | - |
| A | + | − | − | − | − | − | Gas gangrene in humans and animals; necrotic enteritis in fowls and piglets; human food poisoning and antibiotic-associated diarrhoea |
| B | + | + | + | − | − | − | Haemorrhagic enteritis in calves, foals and sheep; dysentery in lambs |
| C | + | + | − | − | ± | − | Enterotoxemia in sheep; necrotising enteritis in humans (pigbel), pigs, calves, goats, and foals |
| D | + | − | + | − | ± | − | Enterotoxemia in lambs (pulpy kidney disease), goats, and cattle |
| E | + | − | − | + | ± | − | Enterotoxemia in calves and lambs |
| F | + | − | − | − | + | − | Food and feed poisoning of humans and animals |
| G | + | − | − | − | − | + | Avian necrotic enteritis |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grenda, T.; Jarosz, A.; Sapała, M.; Grenda, A.; Patyra, E.; Kwiatek, K. Clostridium perfringens—Opportunistic Foodborne Pathogen, Its Diversity and Epidemiological Significance. Pathogens 2023, 12, 768. https://doi.org/10.3390/pathogens12060768
Grenda T, Jarosz A, Sapała M, Grenda A, Patyra E, Kwiatek K. Clostridium perfringens—Opportunistic Foodborne Pathogen, Its Diversity and Epidemiological Significance. Pathogens. 2023; 12(6):768. https://doi.org/10.3390/pathogens12060768
Chicago/Turabian StyleGrenda, Tomasz, Aleksandra Jarosz, Magdalena Sapała, Anna Grenda, Ewelina Patyra, and Krzysztof Kwiatek. 2023. "Clostridium perfringens—Opportunistic Foodborne Pathogen, Its Diversity and Epidemiological Significance" Pathogens 12, no. 6: 768. https://doi.org/10.3390/pathogens12060768
APA StyleGrenda, T., Jarosz, A., Sapała, M., Grenda, A., Patyra, E., & Kwiatek, K. (2023). Clostridium perfringens—Opportunistic Foodborne Pathogen, Its Diversity and Epidemiological Significance. Pathogens, 12(6), 768. https://doi.org/10.3390/pathogens12060768