


First Report of Swinepox in a Wild Boar in Italy: Pathologic and Molecular Findings
 , , , , ,
, , , , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Post-Mortem Examination
2.2. Histopathology
2.3. Molecular Analysis
3. Results
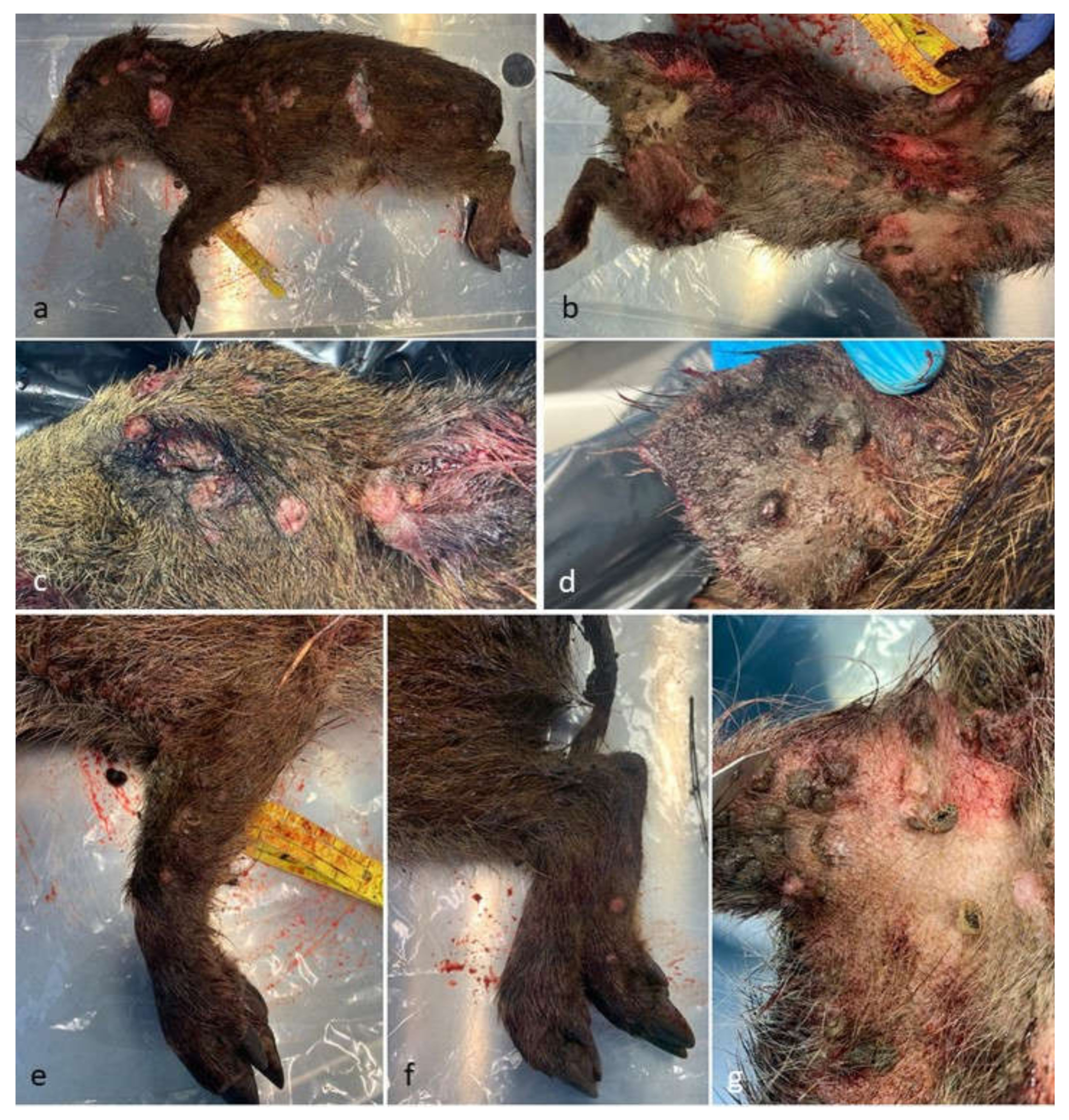
3.1. Post-Mortem Examination
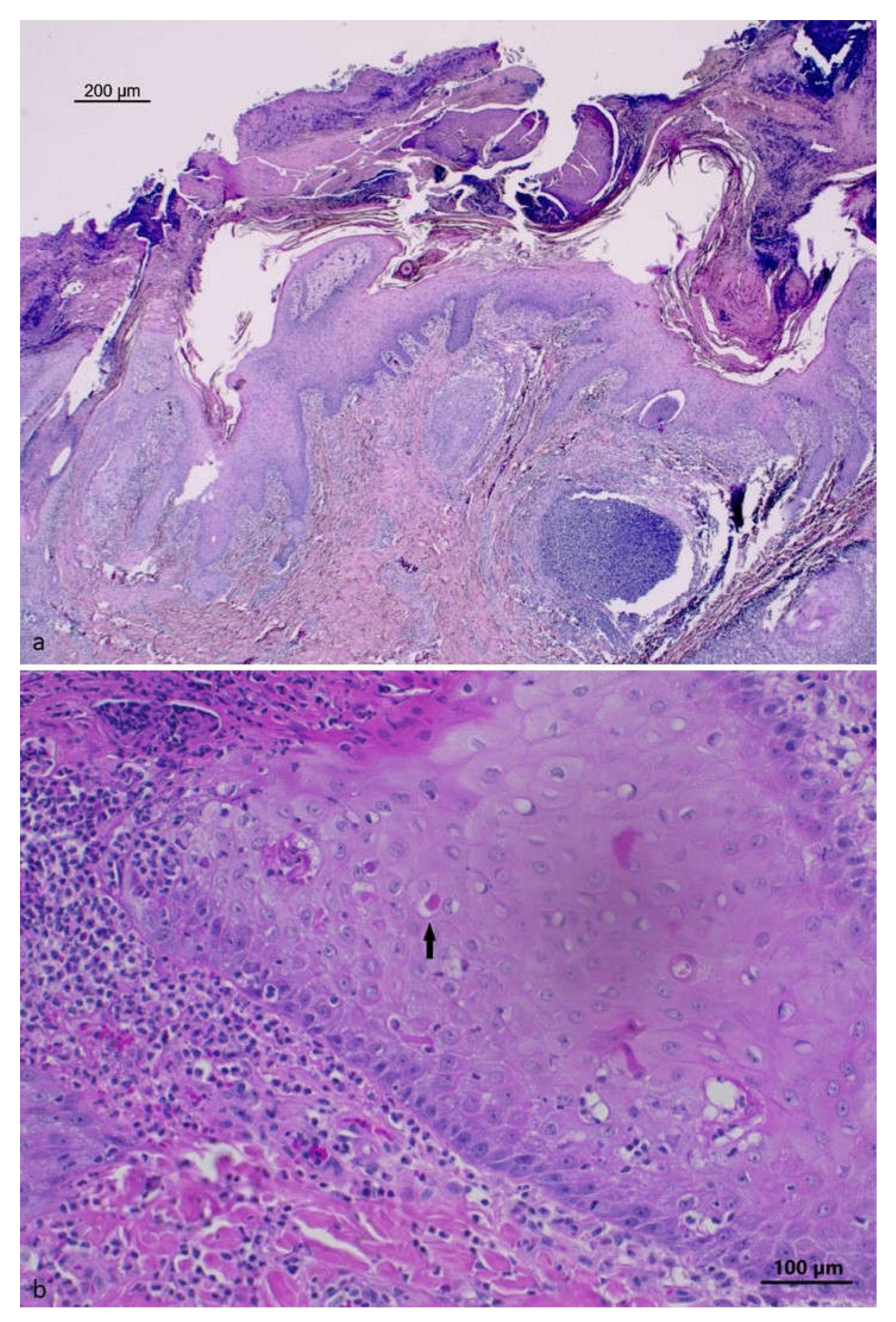
3.2. Histopathology
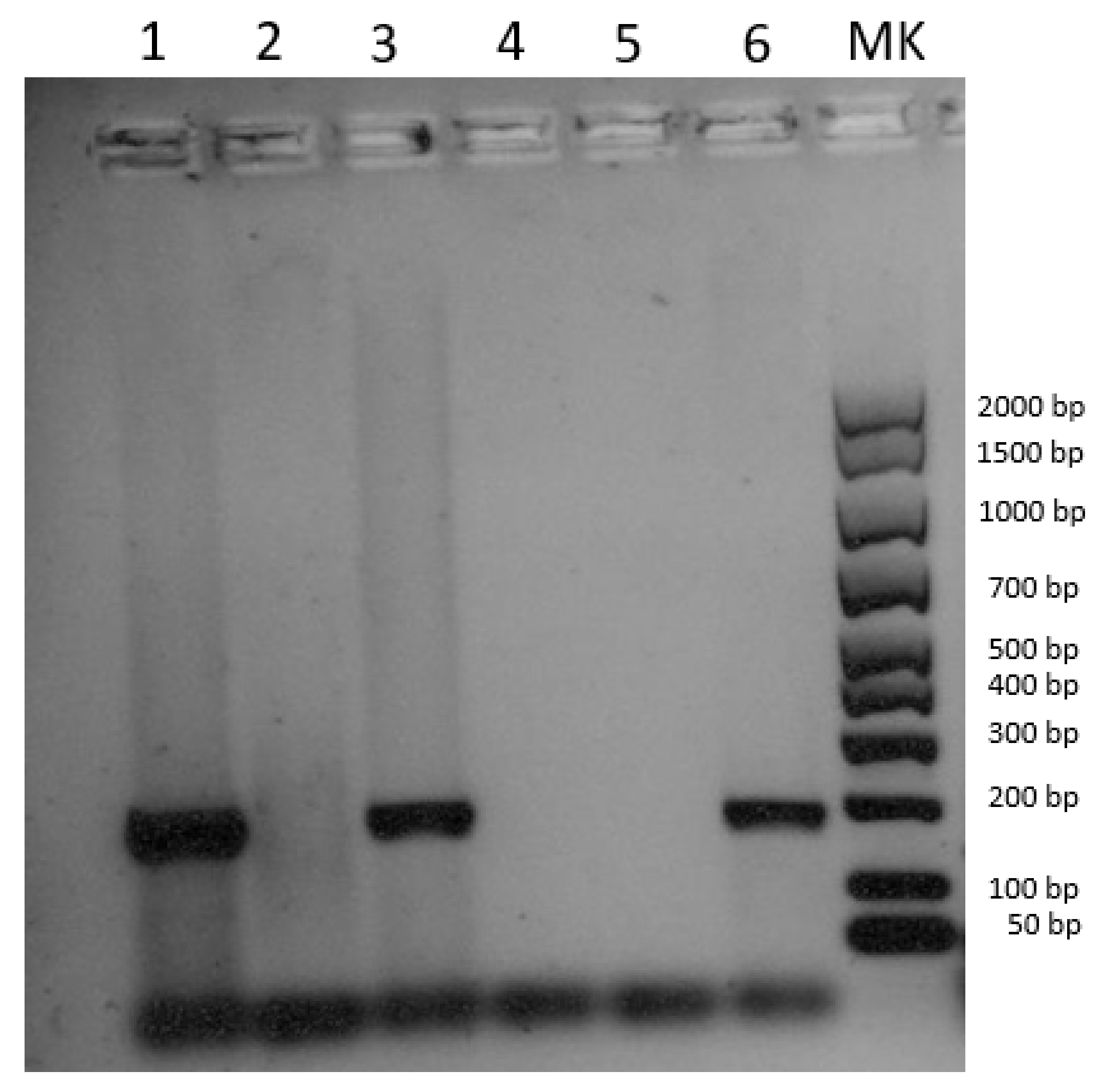
3.3. Molecular Analysis
3.3.1. SWPV
3.3.2. Other Viral Infections
4. Discussion
4.1. Gross and Histopathologic Features of Swinepox Virus Infection
4.2. Potential Vector-Borne Transmission
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kaiser, F.K.; Wiedemann, A.; Kühl, B.; Menke, L.; Beineke, A.; Baumgärtner, W.; Wohlsein, P.; Rigbers, K.; Becher, P.; Peters, M.; et al. Swinepox Virus Strains Isolated from Domestic Pigs and Wild Boar in Germany Display Altered Coding Capacity in the Terminal Genome Region Encoding for Species-Specific Genes. Viruses 2021, 13, 2038. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishnan, M.A.; Ashokkumar, D. Swinepox virus. In Recent Advances in Animal Virology; Springer Nature: Singapore, 2019; pp. 161–169. [Google Scholar] [CrossRef]
- Shope, R.E. Swine Pox. Arch. Gesamte Virusforsch. 1940, 1, 457–467. [Google Scholar] [CrossRef]
- Thibault, S.; Drolet, R.; Alain, R.; Dea, S. Congenital swine pox: A sporadic skin disorder in nursing piglets. Swine Health Prod. 1998, 6, 276–278. [Google Scholar]
- Spinola, W.T.J. Die Krankheiten der Schweine (1842); Hieschwald, A., Ed.; Kessinger Publishing: Berlin, Germany, 2010; p. 204.2. [Google Scholar]
- McNutt, S.; Murray, C.; Purwin, P. Swine Pox. J. Am. Vet. Med. Assoc. 1929, 74, 752–761. [Google Scholar]
- Bora, M.; Borah, B.; Kakati, P.; Nehar, S.; Dutta, L.J.; Mech, P.; Barman, N.N.; Venkatesan, G.; Reddy, G.M.; Das, S.K. Detection and characterization of Swinepox virus from pig population of Assam, a North-Eastern state of India. Indian J. Anim. Res. 2017, B-3352, 1–6. [Google Scholar] [CrossRef]
- Copland, J.W. Swine pox in Papua New Guinea. Trop. Anim. Health Prod. 1974, 6, 153–157. [Google Scholar] [CrossRef]
- Jubb, T.; Ellis, T.; Peet, R.; Parkinson, J. Swinepox in pigs in northern Western Australia. Aust. Vet. J. 1992, 69, 99. [Google Scholar] [CrossRef]
- Mech, P.; Bora, D.P.; Neher, S.; Barman, N.N.; Borah, P.; Tamuly, S.; Dutta, L.J.; Das, S.K. Identification of swinepox virus from natural outbreaks in pig population of Assam. Virus Dis. 2018, 29, 395–399. [Google Scholar] [CrossRef]
- Medaglia, M.L.G.; Pereira, A.D.C.; Freitas, T.R.; Damaso, C.R. Swinepox Virus Outbreak, Brazil, 2011. Emerg. Infect. Dis. 2011, 17, 1976–1978. [Google Scholar] [CrossRef]
- Olinda, R.G.; Maia, L.A.; Cargnelutti, J.F.; Gois, R.C.; Batista, J.S.; Flores, E.F.; Riet-Correa, F.; Dantas, A.F. Swinepox dermatitis in backyard pigs in Northeastern Brazil. Pesqui. Vet. Bras. 2016, 36, 468–472. [Google Scholar] [CrossRef]
- Olufemi, B.E.; Ayoade, G.O.; Ikede, B.O.; Akpavie, S.O.; Nwufoh, K.J. Swine pox in Nigeria. Vet. Rec. 1981, 109, 278–280. [Google Scholar] [CrossRef]
- Rheinbaben, F.v.; Gebel, J.; Exner, M.; Schmidt, A. Environmental resistance, disinfection, and sterilization of poxviruses. In Poxviruses; Birkhäuser: Basel, Switzerland, 2007; pp. 397–405. [Google Scholar] [CrossRef]
- Delhon, G.; Tulman, E.R.; Afonso, C.L.; Rock, D.L. Diseases of Swine, 10th ed.; Wiley-Blackwell: Chichester/Ames, UK, 2012. [Google Scholar]
- Steineck, T.; Kolodziejek, J.; Nowotny, N.; Schilcher, F. A case of Swine Pox in a Wild Boar (Sus Scrofa) in Austria. In Proceedings of the 6th Conference of the European Wildlife Disease Association, Uppsala, Sweden, 8–12 September 2004. [Google Scholar]
- Tittarelli, C.; Sozzi, E.; Spaggiari, B.; Alborali, G.L.; Cordioli, P.; Lavazza, A. Diagnosi al microscopio elettronico di swinepoxvirus, agente di malattia cutanea sporadica, durante il periodo 2002–2008 nel Nord Italia. In Proceedings of the Atti della SIPAS, Società Italiana di Patologia ed Allevamento dei suini XXXV Meeting Annuale, Modena, Singapore, 12–13 March 2009; pp. 507–512. [Google Scholar]
- Mariano, V.; Nardi, A.; Vergari, E.; Carletti, F.; Barbieri, L.; Cardeti, G. Poxvirus in a swine farm in Italy: A sporadic outbreak. Large Anim. Rev. 2015, 21, 219–220. [Google Scholar]
- Li, Y.; Meyer, H.; Zhao, H.; Damon, I.K. GC Content-Based Pan-Pox Universal PCR Assays for Poxvirus Detection. J. Clin. Microbiol. 2010, 48, 268–276. [Google Scholar] [CrossRef]
- Hoffmann, B.; Beer, M.; Schelp, C.; Schirrmeier, H.; Depner, K. Validation of a real-time RT-PCR assay for sensitive and specific detection of classical swine fever. J. Virol. Methods 2005, 130, 36–44. [Google Scholar] [CrossRef]
- Ogawa, H.; Taira, O.; Hirai, T.; Takeuchi, H.; Nagao, A.; Ishikawa, Y.; Tuchiya, K.; Nunoya, T.; Ueda, S. Multiplex PCR and multiplex RT-PCR for inclusive detection of major swine DNA and RNA viruses in pigs with multiple infections. J. Virol. Methods 2009, 160, 210–214. [Google Scholar] [CrossRef]
- Larochelle, R.; Antaya, M.; Morin, M.; Magar, R. Typing of porcine circovirus in clinical specimens by multiplex PCR. J. Virol. Methods 1999, 80, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Lager, K.M.; Richt, J.A.; Stoffregen, W.C.; Zhou, F.; Yoon, K.-J. Development of Real-Time Polymerase Chain Reaction Assays for Rapid Detection and Differentiation of Wild-Type Pseudorabies and Gene-Deleted Vaccine Viruses. J. Vet. Diagn. Investig. 2008, 20, 440–447. [Google Scholar] [CrossRef]
- Jothikumar, N.; Cromeans, T.L.; Robertson, B.H.; Meng, X.J.; Hill, V.R. A broadly reactive one-step real-time RT-PCR assay for rapid and sensitive detection of hepatitis E virus. J. Virol. Methods 2006, 131, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Gupta, N.; Fayaz, A.; Mageswary, R.; Bano, R.; ChandraSekar, S.; Muthuchelvan, D.; Dhama, K.; Pandey, A.B.; Andavar, R.M. Molecular epidemiology of swinepox viruses circulating in India. Vet. Q. 2023, 43, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Munz, E.; Dumbell, K. Swinepox. In Infectious Diseases of Livestock; Coetzer, J.A.W., Thomson, G.R., Tustin, R.C., Eds.; Oxford University Press: New York, NY, USA, 1994; pp. 627–629. [Google Scholar]
- Bratke, K.A.; McLysaght, A.; Rothenburg, S. A survey of host range genes in poxvirus genomes. Infect. Genet. Evol. 2013, 14, 406–425. [Google Scholar] [CrossRef]
- Paton, D.J.; Brown, I.H.; Fitton, J.; Wrathall, A.E. Congenital pig pox: A case report. Vet. Rec. 1990, 127, 204. [Google Scholar] [PubMed]
- Freitas, T.R.P. Swinepox Virus. In Diseases of Swine; John Wiley & Sons: Hoboken, NJ, USA, 2019; pp. 709–714. [Google Scholar]
- Riyesh, T.; Barua, S.; Kumar, N.; Jindal, N.; Bera, B.C.; Narang, G.; Mahajan, N.K.; Arora, D.; Anand, T.; Vaid, R.K.; et al. Isolation and genetic characterization of swinepox virus from pigs in India. Comp. Immunol. Microbiol. Infect. Dis. 2016, 46, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.B.; Olson, L.D. Experimental induction of cutaneous streptococcal abscesses in swine as a sequela to swinepox. Am. J. Vet. Res. 1980, 41, 341–347. [Google Scholar] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Etiological Agent | Method | Tested Sample |
|---|---|---|
| Swinepox virus | • End-point PCR (2720 Thermal cycler, Life Technologies) • Primers: ACACCAAAAACTCATATAACTTCT; CCTATTTTACTCCTTAGTAAATGAT (Li et al. [19]) • Target gene: 220 bp covering the insulin metalloproteinase-like protein gene and the intracellular mature virion membrane protein gene • PCR mixture: 1X Platinum qPCR SuperMix-UDG (Invitrogen), 0.1 µM of each primer and 2 µL of the template, in a total volume of 20 µL • PCR cycling conditions: 95 °C/5 min, followed by 40 cycles at 95 °C/30 s and 68 °C/60 s • Amplification products loaded onto agarose gel (1.8%) and compared to the Amplisize Molecular Ruler (Biorad) | Skin lesions Submandibular lymph node Spleen |
| African Swine Fever virus | • Real-time PCR (CFX96 Biorad) • Primers and probe: NA • Target gene: virion capsid protein p72 encoded by the B646L gene • PCR mixture: ID Gene™ African Swine Fever Duplex kit (IDVet) following the manufacturer’s protocol • PCR cycling conditions: 95 °C/10 min, followed by 40 cycles at 95 °C/15 s and 60 °C/60 s | Spleen |
| Classical Swine Fever virus | • Real-time PCR (CFX96 Biorad) • Primers and probe: CSF100-F: ATGCCCAYAGTAGGACTAGCA; CSF192-R: CTACTGACGACTGTCCTGTAC; CSF-P: TGGCGAGCTCCCTGGGTGGTCTAAGT (Hoffman et al. [20]) • Target gene: 93 bp pf the 5′ non-translated region • PCR mixture: 4X Reliance One Step multiplex Supermix (Biorad), 0.2 µM of each primer and 5 µL of the template, in a total volume of 20 µL • PCR cycling conditions: 50 °C/10 min, 95 °C/10 min, followed by 40 cycles at 95 °C/10 s and 60 °C/30 s | Spleen |
| Porcine Parvovirus (PPV) | • End-point PCR (2720 Thermal cycler, Applied Biosystems) • Primers: PPV-F: CACAGAAGCAACAGCAATTAGG; PPV-R: CTAGCTCTTGTGAAGATGTGG (Ogawa et al. [21]) • Target gene: 203 bp of the VP2 gene • PCR mixture: 1X Buffer of Taq Platinum DNA polymerase (Invitrogen), 0.05 U/µL of Taq polymerase, 3 mM of MgCl2, 200 µM of dNTP, 1µM of each primer and 2 µL of the template, in a total volume of 25 µL • PCR cycling conditions: 94 °C/1 min, followed by 35 cycles at 94 °C/30 min, 53 °C/90 s and 72 °C/90 s; finally, 72 °C/10 min • Amplification products were loaded onto agarose gel (1.8%) and compared to the Amplisize Molecular Ruler (Biorad) | Mesenteric lymph node |
| Porcine circovirus type II (PCV type II) | • End-point PCR (2720 Thermal cycler, Applied Biosystems) • Primers: CF8: TAGGTTAGGGCTGTGGCCTT; CR8: CCGCACCTTCGGATATACTG (Larochelle et al. [22]) • Target gene: 263 bp fragment of the ORF2 region • PCR mixture: 1x Buffer of AmpliTaq Gold DNA polymerase (Thermofisher Scientific), 0.05 U/µL of Taq polymerase, 1.5 mM of MgCl2, 200 µM of dNTP, 1µM of each primer and 2 µL of the template, in a total volume of 50 µL • PCR cycling conditions: 95 °C/15 min, 35 cycles of denaturation at 95 °C/20 s, annealing at 55 °C/30 s and extension at 72 °C/30 s and a final extension of 72 °C/5 min | Mesenteric lymph node |
| Aujeszky’s disease virus | • Real-time PCR (CFX96 Biorad) • Primers and probe: gB718F: ACAAGTTCAAGGCCCACATCTAC; gB812R: GTCYGTGAAGCGGTTCGTGAT; gB785P: ACGTCATCGTCACGACC (Ma et al. [23]) • Target gene: glycoproteins B gene • PCR mixture: 1X Platinum qPCR SuperMix-UDG (Invitrogen), 0.4 µM of each primer, 0.2 µM of the probe and 2.5 µL of the template, in a total volume of 25 µL • PCR cycling conditions: 50 °C for 2 min, 95 °C for 15 min, followed by 45 cycles at 94 °C for 15 s and 62 °C for 60 s | Lungs |
| Hepatitis E virus (HEV) | • Real-time PCR (CFX96 Biorad) • Primers and probes: JVHEVF: GGTGGTTTCTGGGGTGAC; JVHEVR: AGGGGTTGGTTGGATGAA; JVHEVP: TGATTCTCAGCCCTTCGC (Jothikumar et al. [24]). • Target gene: ORF3 region • PCR mixture: 1X Reaction mix of SuperScriptIII Platinum One-Step qRT-PCR System (Invitrogen), 0.5 µL of Taq mix, 1 mM of MgSO4, 0.25 µM of each primer, 0.1µM of the probe and 3 µL of the template, in a total volume of 25 µL • PCR cycling conditions: 50 °C for 30 min and 95 °C for 15 min followed by 45 cycles at 95 °C for 10 s, 55 °C for 20 s and 72 °C for 15 s | Liver |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guardone, L.; Varello, K.; Listorti, V.; Peletto, S.; Wolfsgruber, L.; Zoccola, R.; Montemurro, V.; Messana, E.; Bozzetta, E.; Acutis, P.; et al. First Report of Swinepox in a Wild Boar in Italy: Pathologic and Molecular Findings. Pathogens 2023, 12, 472. https://doi.org/10.3390/pathogens12030472
Guardone L, Varello K, Listorti V, Peletto S, Wolfsgruber L, Zoccola R, Montemurro V, Messana E, Bozzetta E, Acutis P, et al. First Report of Swinepox in a Wild Boar in Italy: Pathologic and Molecular Findings. Pathogens. 2023; 12(3):472. https://doi.org/10.3390/pathogens12030472
Chicago/Turabian StyleGuardone, Lisa, Katia Varello, Valeria Listorti, Simone Peletto, Lara Wolfsgruber, Roberto Zoccola, Vittoria Montemurro, Erika Messana, Elena Bozzetta, Pierluigi Acutis, and et al. 2023. "First Report of Swinepox in a Wild Boar in Italy: Pathologic and Molecular Findings" Pathogens 12, no. 3: 472. https://doi.org/10.3390/pathogens12030472
APA StyleGuardone, L., Varello, K., Listorti, V., Peletto, S., Wolfsgruber, L., Zoccola, R., Montemurro, V., Messana, E., Bozzetta, E., Acutis, P., Masoero, L., & Razzuoli, E. (2023). First Report of Swinepox in a Wild Boar in Italy: Pathologic and Molecular Findings. Pathogens, 12(3), 472. https://doi.org/10.3390/pathogens12030472