
Screening for Influenza and Morbillivirus in Seals and Porpoises in the Baltic and North Sea

 , , , , , , , , , and
, , , , , , , , , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Virus Screening
3. Results & Discussion
3.1. Viral Pathogens Only Detected in Samples Collected during Known UMEs
3.2. Morbillivirus in North European Marine Mammals
3.3. Influenza (IAV) in North European Marine Mammals
3.4. Implications and Considerations for Future Screening Efforts
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harwood, J.; Hall, A. Mass Mortality in Marine Mammals: Its Implications for Population Dynamics and Genetics. Trends Ecol. Evol. 1990, 5, 254–257. [Google Scholar] [CrossRef]
- Ross, P.; de Swart, R.; Addison, R.; van Loveren, H.; Vos, J.; Osterhaus, A. Contaminant-Induced Immunotoxicity in Harbour Seals: Wildlife at Risk? Toxicology 1996, 112, 157–169. [Google Scholar] [CrossRef]
- Albouy, C.; Delattre, V.; Donati, G.; Frölicher, T.L.; Albouy-Boyer, S.; Rufino, M.; Pellissier, L.; Mouillot, D.; Leprieur, F. Global Vulnerability of Marine Mammals to Global Warming. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Sanderson, C.E.; Alexander, K.A. Unchartered Waters: Climate Change Likely to Intensify Infectious Disease Outbreaks Causing Mass Mortality Events in Marine Mammals. Glob. Chang. Biol. 2020, 26, 4284–4301. [Google Scholar] [CrossRef]
- Wilcox, C.; Mallos, N.J.; Leonard, G.H.; Rodriguez, A.; Hardesty, B.D. Using Expert Elicitation to Estimate the Impacts of Plastic Pollution on Marine Wildlife. Mar. Policy 2016, 65, 107–114. [Google Scholar] [CrossRef]
- Zantis, L.J.; Carroll, E.L.; Nelms, S.E.; Bosker, T. Marine Mammals and Microplastics: A Systematic Review and Call for Standardisation. Environ. Pollut. 2021, 269, 116142. [Google Scholar] [CrossRef] [PubMed]
- Helle, E.; Olsson, M.; Jensen, S. PCB Levels Correlated with Pathological Changes in Seal Uteri. Ambio 1976, 5, 261–262. [Google Scholar] [CrossRef]
- Helle, E.; Olsson, M.; Jensen, S. DDT and PCB Levels and: Reproduction in Ringed Seal from the Bothnian Bay. Ambio 1976, 5, 188–189. [Google Scholar]
- Bergman, A.; Olsson, M. Pathology of Baltic Grey Seal and Ringed Seal Females with Special Reference to Adrenocortical Hyperplasia: Is Environmental Pollution the Cause of a Widely Distributed Disease Syndrome? Finn. Game Res. 1985, 44, 47–62. [Google Scholar]
- Bergman, A. Pathological Changes in Seals in Swedish Waters: The Relation to Environmental Pollution; Swedish University of Agricultural Sciences: Uppsala, Sweden, 2007. [Google Scholar]
- Harding, K.C.; Härkönen, T.J. Development in the Baltic Grey Seal (Halichoerus grypus) and Ringed Seal (Phoca hispida) Populations during the 20th Century. Ambio 1999, 28, 619–627. [Google Scholar] [CrossRef]
- Olsen, M.T.; Andersen, S.M.; Teilman, J.; Dietz, R.; Edrén, S.M.C.; Linnet, A.; Härkönen, T. Status of the Harbour Seal (Phoca vitulina) in Southern Scandinavia. NAMMCO Sci. Publ. 2010, 8, 77–94. [Google Scholar] [CrossRef]
- Härkönen, T.; Stenman, O.; Jüssi, M.; Jüssi, I.; Sagitov, R.; Verevkin, M. Population Size and Distribution of the Baltic Ringed Seal (Phoca hispida botnica). NAMMCO Sci. Publ. 1998, 1, 167. [Google Scholar] [CrossRef]
- Lacey, C.; Gilles, A.; Börjesson, P.; Herr, H.; Macleod, K.; Ridoux, V.; Santos, M.B.; Scheidat, M.; Teilmann, J.; Sveegaard, S.; et al. Modelled Density Surfaces of Cetaceans in European Atlantic Waters in Summer 2016 from the SCANS-III Aerial and Shipboard Surveys. Final Report. 2022. Available online: https://scans3.wp.st-andrews.ac.uk/files/2022/08/SCANS-III_density_surface_modelling_report_final_20220815.pdf (accessed on 20 February 2023).
- ASCOBANS. Recovery Plan for Baltic Harbour Porpoises (Jastarnia Plan). 2002. Available online: www.bfn.de/fileadmin/MDB/documents/themen/artenschutz/pdf/Jastarnia_Plan.pdf (accessed on 16 August 2021).
- ASCOBANS. Recovery Plan for Baltic Harbour Porpoises Jastarnia Plan (2009 Revision). 2009. Available online: https://www.bfn.de/fileadmin/BfN/artenschutz/Dokumente/Jastarnia_Plan_2009_Revision.pdf (accessed on 16 August 2021).
- Benke, H.; Bräger, S.; Dähne, M.; Gallus, A.; Hansen, S.; Honnef, C.G.; Jabbusch, M.; Koblitz, J.C.; Krügel, K.; Liebschner, A.; et al. Baltic Sea Harbour Porpoise Populations: Status and Conservation Needs Derived from Recent Survey Results. Mar. Ecol. Prog. Ser. 2014, 495, 275–290. [Google Scholar] [CrossRef]
- Carlén, I.; Thomas, L.; Carlström, J.; Amundin, M.; Teilmann, J.; Tregenza, N.; Tougaard, J.; Koblitz, J.C.; Sveegaard, S.; Wennerberg, D.; et al. Basin-Scale Distribution of Harbour Porpoises in the Baltic Sea Provides Basis for Effective Conservation Actions. Biol. Conserv. 2018, 226, 42–53. [Google Scholar] [CrossRef]
- Sonne, C.; Lakemeyer, J.; Desforges, J.-P.; Eulaers, I.; Persson, S.; Stokholm, I.; Galatius, A.; Gross, S.; Gonnsen, K.; Lehnert, K.; et al. A Review of Pathogens in Selected Baltic Sea Indicator Species. Environ. Int. 2020, 137, 105565. [Google Scholar] [CrossRef] [PubMed]
- Bodewes, R.; Zohari, S.; Krog, J.S.; Hall, M.D.; Harder, T.C.; Bestebroer, T.M.; van de Bildt, M.W.G.; Spronken, M.I.; Larsen, L.E.; Siebert, U.; et al. Spatiotemporal Analysis of the Genetic Diversity of Seal Influenza A(H10N7) Virus, Northwestern Europe. J. Virol. 2016, 90, 4269–4277. [Google Scholar] [CrossRef]
- Stokholm, I.; Härkönen, T.; Harding, K.C.; Siebert, U.; Lehnert, K.; Dietz, R.; Teilmann, J.; Galatius, A.; Havmøller, L.W.; Carroll, E.L.; et al. Phylogenomic Insights to the Origin and Spread of Phocine Distemper Virus in European Harbour Seals in 1988 and 2002. Dis. Aquat. Organ. 2019, 133, 47–56. [Google Scholar] [CrossRef]
- Härkönen, T.; Bäcklin, B.M.; Barrett, T.; Bergman, A.; Corteyn, M.; Dietz, R.; Harding, K.C.; Malmsten, J.; Roos, A.; Teilmann, J. Mass Mortality in Harbour Seals and Harbour Porpoises Caused by an Unknown Pathogen. Vet. Rec. 2008, 162, 555–556. [Google Scholar] [CrossRef]
- Härkönen, T.; Dietz, R.; Reijnders, P.; Teilmann, J.; Harding, K.; Hall, A.; Brasseur, S.; Siebert, U.; Goodman, S.J.; Jepson, P.D.; et al. A Review of the 1988 and 2002 Phocine Distemper Virus Epidemics in European Harbour Seals. Dis. Aquat. Organ. 2006, 68, 115–130. [Google Scholar] [CrossRef]
- Zohari, S.; Neimanis, A.; Härkönen, T.; Moraeus, C.; Valarcher, J.F. Avian Influenza A (H10N7) Virus Involvement in Mass Mortality of Harbour Seals (Phoca vitulina) in Sweden, March through October 2014. Eurosurveillance 2014, 19, 20967. [Google Scholar] [CrossRef]
- Krog, J.S.; Hansen, M.S.; Holm, E.; Hjulsager, C.K.; Chriél, M.; Pedersen, K.; Andresen, L.O.; Abildstrøm, M.; Jensen, T.H.; Larsen, L.E. Influenza A (H10N7) Virus in Dead Harbor Seals, Denmark. Emerg. Infect. Dis. 2015, 21, 684–687. [Google Scholar] [CrossRef] [PubMed]
- van Bressem, M.-F.; Duignan, P.; Banyard, A.; Barbieri, M.; Colegrove, K.; de Guise, S.; di Guardo, G.; Dobson, A.; Domingo, M.; Fauquier, D.; et al. Cetacean Morbillivirus: Current Knowledge and Future Directions. Viruses 2014, 6, 5145–5181. [Google Scholar] [CrossRef] [PubMed]
- McCullough, S.J.; McNeilly, F.; Allan, G.M.; Kennedy, S.; Smyth, J.A.; Cosby, S.L.; McQuaid, S.; Rima, B.K. Isolation and Characterisation of a Porpoise Morbillivirus. Arch. Virol. 1991, 118, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, S.; Smyth, J.A.; Cush, P.F.; McCullough, S.J.; Allan, G.M.; McQuaid, S. Viral Distemper Now Found in Porpoises. Nature 1988, 336, 21. [Google Scholar] [CrossRef]
- Domingo, M.; Ferrer, L.; Pumarola, M.; Marco, A. Morbillivirus in Dophins. Nature 1990, 348, 21. [Google Scholar] [CrossRef]
- Visser, I.K.G.; van Bressem, M.F.; de Swart, R.L.; van de Bildt, M.W.G.; Vos, H.W.; van der Heijden, R.W.J.; Saliki, J.T.; Orvell, C.; Kitching, P.; Kuiken, T.; et al. Characterization of Morbilliviruses Isolated from Dolphins and Porpoises in Europe. J. Gen. Virol. 1993, 74, 631–641. [Google Scholar] [CrossRef]
- Birkun, A.; Kuiken, T.; Krivokhizhin, S.; Haines, D.M.; Osterhaus, A.D.; van de Bildt, M.W.; Joiris, C.R.; Siebert, U. Epizootic of Morbilliviral Disease in Common Dolphins (Deiphinus deiphis ponticus) from the Black Sea. Vet. Rec. 1999, 144, 85–92. [Google Scholar] [CrossRef]
- Jacob, J.M.; West, K.L.; Levine, G.; Sanchez, S.; Jensen, B.A. Initial Characterization of Novel Beaked Whale Morbillivirus in Hawaiian Cetaceans. Dis. Aquat. Organ. 2016, 117, 215–227. [Google Scholar] [CrossRef]
- Jo, W.K.; Grilo, M.L.; Wohlsein, P.; Andersen-Ranberg, E.U.; Hansen, M.S.; Kinze, C.C.; Hjulsager, C.K.; Olsen, M.T.; Lehnert, K.; Prenger-Berninghoff, E.; et al. Dolphin Morbillivirus in a Fin Whale (Balaenoptera physalus) in Denmark, 2016. J. Wildl. Dis. 2017, 53, 921–924. [Google Scholar] [CrossRef]
- Lvov, D.K.; Zdanov, V.M.; Sazonov, A.A.; Braude, N.A.; Vladimirtceva, E.A.; Agafonova, L.V.; Skljanskaja, E.I.; Kaverin, N.V.; Reznik, V.I.; Pysina, T.V.; et al. Comparison of Influenza Viruses Isolated from Man and from Whales. Bull. World Health Organ. 1978, 56, 923–930. [Google Scholar]
- Hinshaw, V.S.; Bean, W.J.; Geraci, J.; Fiorelli, P.; Early, G.; Webster, R.G. Characterization of Two Influenza A Viruses from a Pilot Whale. J. Virol. 1986, 58, 655–656. [Google Scholar] [CrossRef]
- Nielsen, O.; Clavijo, A.; Boughen, J.A. Serologic Evidence of Influenza a Infection in Marine Mammals of Arctic Canada. J. Wildl. Dis. 2001, 37, 820–825. [Google Scholar] [CrossRef]
- Ohishi, K.; Maruyama, T.; Ninomiya, A.; Kida, H.; Zenitani, R.; Bando, T.; Fujise, Y.; Nakamatsu, K.; Miyazaki, N.; Boltunov, A.N. Serologic Investigation of Influenza A Virus Infection in Cetaceans from the Western North Pacific and the Southern Oceans. Mar. Mamm. Sci. 2006, 22, 214–221. [Google Scholar] [CrossRef]
- Bodewes, R.; Van De Bildt, M.W.G.; Van Elk, C.E.; Bunskoek, P.E.; Van De Vijver, D.A.M.C.; Smits, S.L.; Osterhaus, A.D.M.E.; Kuiken, T. No Serological Evidence That Harbour Porpoises Are Additional Hosts of Influenza B Viruses. PLoS ONE 2014, 9, e89058. [Google Scholar] [CrossRef]
- HELCOM. Report of the “HELCOM Marine Mammal Health Group” Activities in 2019. In Proceedings of the 13th Meeting of HELCOM Expert Group on Marine Mammals, Helsinki, Finland, 24–26 September 2019; pp. 1–7. [Google Scholar]
- Siebert, U.; Wohlsein, P.; Lehnert, K.; Baumgärtner, W. Pathological Findings in Harbour Seals (Phoca vitulina): 1996–2005. J. Comp. Pathol. 2007, 137, 47–58. [Google Scholar] [CrossRef]
- Siebert, U.; Wünschmann, A.; Weiss, R.; Frank, H.; Benke, H.; Frese, K. Post-Mortem Findings in Harbour Porpoises (Phocoena phocoena) from the German North and Baltic Seas. J. Comp. Pathol. 2001, 124, 102–114. [Google Scholar] [CrossRef]
- Sierra, E.; Sánchez, S.; Saliki, J.T.; Blas-Machado, U.; Arbelo, M.; Zucca, D.; Fernández, A. Retrospective Study of Etiologic Agents Associated with Nonsuppurative Meningoencephalitis in Stranded Cetaceans in the Canary Islands. J. Clin. Microbiol. 2014, 52, 2390–2397. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.L.; Siebert, U.; Lakemeyer, J.; Grilo, M.; Pawliczka, I.; Wu, N.H.; Valentin-Weigand, P.; Haas, L.; Herrler, G. Highly Pathogenic Avian Influenza A (H5N8) Virus in Gray Seals, Baltic Sea. Emerg. Infect. Dis. 2019, 25, 2295–2298. [Google Scholar] [CrossRef] [PubMed]
- Postel, A.; King, J.; Kaiser, F.K.; Kennedy, J.; Lombardo, M.S.; Reineking, W.; de le Roi, M.; Harder, T.; Pohlmann, A.; Gerlach, T.; et al. Infections with Highly Pathogenic Avian Influenza A Virus (HPAIV) H5N8 in Harbor Seals at the German North Sea Coast, 2021. Emerg. Microbes Infect. 2022, 11, 725–729. [Google Scholar] [CrossRef] [PubMed]
- Ludes-Wehrmeister, E.; Dupke, C.; Harder, T.C.; Baumgärtner, W.; Haas, L.; Teilmann, J.; Dietz, R.; Jensen, L.F.; Siebert, U. Phocine Distemper Virus (PDV) Seroprevalence as Predictor for Future Outbreaks in Harbour Seals. Vet. Microbiol. 2016, 183, 43–49. [Google Scholar] [CrossRef]
- Jauniaux, T.; Boseret, G.; Desmecht, M.; Haelters, J.; Manteca, C.; Tavernier, J.; van Gompel, J.; Coignoul, F. Morbillivirus in Common Seals Stranded on the Coasts of Belgium and Northern France during Summer 1998. Vet. Rec. 2001, 148, 587–591. [Google Scholar] [CrossRef] [PubMed]
- Henderson, G.; Trudgett, A.; Lyons, C.; Ronald, K. Demonstration of Antibodies in Archival Sera from Canadian Seals Reactive with a European Isolate of Phocine Distemper Virus. Sci. Total Environ. 1992, 115, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Dietz, R.; Ansen, C.T.; Have, P.; Heide-Jørgensen, M.P. Clue to Seal Epizootic? Nature 1989, 338, 627. [Google Scholar] [CrossRef] [PubMed]
- Daoust, P.-Y.; Haines, D.M.; Thorsen, J.; Duignan, P.J.; Geraci, J.R. Phocine Distemper in Harp Seals (Phoca groenlandica) from the Gulf of St. Lawrence, Canada. J. Wildl. Dis. 1993, 29, 114–117. [Google Scholar] [CrossRef]
- Duignan, P.J.; Saliki, J.T.; St Aubin, D.J.; Early, G.; Sadove, S.; House, J.A.; Kovacs, K.; Geraci, J.R. Epizootiology of Morbillivirus Infection in North American Harbor Seals (Phoca vitulina) and Gray Seals (Halichoerus grypus). J. Wildl. Dis. 1995, 31, 491–501. [Google Scholar] [CrossRef]
- Duignan, P.J.; Nielsen, O.; House, C.; Kovacs, K.M.; Duffy, N.; Early, G.; Sadove, S.; St. Aubin, D.J.; Rima, B.K.; Geraci, J.R. Epizootiology of Morbillivirus Infection in Harp, Hooded, and Ringed Seals from the Canadian Arctic and Western Atlantic. J. Wildl. Dis. 1997, 33, 7–19. [Google Scholar] [CrossRef]
- VanWormer, E.; Mazet, J.A.K.; Hall, A.; Gill, V.A.; Boveng, P.L.; London, J.M.; Gelatt, T.; Fadely, B.S.; Lander, M.E.; Sterling, J.; et al. Viral Emergence in Marine Mammals in the North Pacific May Be Linked to Arctic Sea Ice Reduction. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Puryear, W.; Sawatzki, K.; Bogomolni, A.; Hill, N.; Foss, A.; Stokholm, I.; Olsen, M.T.; Nielsen, O.; Waltzek, T.; Goldstein, T.; et al. Longitudinal Analysis of Pinnipeds in the Northwest Atlantic Provides Insights on Endemic Circulation and Regional Immunity to Phocine Distemper Virus. Proc. R. Soc. B 2021, 288, 1–10. [Google Scholar] [CrossRef]
- Van Elk, C.E.; Van De Bildt, M.W.G.; Van Run, P.R.W.A.; Bunskoek, P.; Meerbeek, J.; Foster, G.; Osterhaus, A.D.M.E.; Kuiken, T. Clinical, Pathological, and Laboratory Diagnoses of Diseases of Harbour Porpoises (Phocoena phocoena), Live Stranded on the Dutch and Adjacent Coasts from 2003 to 2016. Vet. Res. 2019, 50, 1–17. [Google Scholar] [CrossRef]
- Siebert, U.; Pawliczka, I.; Benke, H.; von Vietinghoff, V.; Wolf, P.; Pilāts, V.; Kesselring, T.; Lehnert, K.; Prenger-Berninghoff, E.; Galatius, A.; et al. Health Assessment of Harbour Porpoises (Phocoena phocoena) from Baltic Area of Denmark, Germany, Poland and Latvia. Environ. Int. 2020, 143, 105904. [Google Scholar] [CrossRef]
- Wohlseinf, P.; Puff, C.; Kreutzer, M.; Siebert, U.; Baumgärtner, W. Distemper in a Dolphin. Emerg. Infect. Dis. 2007, 13, 1959–1961. [Google Scholar] [CrossRef] [PubMed]
- Müller, G.; Siebert, U.; Wünschmann, A.; Artelt, A.; Baumgärtner, W. Immunohistological and Serological Investigation of Morbillivirus Infection in Harbour Porpoises (Phocoena phocoena) from the German Baltic and North Sea. Vet. Microbiol. 2000, 75, 17–25. [Google Scholar] [CrossRef]
- Van Bressem, M.F.; Jepson, P.; Barrett, T. Further Insight on the Epidemiology of Cetacean Morbillivirus in the Northeastern Atlantic. Mar. Mamm. Sci. 1998, 14, 605–613. [Google Scholar] [CrossRef]
- Wunschmann, A.; Siebert, U.; Frese, K.; Weiss, R.; Lockyer, C.; Heide-Jørgensen, M.P.; Müller, G.; Baumgärtner, W. Evidence of Infectious Diseases in Harbour Porpoises (Phocoena phocoena) Hunted in the Waters of Greenland and by-Caught in the German North Sea and Baltic Sea. Vet. Rec. 2001, 148, 715–720. [Google Scholar] [CrossRef]
- Duignan, P.J.; House, C.; Odell, D.K.; Wells, R.S.; Hansen, L.J.; Walsh, M.T.; St. Aubin, D.J.; Rima, B.K.; Geraci, J.R. Morbillivirus Infection in Bottlenose Dolphins: Evidence for Recurrent Epizootics in the Western Atlantic and Gulf of Mexico. Mar. Mamm. Sci. 1996, 12, 499–515. [Google Scholar] [CrossRef]
- Lipscomb, P.; Yvonne, F.; Moffett, D.; Kennedy, S. Morbilliviral Disease in Atlantic Bottlenose Dolphins (Tursiops truncatus) from the 1987-1988 Epizootic. J. Wildl. Dis. 1994, 30, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Jo, W.K.; Kruppa, J.; Habierski, A.; van de Bildt, M.; Mazzariol, S.; di Guardo, G.; Siebert, U.; Kuiken, T.; Jung, K.; Osterhaus, A.; et al. Evolutionary Evidence for Multi-Host Transmission of Cetacean Morbillivirus. Emerg. Microbes Infect. 2018, 7, 201. [Google Scholar] [CrossRef] [PubMed]
- Adlhoch, C.; Fusaro, A.; Gonzales, J.L.; Kuiken, T.; Marangon, S.; Niqueux, É.; Staubach, C.; Terregino, C.; Aznar, I.; Guajardo, I.M.; et al. Avian Influenza Overview March–June 2022. EFSA J. 2022, 20, 1–67. [Google Scholar] [CrossRef]
- Venkatesh, D.; Bianco, C.; Núnez, A.; Collins, R.; Thorpe, D.; Reid, S.M.; Brookes, S.M.; Essen, S.; McGinn, N.; Seekings, J.; et al. Detection of H3N8 Influenza A Virus with Multiple Mammalian-Adaptive Mutations in a Rescued Grey Seal (Halichoerus grypus) Pup. Virus Evol. 2020, 6, 1–12. [Google Scholar] [CrossRef]
- Floyd, T.; Banyard, A.C.; Lean, F.Z.X.; Byrne, A.M.P.; Fullick, E.; Whittard, E.; Mollett, B.C.; Bexton, S.; Swinson, V.; Macrelli, M.; et al. Encephalitis and Death in Wild Mammals at a Rehabilitation Center after Infection with Highly Pathogenic Avian Influenza A (H5N8) Virus, United Kingdom. Emerg. Infect. Dis. 2021, 27, 2856–2863. [Google Scholar] [CrossRef]
- Statens Serum Institut. Fugleinfluenza i Dansk Sæl. Available online: https://www.ssi.dk/aktuelt/nyheder/2022/fugleinfluenza-i-dansk-sael (accessed on 19 September 2022). (In Danish).
- de Boer, G.F.; Back, W.; Osterhaus, A.D.M.E. An ELISA for Detection of Antibodies against Influenza A Nucleoprotein in Humans and Various Animal Species. Arch. Virol. 1990, 115, 47–61. [Google Scholar] [CrossRef] [PubMed]
- Bodewes, R.; Garcia, A.R.; Brasseur, S.M.; Conteras, G.J.S.; van de Bildt, M.W.G.; Koopmans, M.P.G.; Osterhaus, A.D.M.E.; Kuiken, T. Seroprevalence of Antibodies against Seal Influenza A (H10N7) Virus in Harbor Seals and Gray Seals from the Netherlands. PLoS ONE 2015, 10, e0144899. [Google Scholar] [CrossRef]
- Puryear, W.B.; Keogh, M.; Hill, N.; Moxley, J.; Josephson, E.; Davis, K.R.; Bandoro, C.; Lidgard, D.; Bogomolni, A.; Levin, M.; et al. Prevalence of Influenza A Virus in Live-Captured North Atlantic Gray Seals: A Possible Wild Reservoir. Emerg. Microbes Infect. 2016, 5, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Silva, W.T.A.F.; Bottagisio, E.; Härkönen, T.; Galatius, A.; Olsen, M.T.; Harding, K.C. Risk for Overexploiting a Seemingly Stable Seal Population: Influence of Multiple Stressors and Hunting. Ecosphere 2021, 12, 1–17. [Google Scholar] [CrossRef]
- Bäcklin, B.M.; Moraeus, C.; Strömberg, A.; Karlsson, O.; Härkönen, T. Havet 2015/2016—Om Miljötillståndet I Svenska havsområden, Sälpopulationer och Sälhälsa. The Swedish Institute for the Marine Environment. May 2016, pp. 116–118. Available online: https://www.havochvatten.se/download/18.44ebc86154b1fe664adb068/1463996530913/havet_2015_16.pdf (accessed on 20 February 2023). (In Swedish).
- Galatius, A.; Teilmann, J.; Dähne, M.; Ahola, M.; Westphal, L.; Kyhn, L.A.; Pawliczka, I.; Olsen, M.T.; Dietz, R. Grey Seal Halichoerus grypus Recolonisation of the Southern Baltic Sea, Danish Straits and Kattegat. Wildl. Biol. 2020, 4, 1–10. [Google Scholar] [CrossRef]
- Ahola, M.; Authier, M.; Brasseur, S.; Carlström, J.; Chaudry, F.; Culloch, R.; Evans, P.; Galatius, A.; Geelhoed, S.; Gilles, A.; et al. Working Group on Marine Mammal Ecology (WGMME). ICES Sci. Rep. 2020, 2, 85. [Google Scholar]
- ICES. Working Group on Marine Mammal Ecology (WGMME). ICES Sci. Rep. 2022, 4, 1–151. [Google Scholar] [CrossRef]
- Ahola, M.; Halkka, A.; Jüssi, I.; Jüssi, M.; Kunnasranta, M.; Laine, A.; Nordström, M.; Tolvanen, P.; Verevkin, M.; Vysotsky, V. The Baltic Ringed Seal—An Arctic Seal in European Waters; WWF Suomi: Helsinki, Finland, 2017; pp. 1–32. Available online: https://wwf.fi/app/uploads/2/r/u/z4bm4bbejniod2hde4g2kce/wwf_norppa_2017_web_korj_d.pdf (accessed on 20 February 2023).
- ICES. Report of the Working Group on Marine Mammal Ecology (WGMME), 19–22 February 2018, La Rochelle, France; ICES CM 2018/ACOM:28; ICES: Copenhagen, Denmark, 2018. [Google Scholar]
- Amundin, M.; Carlström, J.; Thomas, L.; Carlén, I.; Teilmann, J.; Tougaard, J.; Loisa, O.; Kyhn, L.A.; Sveegaard, S.; Burt, M.L.; et al. Estimating the Abundance of the Critically Endangered Baltic Proper Harbour Porpoise (Phocoena phocoena) Population Using Passive Acoustic Monitoring. Ecol. Evol. 2022, 12, 56. [Google Scholar] [CrossRef] [PubMed]
- Owen, K.; Sköld, M.; Carlström, J. An Increase in Detection Rates of the Critically Endangered Baltic Proper Harbor Porpoise in Swedish Waters in Recent Years. Conserv. Sci. Pract. 2021, 3, 1–13. [Google Scholar] [CrossRef]
- Unger, B.; Nachtsheim, D.; Ramírez Martínez, N.; Siebert, U.; Sveegaard, S.; Kyhn, L.; Balle, J.D.; Teilmann, J.; Carlström, J.; Owen, K.; et al. MiniSCANS-II: Aerial Survey for Harbour Porpoises in the Western Baltic Sea, Belt Sea, the Sound and Kattegat in 2020. Joint Survey by Denmark, Germany and Sweden. Final Report to the Danish Environmental Protection Agency, the German Federal Agency for Nature Conservation and Swedish Agency for Marine and Water Management. 2021, pp. 1–30. Available online: https://dce.au.dk/fileadmin/dce.au.dk/Udgivelser/Eksterne_udgivelser/20210913_Report_MiniSCANSII_2020_revised.pdf (accessed on 20 February 2023).
- Black, F.L. Measles Endemicity in Insular Populations: Critical Community Size and Its Evolutionary Implication. J. Theor. Biol. 1966, 11, 207–211. [Google Scholar] [CrossRef]
- Galatius, A.; Abel, C.; Brackmann, J.; Brasseur, S.; Jess, A.; Meise, K.; Meyer, J.; Schop, J.; Siebert, U.; Teilmann, J.; et al. Harbour Seal Surveys in the Wadden Sea and Helgoland 2021; Common Wadden Sea Secretariat: Wilhelmshaven, Germany, 2021; pp. 1–6. Available online: https://www.waddensea-worldheritage.org/sites/default/files/2021_Harbour_Seal_Report.pdf (accessed on 20 February 2023).
- Brasseur, S.; Abel, C.; Galatius, A.; Jess, A.; Körber, P.; Meise, K.; Schop, J.; Siebert, U.; Teilmann, J.; Thøstesen, C.B. EG-Marine Mammals Grey Seal Surveys in the Wadden Sea and Helgoland in 2020–2021; Common Wadden Sea Secretariat: Wilhelmshaven, Germany, 2021; pp. 1–6. Available online: https://www.waddensea-worldheritage.org/sites/default/files/Wadden%20Sea_Grey_Seal_Report_2021_0.pdf (accessed on 20 February 2023).
- Hammond, P.; Lacey, C.; Gilles, A.; Viquerat, S.; Börjesson, P.; Herr, H.; Macleod, K.; Ridoux, V.; Santos, M.; Scheidat, M.; et al. Estimates of Cetacean Abundance in European Atlantic Waters in Summer 2016 from the SCANS-III Aerial and Shipboard Surveys; Wageningen Marine Research: Yerseke, The Netherlands, 2017. [Google Scholar]
- Fietz, K.; Galatius, A.; Teilmann, J.; Dietz, R.; Frie, A.K.; Klimova, A.; Palsbøll, P.J.; Jensen, L.F.; Graves, J.A.; Hoffman, J.I. Shift of Grey Seal Subspecies Boundaries in Response to Climate, Culling and Conservation. Mol. Ecol. 2016, 25, 4097–4112. [Google Scholar] [CrossRef] [PubMed]
- Svensson, C.J. Seal Dynamics on the Swedish West Coast: Scenarios of Competition as Baltic Grey Seal Intrude on Harbour Seal Territory. J. Sea Res. 2012, 71, 9–13. [Google Scholar] [CrossRef]
- van Neer, A.; Jensen, L.F.; Siebert, U. Grey Seal (Halichoerus grypus) Predation on Harbour Seals (Phoca vitulina) on the Island of Helgoland, Germany. J. Sea Res. 2015, 97, 1–4. [Google Scholar] [CrossRef]
- Wilson, L.J.; Hammond, P.S. The Diet of Harbour and Grey Seals around Britain: Examining the Role of Prey as a Potential Cause of Harbour Seal Declines. Aquat. Conserv. 2019, 29, 71–85. [Google Scholar] [CrossRef]
- Puryear, W.; Sawatzki, K.; Hill, N.; Foss, A.; Stone, J.J.; Walk, D.; Gilbert, K.; Murray, M.; Cox, E.; Patel, P. Outbreak of Highly Pathogenic Avian Influenza H5N1 in New England Seals. bioRxiv 2022, 1–10. [Google Scholar]
- Kydyrmanov, A.I.; Karamendin, K.O. Viruses of Marine Mammals and Metagenomic Monitoring of Infectious Diseases. Bulletin 2019, 4, 147–153. [Google Scholar] [CrossRef]


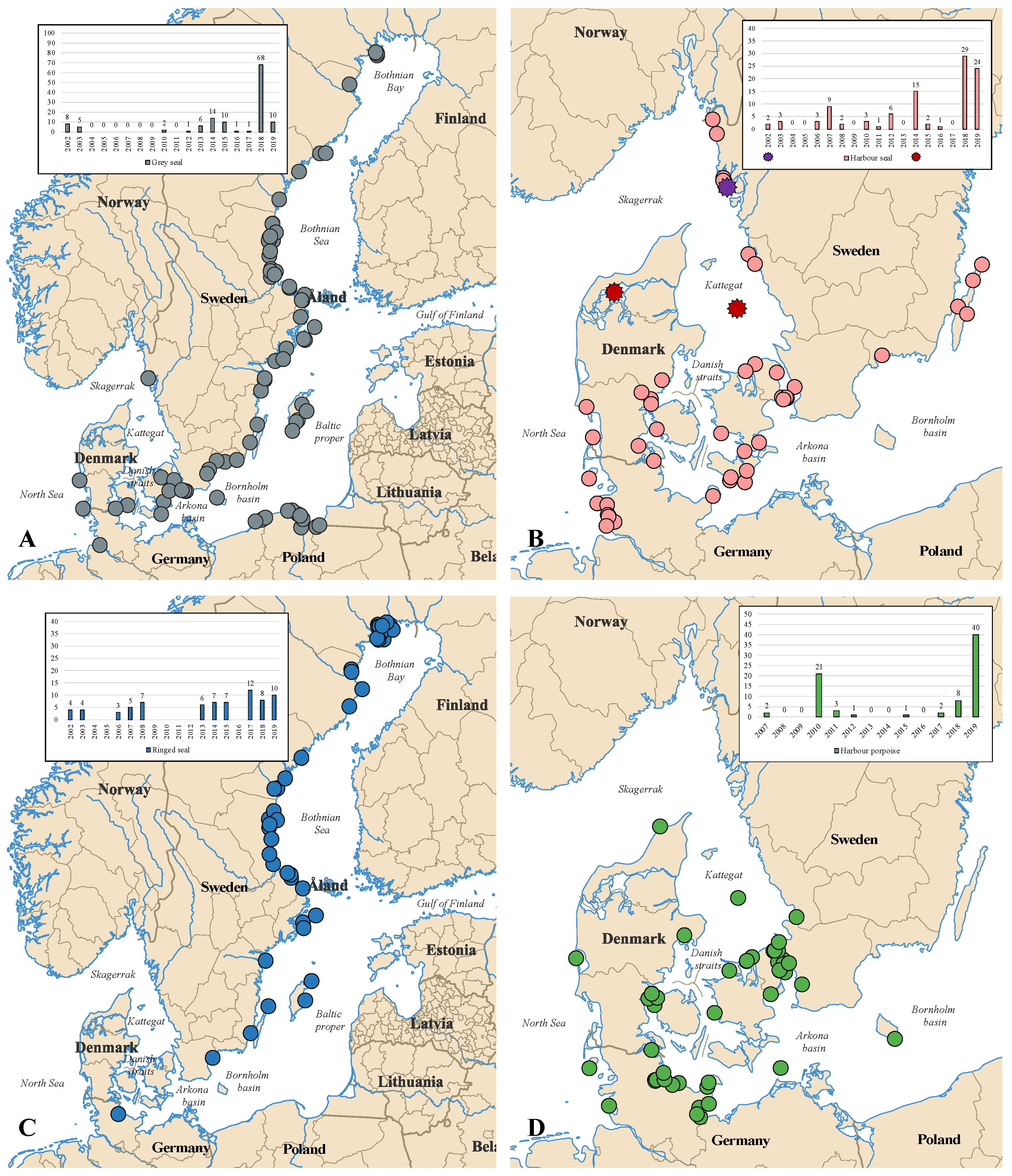{kind=link}
| Grey Seal | |||||
| Area | 2002–2003 | 2006–2008 | 2010–2012 | 2013–2015 | 2017–2019 |
| Skagerrak/Kattegat/the Sound/Danish straits/ | 1 | 3 | 3 | ||
| Arkona Basin | 1 | 2 | 5 | ||
| Bornholm Basin | 1 | 5 | 8 | ||
| Baltic proper | 3 | 5 | 27 | ||
| Bothnian Sea | 5 | 9 | 34 | ||
| Bothnian Bay | 2 | 5 | 2 | ||
| North Sea | 1 | 6 | |||
| Harbour seal | |||||
| Area | 2002–2003 | 2006–2008 | 2010–2012 | 2013–2015 | 2017–2019 |
| Skagerrak/Kattegat/the Sound/Danish straits/ | 3 | 14 | 5 | 9 | 32 |
| Arkona Basin | 4 | 1 | |||
| Bornholm Basin | 1 | ||||
| Baltic proper | 2 | 1 | 2 | ||
| Bothnian Sea | |||||
| Bothnian Bay | |||||
| North Sea | 2 | 19 | |||
| The Limfjord | 4 | ||||
| Ringed seal | |||||
| Area | 2002–2003 | 2006–2008 | 2010–2012 | 2013–2015 | 2017–2019 |
| Skagerrak/Kattegat/the Sound/Danish straits/ | 1 | ||||
| Arkona Basin | |||||
| Bornholm Basin | 1 | ||||
| Baltic proper | 2 | 1 | 3 | 1 | |
| Bothnian Sea | 5 | 8 | 7 | 3 | |
| Bothnian Bay | 1 | 5 | 10 | 25 | |
| Harbour porpoise | |||||
| Area | 2002–2003 | 2006–2008 | 2010–2012 | 2013–2015 | 2017–2019 |
| Skagerrak/Kattegat/the Sound/Danish straits/ | 2 | 24 | 1 | 46 | |
| Arkona Basin | |||||
| Bornholm Basin | 1 | ||||
| Baltic proper | |||||
| Bothnian Sea | |||||
| Bothnian Bay | |||||
| North Sea | 1 | 3 | |||
| Harbour Seal | |||||||||
| Year | Pups/calfs | Juveniles | Adults | NA | Total | IAV | PDV | IAV % | PDV % |
| 2002 | - | 1 | 1 | - | 2 | 0 | 1 | 0 | 50 |
| 2014 | 1 | 5 | 4 | 5 | 15 | 2 | 0 | 13 | 0 |
| 2002–2019 | 14 | 34 | 34 | 17 | 99 | 0 | 0 | 2 | 1 |
| Grey seal | |||||||||
| Year | Pups/calfs | Juveniles | Adults | NA | Total | IAV | PDV | IAV % | PDV % |
| 2002–2019 | 12 | 35 | 51 | 28 | 126 | 0 | 0 | 0 | 0 |
| Ringed seal | |||||||||
| Year | Pups/calfs | Juveniles | Adults | NA | Total | IAV | PDV | IAV % | PDV % |
| 2002–2019 | 21 | 8 | 20 | 24 | 73 | 0 | 0 | 0 | 0 |
| Harbour porpoise | |||||||||
| Year | Pups/calfs | Juveniles | Adults | NA | Total | IAV | PDV | IAV % | PDV % |
| 2007–2019 | 9 | 32 | 11 | 26 | 78 | 0 | 0 | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stokholm, I.; Baechlein, C.; Persson, S.; Roos, A.; Galatius, A.; Kyhn, L.A.; Sveegaard, S.; Thøstesen, C.B.; Olsen, M.T.; Becher, P.; et al. Screening for Influenza and Morbillivirus in Seals and Porpoises in the Baltic and North Sea. Pathogens 2023, 12, 357. https://doi.org/10.3390/pathogens12030357
Stokholm I, Baechlein C, Persson S, Roos A, Galatius A, Kyhn LA, Sveegaard S, Thøstesen CB, Olsen MT, Becher P, et al. Screening for Influenza and Morbillivirus in Seals and Porpoises in the Baltic and North Sea. Pathogens. 2023; 12(3):357. https://doi.org/10.3390/pathogens12030357
Chicago/Turabian StyleStokholm, Iben, Christine Baechlein, Sara Persson, Anna Roos, Anders Galatius, Line Anker Kyhn, Signe Sveegaard, Charlotte Bie Thøstesen, Morten Tange Olsen, Paul Becher, and et al. 2023. "Screening for Influenza and Morbillivirus in Seals and Porpoises in the Baltic and North Sea" Pathogens 12, no. 3: 357. https://doi.org/10.3390/pathogens12030357
APA StyleStokholm, I., Baechlein, C., Persson, S., Roos, A., Galatius, A., Kyhn, L. A., Sveegaard, S., Thøstesen, C. B., Olsen, M. T., Becher, P., & Siebert, U. (2023). Screening for Influenza and Morbillivirus in Seals and Porpoises in the Baltic and North Sea. Pathogens, 12(3), 357. https://doi.org/10.3390/pathogens12030357