
Differential Resistance of Borrelia burgdorferi Clones to Human Serum-Mediated Killing Does Not Correspond to Their Predicted Invasiveness


Abstract
:1. Introduction
2. Materials and Methods
2.1. B. burgdorferi ospC Clone Isolation and Characterization
2.2. Human Serum Collection and Validation
2.3. Serum Sensitivity Assay
2.4. Statistical Analysis
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kraiczy, P. Travelling between Two Worlds: Complement as a Gatekeeper for an Expanded Host Range of Lyme Disease Spirochetes. Vet. Sci. 2016, 3, 12. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-P.; Diuk-Wasser, M.A.; Stevenson, B.; Kraiczy, P. Complement Evasion Contributes to Lyme Borreliae–Host Associations. Trends Parasitol. 2020, 36, 634–645. [Google Scholar] [CrossRef] [PubMed]
- Tufts, D.M.; Hart, T.M.; Chen, G.F.; Kolokotronis, S.; Diuk-Wasser, M.A.; Lin, Y. Outer Surface Protein Polymorphisms Linked to Host-spirochete Association in Lyme Borreliae. Mol. Microbiol. 2019, 111, 868–882. [Google Scholar] [CrossRef] [PubMed]
- Kurtenbach, K.; De Michelis, S.; Etti, S.; Schäfer, S.M.; Sewell, H.-S.; Brade, V.; Kraiczy, P. Host Association of Borrelia burgdorferi Sensu Lato—The Key Role of Host Complement. Trends Microbiol. 2002, 10, 74–79. [Google Scholar] [CrossRef]
- Brisson, D.; Dykhuizen, D.E. ospC Diversity in Borrelia burgdorferi. Genetics 2004, 168, 713–722. [Google Scholar] [CrossRef]
- Hanincová, K.; Taragelová, V.; Koci, J.; Schäfer, S.M.; Hails, R.; Ullmann, A.J.; Piesman, J.; Labuda, M.; Kurtenbach, K. Association of Borrelia garinii and B. valaisiana with Songbirds in Slovakia. Appl. Environ. Microbiol. 2003, 69, 2825–2830. [Google Scholar] [CrossRef]
- Hanincová, K.; Schäfer, S.M.; Etti, S.; Sewell, H.-S.; Taragelová, V.; Ziak, D.; Labuda, M.; Kurtenbach, K. Association of Borrelia afzelii with Rodents in Europe. Parasitology 2003, 126, 11–20. [Google Scholar] [CrossRef]
- Kurtenbach, K.; Sewell, H.-S.; Ogden, N.H.; Randolph, S.E.; Nuttall, P.A. Serum Complement Sensitivity as a Key Factor in Lyme Disease Ecology. Infect. Immun. 1998, 66, 1248–1251. [Google Scholar] [CrossRef]
- Skare, J.T.; Garcia, B.L. Complement Evasion by Lyme Disease Spirochetes. Trends Microbiol. 2020, 28, 889–899. [Google Scholar] [CrossRef]
- Rus, H.; Cudrici, C.; Niculescu, F. The Role of the Complement System in Innate Immunity. IR 2005, 33, 103–112. [Google Scholar] [CrossRef]
- Sarma, J.V.; Ward, P.A. The Complement System. Cell Tissue Res. 2011, 343, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Combs, M.; Marcinkiewicz, A.L.; Dupuis, A.P.; Davis, A.D.; Lederman, P.; Nowak, T.A.; Stout, J.L.; Strle, K.; Fingerle, V.; Margos, G.; et al. Phylogenomic Diversity Elucidates Mechanistic Insights into Lyme Borreliae-Host Association. mSystems 2022, 7, e00488-22. [Google Scholar] [CrossRef] [PubMed]
- Steere, A.C.; Strle, F.; Wormser, G.P.; Hu, L.T.; Branda, J.A.; Hovius, J.W.R.; Li, X.; Mead, P.S. Lyme Borreliosis. Nat. Rev. Dis. Primers 2016, 2, 16090. [Google Scholar] [CrossRef]
- Wang, I.-N.; Dykhuizen, D.E.; Qiu, W.; Dunn, J.J.; Bosler, E.M.; Luft, B.J. Genetic Diversity of ospC in a Local Population of Borrelia burgdorferi Sensu Stricto. Genetics 1999, 151, 15–30. [Google Scholar] [CrossRef] [PubMed]
- Barbour, A.G.; Travinsky, B. Evolution and Distribution of the ospC Gene, a Transferable Serotype Determinant of Borrelia burgdorferi. mBio 2010, 1, e00153-10. [Google Scholar] [CrossRef]
- Hanincová, K.; Ogden, N.H.; Diuk-Wasser, M.; Pappas, C.J.; Iyer, R.; Fish, D.; Schwartz, I.; Kurtenbach, K. Fitness Variation of Borrelia burgdorferi Sensu Stricto Strains in Mice. Appl. Environ. Microbiol. 2008, 74, 153–157. [Google Scholar] [CrossRef]
- Wang, G.; Ojaimi, C.; Wu, H.; Saksenberg, V.; Iyer, R.; Liveris, D.; McClain, S.A.; Wormser, G.P.; Schwartz, I. Disease Severity in a Murine Model of Lyme Borreliosis Is Associated with the Genotype of the Infecting Borrelia burgdorferi Sensu Stricto Strain. J. Infect. Dis. 2002, 186, 782–791. [Google Scholar] [CrossRef]
- Baum, E.; Hue, F.; Barbour, A.G. Experimental Infections of the Reservoir Species Peromyscus Leucopus with Diverse Strains of Borrelia burgdorferi, a Lyme Disease Agent. mBio 2012, 3, e00434-12. [Google Scholar] [CrossRef]
- Vuong, H.B.; Canham, C.D.; Fonseca, D.M.; Brisson, D.; Morin, P.J.; Smouse, P.E.; Ostfeld, R.S. Occurrence and Transmission Efficiencies of Borrelia burgdorferi ospC Types in Avian and Mammalian Wildlife. Infect. Genet. Evol. 2014, 27, 594–600. [Google Scholar] [CrossRef]
- Mechai, S.; Margos, G.; Feil, E.J.; Barairo, N.; Lindsay, L.R.; Michel, P.; Ogden, N.H. Evidence for Host-Genotype Associations of Borrelia burgdorferi Sensu Stricto. PLoS ONE 2016, 11, e0149345. [Google Scholar] [CrossRef]
- Combs, M.A.; Tufts, D.M.; Adams, B.; Lin, Y.-P.; Kolokotronis, S.-O.; Diuk-Wasser, M.A. Host Adaptation Drives Genetic Diversity in a Vector-Borne Disease System. PNAS Nexus 2023, 2, pgad234. [Google Scholar] [CrossRef] [PubMed]
- Brisson, D.; Baxamusa, N.; Schwartz, I.; Wormser, G.P. Biodiversity of Borrelia burgdorferi Strains in Tissues of Lyme Disease Patients. PLoS ONE 2011, 6, e22926. [Google Scholar] [CrossRef] [PubMed]
- Seinost, G.; Dykhuizen, D.E.; Dattwyler, R.J.; Golde, W.T.; Dunn, J.J.; Wang, I.-N.; Wormser, G.P.; Schriefer, M.E.; Luft, B.J. Four Clones of Borrelia burgdorferi Sensu Stricto Cause Invasive Infection in Humans. Infect. Immun. 1999, 67, 3518–3524. [Google Scholar] [CrossRef] [PubMed]
- Dykhuizen, D.E.; Brisson, D.; Sandigursky, S.; Wormser, G.P.; Nowakowski, J.; Nadelman, R.B.; Schwartz, I. The Propensity of Different Borrelia burgdorferi Sensu Stricto Genotypes to Cause Disseminated Infections in Humans. Am. J. Trop. Med. Hyg. 2008, 78, 806–810. [Google Scholar] [CrossRef]
- Wormser, G.P.; Brisson, D.; Liveris, D.; Hanincová, K.; Sandigursky, S.; Nowakowski, J.; Nadelman, R.B.; Ludin, S.; Schwartz, I. Borrelia burgdorferi Genotype Predicts the Capacity for Hematogenous Dissemination during Early Lyme Disease. J. Infect. Dis. 2008, 198, 1358–1364. [Google Scholar] [CrossRef]
- Vuong, H.B.; Chiu, G.S.; Smouse, P.E.; Fonseca, D.M.; Brisson, D.; Morin, P.J.; Ostfeld, R.S. Influences of Host Community Characteristics on Borrelia burgdorferi Infection Prevalence in Blacklegged Ticks. PLoS ONE 2017, 12, e0167810. [Google Scholar] [CrossRef]
- Lin, Y.P.; Tufts, D.M.; Combs, M.; Ii, A.P.D.; Marcinkiewicz, A.L.; Hirsbrunner, A.D.; Diaz, A.J.; Stout, J.L.; Blom, A.M.; Strle, K.; et al. Cellular and Immunological Mechanisms Influence Host-Adapted Phenotypes in a Vector-Borne Microparasite. Proc. R. Soc. B 2022, 289, 1–10. [Google Scholar] [CrossRef]
- Marcinkiewicz, A.L.; Brangulis, K.; Dupuis, A.P.; Hart, T.M.; Zamba-Campero, M.; Nowak, T.A.; Stout, J.L.; Akopjana, I.; Kazaks, A.; Bogans, J.; et al. Structural Evolution of an Immune Evasion Determinant Shapes Pathogen Host Tropism. Proc. Natl. Acad. Sci. USA 2023, 120, e2301549120. [Google Scholar] [CrossRef]
- de Taeye, S.W.; Kreuk, L.; van Dam, A.P.; Hovius, J.W.; Schuijt, T.J. Complement Evasion by Borrelia burgdorferi: It Takes Three to Tango. Trends Parasitol. 2013, 29, 119–128. [Google Scholar] [CrossRef]
- Kuo, M.M.; Lane, R.S.; Giclas, P.C. A Comparative Study of Mammalian and Reptilian Alternative Pathway of Complement-Mediated Killing of the Lyme Disease Spirochete (Borrelia burgdorferi). J. Parasitol. 2000, 86, 1223–1228. [Google Scholar] [CrossRef]
- van Dam, A.P.; Oei, A.; Jaspars, R.; Fijen, C.; Wilske, B.; Spanjaard, L.; Dankert, J. Complement-Mediated Serum Sensitivity among Spirochetes That Cause Lyme Disease. Infect. Immun. 1997, 65, 1228–1236. [Google Scholar] [CrossRef] [PubMed]
- Bhide, M.R.; Travnicek, M.; Levkutova, M.; Curlik, J.; Revajova, V.; Levkut, M. Sensitivity of Borrelia Genospecies to Serum Complement from Different Animals and Human: A Host–Pathogen Relationship. FEMS Immunol. Med. Microbiol. 2005, 43, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Brade, V.; Kleber, I.; Acker, G. Differences of Two Borrelia burgdorferi Strains in Complement Activation and Serum Resistance. Immunobiology 1992, 185, 453–465. [Google Scholar] [CrossRef] [PubMed]
- Breitner-Ruddock, S.; Würzner, R.; Schulze, J.; Brade, V. Heterogeneity in the Complement-Dependent Bacteriolysis within the Species of Borrelia burgdorferi. Med. Microbiol. Immunol. 1997, 185, 253–260. [Google Scholar] [CrossRef]
- Schuijt, T.J.; Hovius, J.W.R.; van Burgel, N.D.; Ramamoorthi, N.; Fikrig, E.; van Dam, A.P. The Tick Salivary Protein Salp15 Inhibits the Killing of Serum-Sensitive Borrelia burgdorferi Sensu Lato Isolates. Infect. Immun. 2008, 76, 2888–2894. [Google Scholar] [CrossRef]
- Rulison, E.L.; Kuczaj, I.; Pang, G.; Hickling, G.J.; Tsao, J.I.; Ginsberg, H.S. Flagging versus Dragging as Sampling Methods for Nymphal Ixodes scapularis (Acari: Ixodidae). J. Vector Ecol. 2013, 38, 163–167. [Google Scholar] [CrossRef]
- Oliver, J.H.; Kollars, T.M.; Chandler, F.W.; James, A.M.; Masters, E.J.; Lane, R.S.; Huey, L.O. First Isolation and Cultivation of Borrelia burgdorferi Sensu Lato from Missouri. J. Clin. Microbiol. 1998, 36, 1–5. [Google Scholar] [CrossRef]
- Pearson, P.; Skaltsis, O.; Luo, C.-Y.; Xu, G.; Oppler, Z.; Brisson, D.; Rich, S.M. A Borrelia burgdorferi Outer Surface Protein C (OspC) Genotyping Method Using Luminex Technology. PLoS ONE 2022, 17, e0269266. [Google Scholar] [CrossRef]
- Pearson, P.; Rich, C.; Feehan, M.J.R.; Ditchkoff, S.S.; Rich, S.M. White-Tailed Deer Serum Kills the Lyme Disease Spirochete, Borrelia burgdorferi. Vector-Borne Zoonotic Dis. 2023, 23, 303–305. [Google Scholar] [CrossRef]
- Nelson, D.R.; Rooney, S.; Miller, N.J.; Mather, T.N. Complement-Mediated Killing of Borrelia burgdorferi by Nonimmune Sera from Sika Deer. J. Parasitol. 2000, 86, 1232–1238. [Google Scholar] [CrossRef]
- Nowak, T.A.; Lown, L.A.; Marcinkiewicz, A.L.; Sürth, V.; Kraiczy, P.; Burke, R.; Lin, Y.-P. Outer Surface Protein E (OspE) Mediates Borrelia burgdorferi Sensu Stricto Strain-Specific Complement Evasion in the Eastern Fence Lizard, Sceloporus Undulatus. Ticks Tick-Borne Dis. 2023, 14, 102081. [Google Scholar] [CrossRef] [PubMed]
- Alitalo, A.; Meri, T.; Rämö, L.; Jokiranta, T.S.; Heikkilä, T.; Seppälä, I.J.T.; Oksi, J.; Viljanen, M.; Meri, S. Complement Evasion by Borrelia burgdorferi: Serum-Resistant Strains Promote C3b Inactivation. Infect. Immun. 2001, 69, 3685–3691. [Google Scholar] [CrossRef] [PubMed]
- Lachmann, P.J. Preparing Serum for Functional Complement Assays. J. Immunol. Methods 2010, 352, 195–197. [Google Scholar] [CrossRef] [PubMed]
- Lagal, V.; Portnoï, D.; Faure, G.; Postic, D.; Baranton, G. Borrelia burgdorferi Sensu Stricto Invasiveness Is Correlated with OspC–Plasminogen Affinity. Microbes Infect. 2006, 8, 645–652. [Google Scholar] [CrossRef]
- Önder, Ö.; Humphrey, P.T.; McOmber, B.; Korobova, F.; Francella, N.; Greenbaum, D.C.; Brisson, D. OspC Is Potent Plasminogen Receptor on Surface of Borrelia burgdorferi. J. Biol. Chem. 2012, 287, 16860–16868. [Google Scholar] [CrossRef]
- Bierwagen, P.; Szpotkowski, K.; Jaskolski, M.; Urbanowicz, A. Borrelia Outer Surface Protein C Is Capable of Human Fibrinogen Binding. FEBS J. 2019, 286, 2415–2428. [Google Scholar] [CrossRef]
- Caine, J.A.; Lin, Y.-P.; Kessler, J.R.; Sato, H.; Leong, J.M.; Coburn, J. Borrelia burgdorferi Outer Surface Protein C (OspC) Binds Complement Component C4b and Confers Bloodstream Survival. Cell. Microbiol. 2017, 19, e12786. [Google Scholar] [CrossRef]
- Lin, Y.-P.; Tan, X.; Caine, J.A.; Castellanos, M.; Chaconas, G.; Coburn, J.; Leong, J.M. Strain-Specific Joint Invasion and Colonization by Lyme Disease Spirochetes Is Promoted by Outer Surface Protein C. PLoS Pathog. 2020, 16, e1008516. [Google Scholar] [CrossRef]
- Carrasco, S.E.; Troxell, B.; Yang, Y.; Brandt, S.L.; Li, H.; Sandusky, G.E.; Condon, K.W.; Serezani, C.H.; Yang, X.F. Outer Surface Protein OspC Is an Antiphagocytic Factor That Protects Borrelia burgdorferi from Phagocytosis by Macrophages. Infect. Immun. 2015, 83, 4848–4860. [Google Scholar] [CrossRef]
- Mukherjee, P.G.; Liveris, D.; Hanincova, K.; Iyer, R.; Wormser, G.P.; Huang, W.; Schwartz, I. Borrelia burgdorferi Outer Surface Protein C Is Not the Sole Determinant of Dissemination in Mammals. Infect. Immun. 2023, 91, e00456-22. [Google Scholar] [CrossRef]
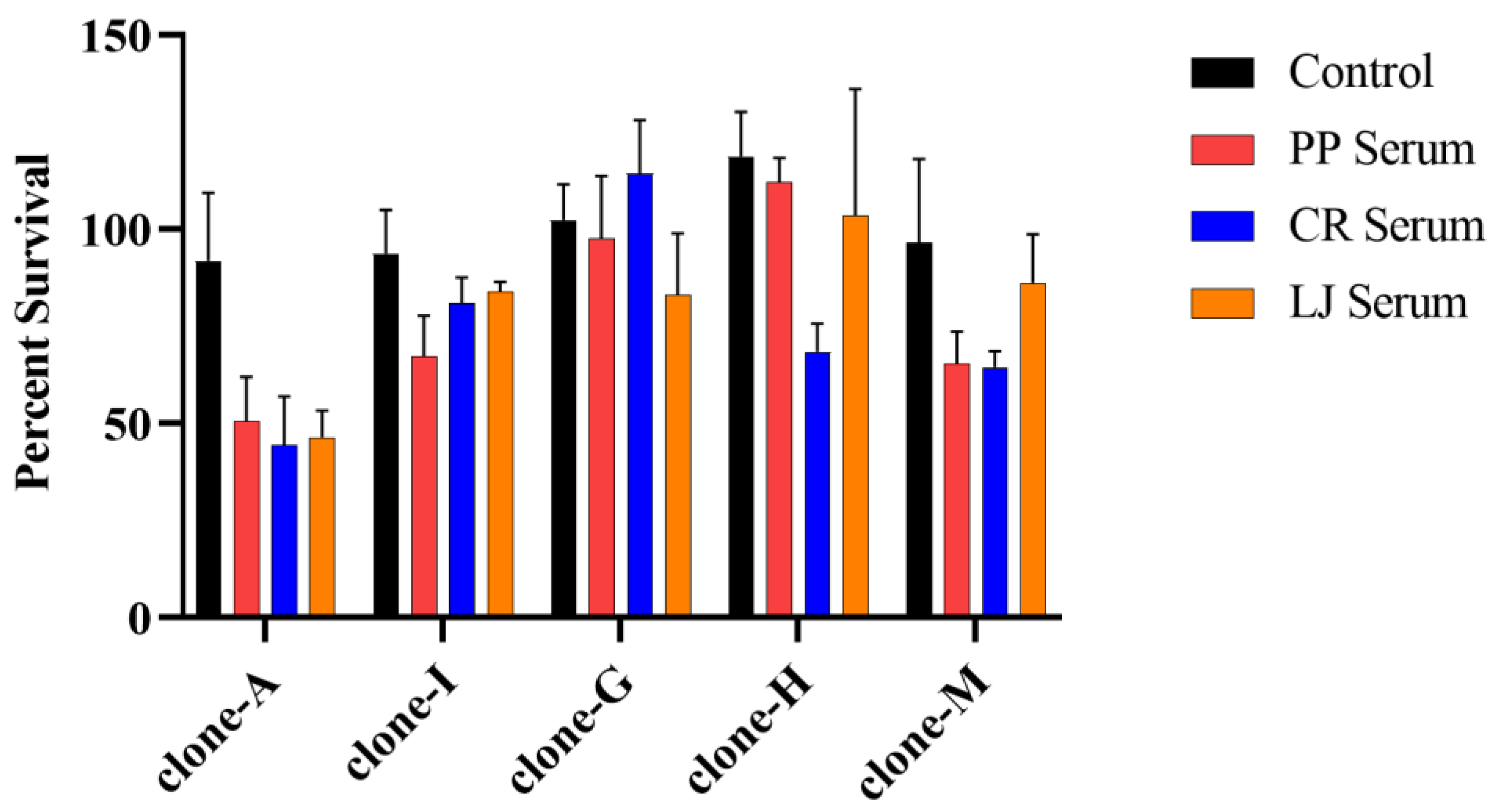

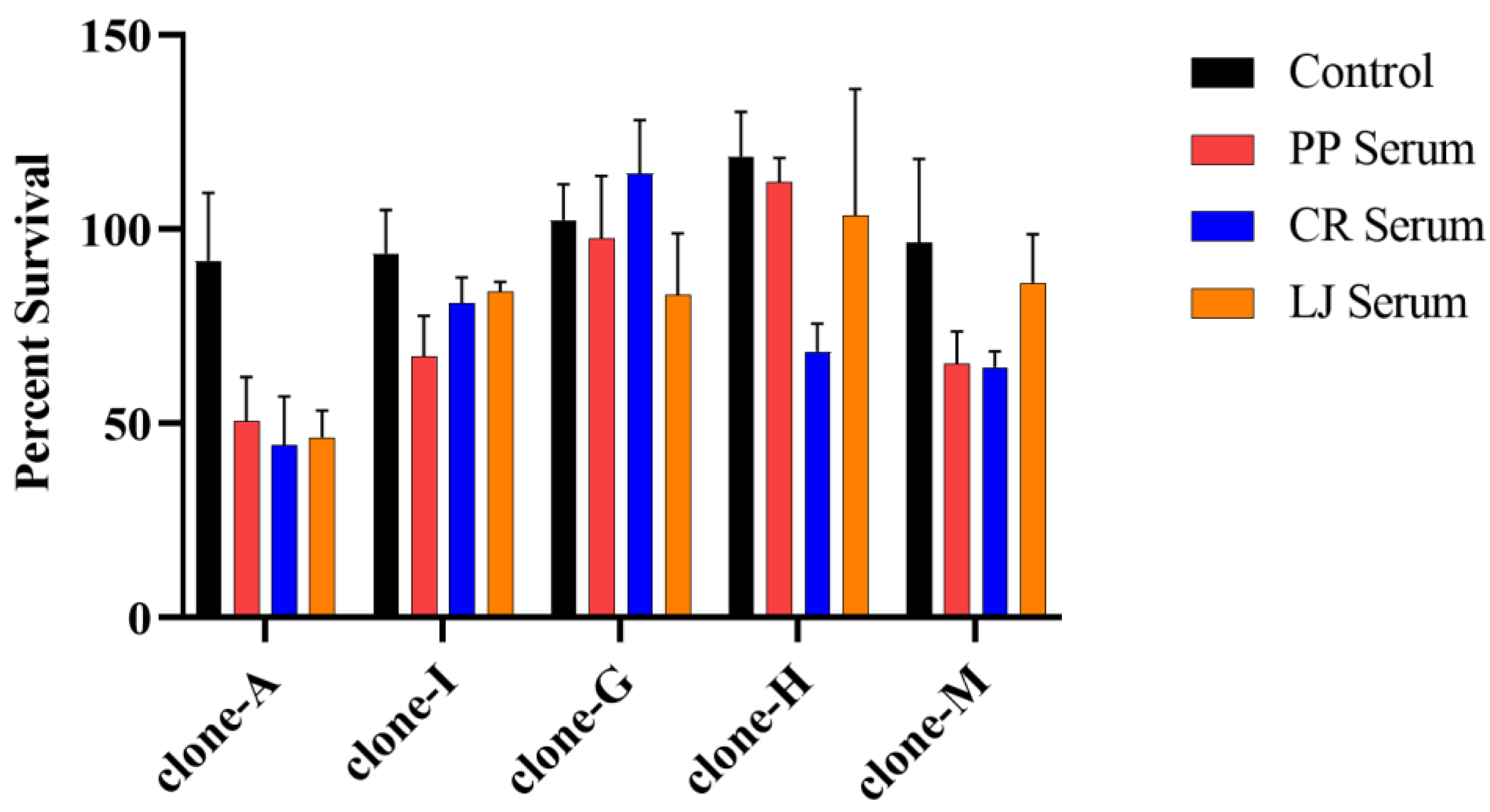{kind=link}
| Clone-A | Clone-I | Clone-G | Clone-H | Clone-M | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ctrl | PP | CR | LJ | Ctrl | PP | CR | LJ | Ctrl | PP | CR | LJ | Ctrl | PP | CR | LJ | Ctrl | PP | CR | LJ | ||
| clone-A | Ctrl | ** | *** | *** | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | |
| PP | ns | ns | *** | ns | ns | * | *** | ** | **** | * | **** | **** | ns | *** | ** | ns | ns | * | |||
| CR | ns | **** | ns | ** | ** | **** | **** | **** | ** | **** | **** | ns | **** | **** | ns | ns | *** | ||||
| LJ | **** | ns | * | ** | **** | *** | **** | ** | **** | **** | ns | **** | *** | ns | ns | ** | |||||
| clone-I | Ctrl | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | |||||
| PP | ns | ns | ns | ns | * | ns | * | * | ns | ns | ns | ns | ns | ns | |||||||
| CR | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ||||||||
| LJ | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | |||||||||
| clone-G | Ctrl | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | |||||||||
| PP | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | |||||||||||
| CR | ns | ns | ns | * | ns | ns | * | * | ns | ||||||||||||
| LJ | ns | ns | ns | ns | ns | ns | ns | ns | |||||||||||||
| clone-H | Ctrl | ns | * | ns | ns | ** | ** | ns | |||||||||||||
| PP | * | ns | ns | * | * | ns | |||||||||||||||
| CR | ns | ns | ns | ns | ns | ||||||||||||||||
| LJ | ns | ns | ns | ns | |||||||||||||||||
| clone-M | Ctrl | ns | ns | ns | |||||||||||||||||
| PP | ns | ns | |||||||||||||||||||
| CR | ns | ||||||||||||||||||||
| LJ | |||||||||||||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pearson, P.; Rich, C.; Siegel, E.L.; Brisson, D.; Rich, S.M. Differential Resistance of Borrelia burgdorferi Clones to Human Serum-Mediated Killing Does Not Correspond to Their Predicted Invasiveness. Pathogens 2023, 12, 1238. https://doi.org/10.3390/pathogens12101238
Pearson P, Rich C, Siegel EL, Brisson D, Rich SM. Differential Resistance of Borrelia burgdorferi Clones to Human Serum-Mediated Killing Does Not Correspond to Their Predicted Invasiveness. Pathogens. 2023; 12(10):1238. https://doi.org/10.3390/pathogens12101238
Chicago/Turabian StylePearson, Patrick, Connor Rich, Eric L. Siegel, Dustin Brisson, and Stephen M. Rich. 2023. "Differential Resistance of Borrelia burgdorferi Clones to Human Serum-Mediated Killing Does Not Correspond to Their Predicted Invasiveness" Pathogens 12, no. 10: 1238. https://doi.org/10.3390/pathogens12101238
APA StylePearson, P., Rich, C., Siegel, E. L., Brisson, D., & Rich, S. M. (2023). Differential Resistance of Borrelia burgdorferi Clones to Human Serum-Mediated Killing Does Not Correspond to Their Predicted Invasiveness. Pathogens, 12(10), 1238. https://doi.org/10.3390/pathogens12101238