
Toll-like Receptor Ligands Enhance Vaccine Efficacy against a Virulent Newcastle Disease Virus Challenge in Chickens

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
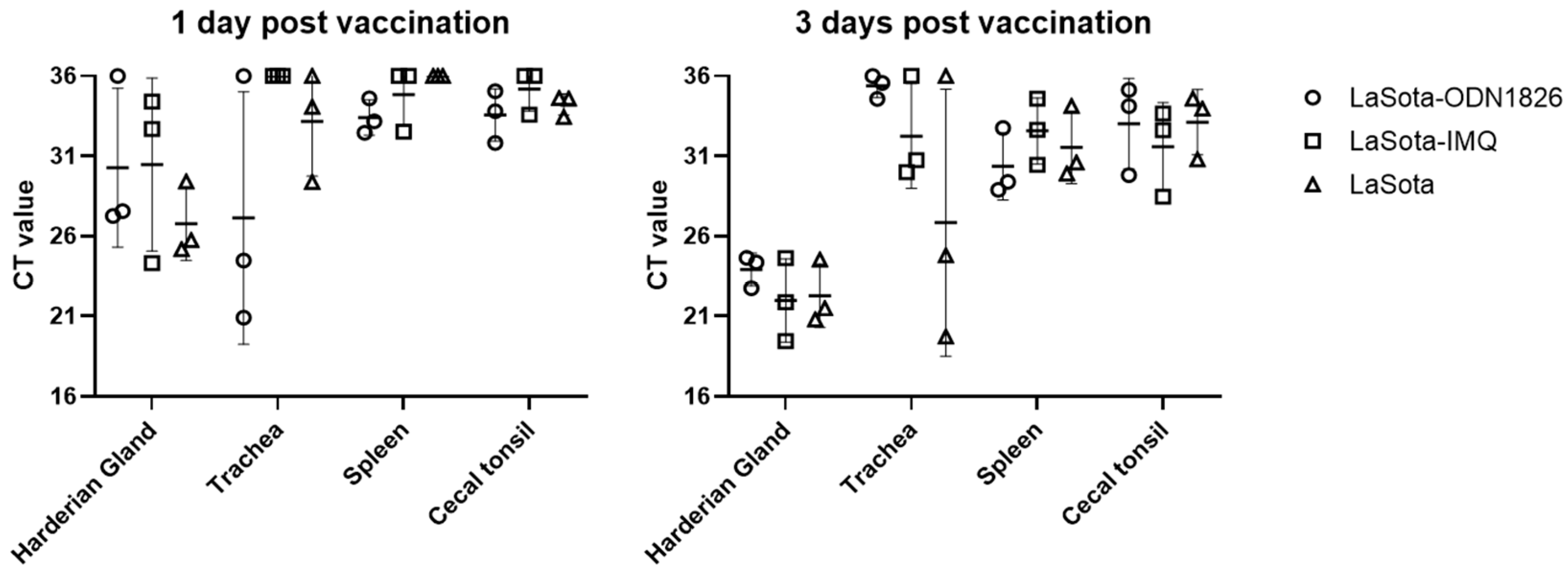
3. Results
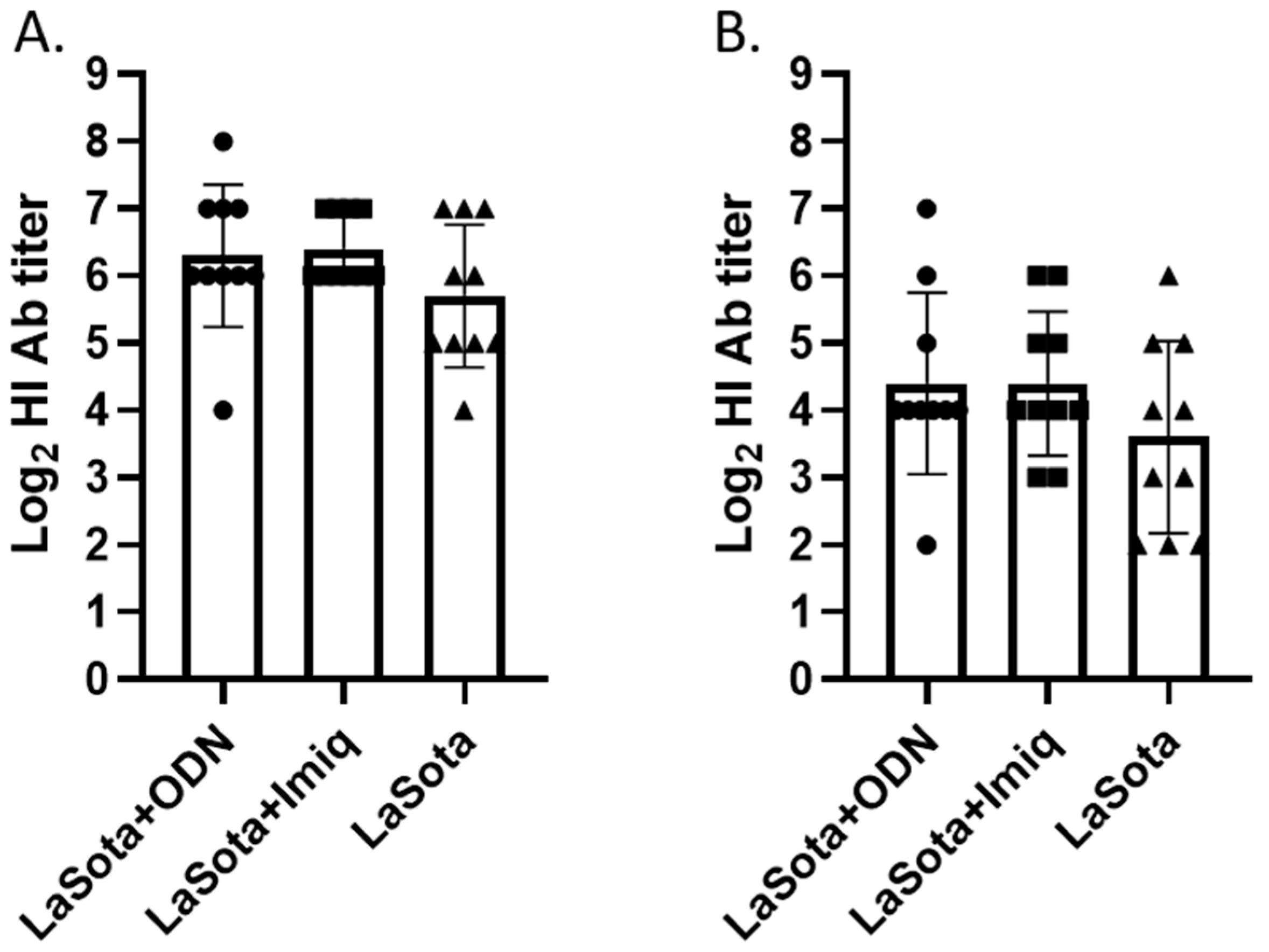
3.1. Hemagglutination Inhibition (HI) Antibody Titer at 2 Week Post-Vaccination
3.2. Morbidity and Mortality Post-Challenge
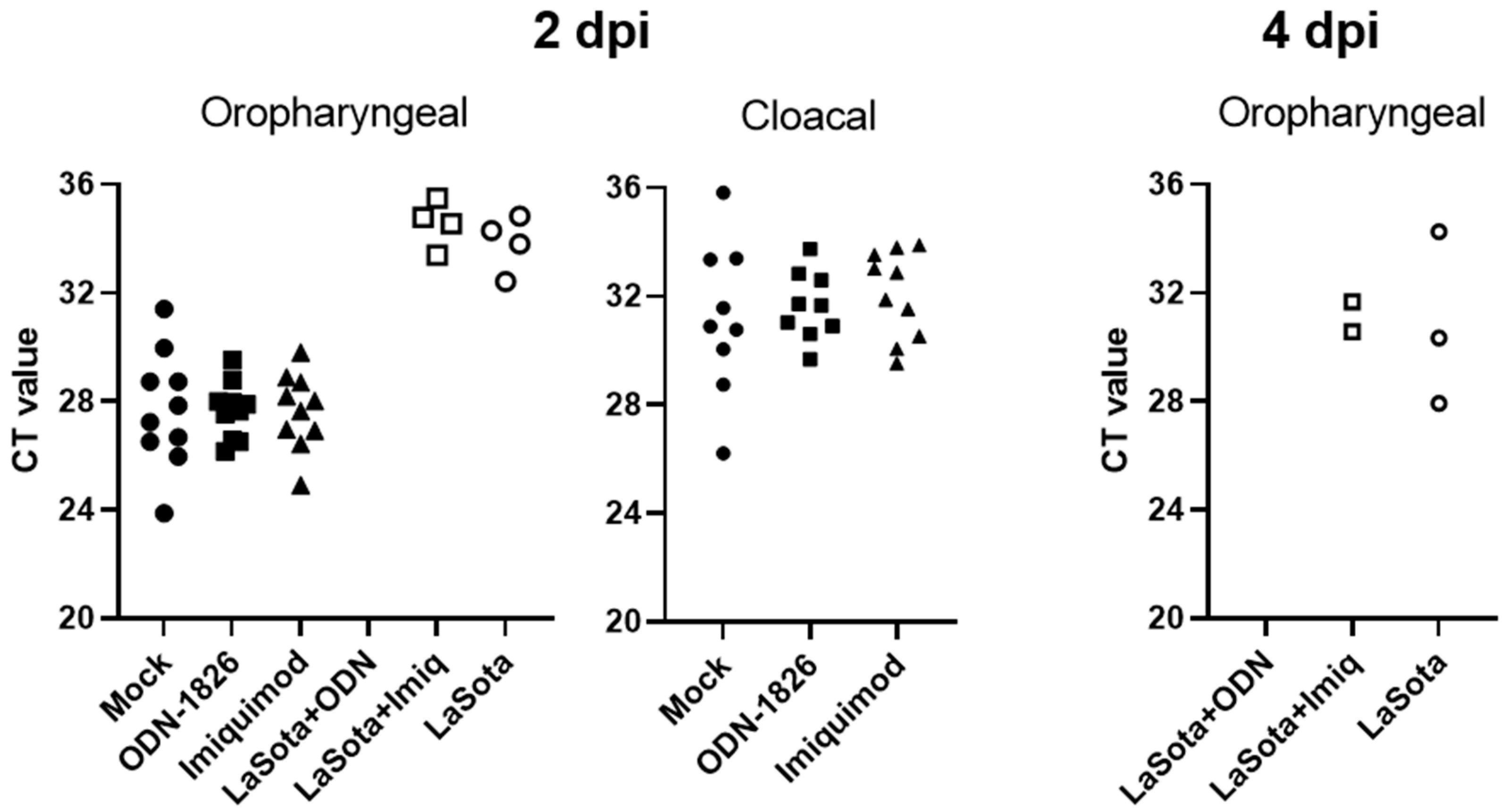
3.3. Viral RNA Detection from Oropharyneal and Cloacal Swabs
3.4. HI Antibody Titer at 2 Week Post-Challenge
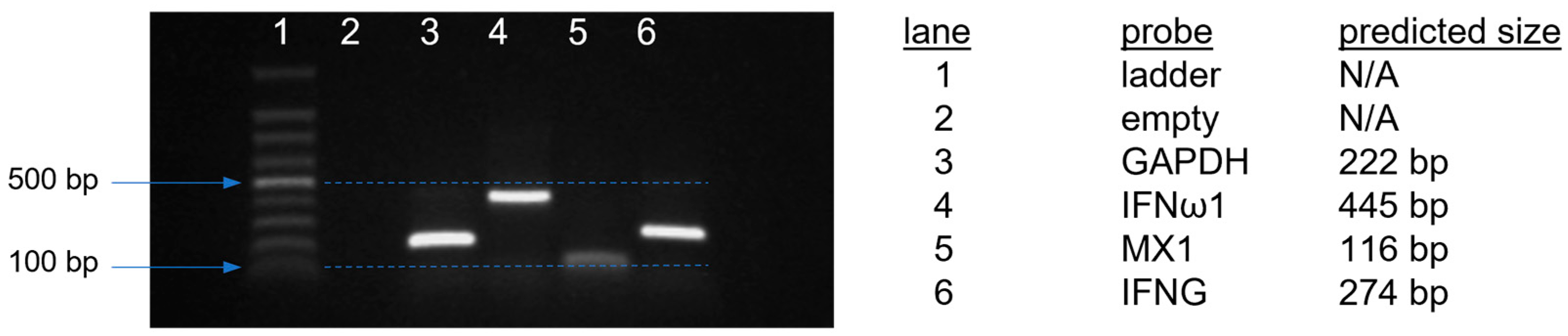
3.5. IFN and IFN-Stimulated Gene Expression after Adjuvant Treatment or Adjuvanted Vaccination
3.6. Vaccine Virus Detection from Tissues
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Organisation for Animal Health. Chapter 3.3.14.—Newcastle Disease (Infection with Newcastle Disease Virus). In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals, 12th ed.; World Organization for Animal Health: Paris, France, 2023; pp. 1–22. [Google Scholar]
- Suarez, D.L.; Miller, P.J.; Koch, G.; Mundt, E.; Rautenschlein, S. Newcastle Disease, Other Avian Paramyxoviruses, and Avian Metapneumovirus Infections. In Diseases of Poultry, 14th ed.; Swayne, D.E., Boulianne, M., Logue, C.M., McDougald, L.R., Nair, V., Suarez, D.L., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2020; pp. 111–166. [Google Scholar]
- Nezworski, J.; Gharaibeh, S. Avian Orthoavulavirus 1 Vaccination Failure in Minnesota Turkeys. Avian Dis. 2021, 65, 63–66. [Google Scholar] [CrossRef]
- Dimitrov, K.M.; Abolnik, C.; Afonso, C.L.; Albina, E.; Bahl, J.; Berg, M.; Briand, F.-X.; Brown, I.H.; Choi, K.-S.; Chvala, I. Updated unified phylogenetic classification system and revised nomenclature for Newcastle disease virus. Infect. Genet. Evol. 2019, 74, 103917–103932. [Google Scholar] [CrossRef]
- Miller, P.J.; Estevez, C.; Yu, Q.; Suarez, D.L.; King, D.J. Comparison of viral shedding following vaccination with inactivated and live Newcastle disease vaccines formulated with wild-type and recombinant viruses. Avian Dis. 2009, 53, 39–49. [Google Scholar] [CrossRef]
- Dimitrov, K.M.; Afonso, C.L.; Yu, Q.; Miller, P.J. Newcastle disease vaccines-A solved problem or a continuous challenge? Vet Microbiol. 2017, 206, 126–136. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on Toll-like receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef]
- Brownlie, R.; Allan, B. Avian toll-like receptors. Cell Tissue Res. 2011, 343, 121–130. [Google Scholar] [CrossRef]
- St Paul, M.; Brisbin, J.T.; Abdul-Careem, M.F.; Sharif, S. Immunostimulatory properties of Toll-like receptor ligands in chickens. Vet. Immunol. Immunopathol. 2013, 152, 191–199. [Google Scholar] [CrossRef]
- Kayraklioglu, N.; Horuluoglu, B.; Klinman, D.M. CpG oligonucleotides as vaccine adjuvants. Methods Mol. Biol. 2021, 2197, 51–85. [Google Scholar]
- Brownlie, R.; Zhu, J.; Allan, B.; Mutwiri, G.K.; Babiuk, L.A.; Potter, A.; Griebel, P. Chicken TLR21 acts as a functional homologue to mammalian TLR9 in the recognition of CpG oligodeoxynucleotides. Mol. Immunol. 2009, 46, 3163–3170. [Google Scholar] [CrossRef]
- Keestra, A.M.; de Zoete, M.R.; Bouwman, L.I.; van Putten, J.P. Chicken TLR21 is an innate CpG DNA receptor distinct from mammalian TLR9. J. Immunol. 2010, 185, 460–467. [Google Scholar] [CrossRef]
- Chuang, Y.C.; Tseng, J.C.; Yang, J.X.; Liu, Y.L.; Yeh, D.W.; Lai, C.Y.; Yu, G.Y.; Hsu, L.C.; Huang, C.M.; Chuang, T.H. Toll-like receptor 21 of chicken and duck recognize a broad array of immunostimulatory CpG-oligodeoxynucleotide sequences. Vaccines 2020, 8, 639. [Google Scholar] [CrossRef]
- Vleugels, B.; Ververken, C.; Goddeeris, B.M. Stimulatory effect of CpG sequences on humoral response in chickens. Poult Sci. 2002, 81, 1317–1321. [Google Scholar] [CrossRef]
- Gomis, S.; Babiuk, L.; Godson, D.L.; Allan, B.; Thrush, T.; Townsend, H.; Willson, P.; Waters, E.; Hecker, R.; Potter, A. Protection of chickens against Escherichia coli infections by DNA containing CpG motifs. Infect. Immun. 2003, 71, 857–863. [Google Scholar] [CrossRef]
- St Paul, M.; Mallick, A.I.; Read, L.R.; Villanueva, A.I.; Parvizi, P.; Abdul-Careem, M.F.; Nagy, É.; Sharif, S. Prophylactic treatment with Toll-like receptor ligands enhances host immunity to avian influenza virus in chickens. Vaccine 2012, 30, 4524–4531. [Google Scholar] [CrossRef] [PubMed]
- Raj, S.; Alizadeh, M.; Shoojadoost, B.; Hodgins, D.; Nagy, É.; Mubareka, S.; Karimi, K.; Behboudi, S.; Sharif, S. Determining the protective efficacy of Toll-like receptor ligands to minimize H9N2 avian influenza virus transmission in chickens. Viruses 2023, 15, 238. [Google Scholar] [CrossRef]
- Barjesteh, N.; Shojadoost, B.; Brisbin, J.T.; Emam, M.; Hodgins, D.C.; Nagy, É.; Sharif, S. Reduction of avian influenza virus shedding by administration of Toll-like receptor ligands to chickens. Vaccine 2015, 33, 4843–4849. [Google Scholar] [CrossRef]
- Liang, J.; Fu, J.; Kang, H.; Lin, J.; Yu, Q.; Yang, Q. Comparison of 3 kinds of Toll-like receptor ligands for inactivated avian H5N1 influenza virus intranasal immunization in chicken. Poult. Sci. 2013, 92, 2651–2660. [Google Scholar] [CrossRef]
- Talebi, A.; Arky-Rezai, S. Efficacy of CpG-ODN administration routes on humoral responses against Newcastle disease in broilers. Arch. Razi Inst. 2019, 74, 357–364. [Google Scholar]
- Zhang, L.; Zhang, M.; Li, J.; Cao, T.; Tian, X.; Zhou, F. Enhancement of mucosal immune responses by intranasal co-delivery of Newcastle disease vaccine plus CpG oligonucleotide in SPF chickens in vivo. Res. Vet. Sci. 2008, 85, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Linghua, Z.; Xingshan, T.; Fengzhen, Z. Vaccination with Newcastle disease vaccine and CpG oligodeoxynucleotides induces specific immunity and protection against Newcastle disease virus in SPF chicken. Vet. Immunol Immunopathol. 2007, 115, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Nigar, S.; Shimosato, T. Cooperation of oligodeoxynucleotides and synthetic molecules as enhanced immune modulators. Front. Nutr. 2019, 6, 140. [Google Scholar] [CrossRef] [PubMed]
- Dockrell, D.; Kinghorn, G. Imiquimod and resiquimod as novel immunomodulators. J. Antimicrob. Chemother. 2001, 48, 751–755. [Google Scholar] [CrossRef] [PubMed]
- Tachibana, T.; Takeda, K.; Khan, M.S.I.; Makino, R.; Cline, M.A. Effects of toll-like receptor-7 agonists on feeding behaviour, voluntary activity, cloacal temperature and crop emptying in chicks. Br. Poult. Sci. 2020, 61, 303–310. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, M.R.; Xia, J.; Smith, A.L.; Magor, K.E. The duck Toll like receptor 7: Genomic organization, expression and function. Mol. Immunol. 2008, 45, 2055–2061. [Google Scholar] [CrossRef]
- Qi, Y.; Chen, S.; Zhao, Q.; Wang, M.; Jia, R.; Zhu, D.; Liu, M.; Liu, F.; Chen, X.; Ceng, A. Molecular cloning, tissue distribution, and immune function of goose TLR7. Immunol. Lett. 2015, 163, 135–142. [Google Scholar] [CrossRef]
- Tang, Y.; Lu, J.; Wu, P.; Liu, Z.; Tian, Z.; Zha, G.; Chen, H.; Wang, Q.; Wang, Q.; Hou, F.; et al. Inactivated vaccine with adjuvants consisting of pattern recognition receptor agonists confers protection against avian influenza viruses in chickens. Vet. Microbiol. 2014, 172, 120–128. [Google Scholar] [CrossRef]
- Zhang, A.; Li, D.; Song, C.; Jing, H.; Li, H.; Mi, J.; Zhang, G.; Jin, S.; Ren, X.; Huangfu, H.; et al. Evaluation of different combination of pam2CSK4, poly (I:C) and imiquimod enhance immune responses to H9N2 avian influenza antigen in dendritic cells and duck. PLoS ONE. 2022, 17, e0271746. [Google Scholar] [CrossRef]
- Annamalai, A.; Ramakrishnan, S.; Sachan, S.; Sharma, B.K.; Anand Kumar, B.S.; Kumar, V.; Badasara, S.K.; Kumar, A.; Saravanan, B.C.; Krishnaswamy, N. Administration of TLR7 agonist, resiquimod, in different types of chicken induces a mixed Th1 and Th2 response in the peripheral blood mononuclear cells. Res. Vet. Sci. 2015, 100, 105–108. [Google Scholar] [CrossRef]
- Gupta, A.; Deka, P.; Kumar, S. Resiquimod inhibits Newcastle disease virus replication by modulating host cytokines: An understanding towards its possible therapeutics. Cytokine 2020, 125, 154811. [Google Scholar] [CrossRef]
- De Silva Senapathi, U.; Aboelkhair, M.; Puro, K.; Ali, M.; Amarasinghe, A.; Abdul-Cader, M.S.; Van Marle, G.; Czub, M.; Abdul-Careem, M.F. In ovo delivered Toll-like receptor 7 ligand, resiquimod enhances host responses against infectious bronchitis corona virus (IBV) Infection. Vaccines 2020, 8, 186. [Google Scholar] [CrossRef]
- Matoo, J.J.; Bashir, K.; Kumar, A.; Krishnaswamy, N.; Dey, S.; Chellappa, M.M.; Ramakrishnan, S. Resiquimod enhances mucosal and systemic immunity against avian infectious bronchitis virus vaccine in the chicken. Microb. Pathog. 2018, 119, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Annamalai, A.; Ramakrishnan, S.; Sachan, S.; Kumar, B.S.A.; Sharma, B.K.; Kumar, V.; Palanivelu, M.; Varghese, B.P.; Kumar, A.; Saravanan, B.C.; et al. Prophylactic potential of resiquimod against very virulent infectious bursal disease virus (vvIBDV) challenge in the chicken. Vet. Microbiol. 2016, 187, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Sachan, S.; Ramakrishnan, S.; Annamalai, A.; Sharma, B.K.; Malik, H.; Saravanan, B.C.; Jain, L.; Saxena, M.; Kumar, A.; Krishnaswamy, N. Adjuvant potential of resiquimod with inactivated Newcastle disease vaccine and its mechanism of action in chicken. Vaccine 2015, 33, 4526–4532. [Google Scholar] [CrossRef] [PubMed]
- Sharma, B.K.; Ramakrishnan, S.; Kaliappan, A.; Singh, M.; Kumar, A.; Dandapat, S.; Dey, S.; Chellappa, M.M. Evaluation of a lipopolysaccharide and resiquimod combination as an adjuvant with inactivated Newcastle disease virus vaccine in chickens. Vaccines 2022, 10, 894. [Google Scholar] [CrossRef]
- Spackman, E.; Stephens, C. Virus Isolation and Propagation in Embryonating Eggs. In A Laboratory Manual for The Isolation, Identification and Characterization of Avian Pathogens, 6th ed.; Williams, S.M., Dufour-Zavala, L., Jackwood, M.W., Lee, M.D., Lupiani, B., Reed, W.M., Spackman, E., Woolcock, P.R., Eds.; American Association of Avian Pathologists: Jacksonville, FL, USA, 2016; pp. 361–367. [Google Scholar]
- Villegas, P. Titration of Biological Suspensions. In A Laboratory Manual for The Isolation, Identification and Characterization of Avian Pathogens, 6th ed.; Williams, S.M., Dufour-Zavala, L., Jackwood, M.W., Lee, M.D., Lupiani, B., Reed, W.M., Spackman, E., Woolcock, P.R., Eds.; American Association of Avian Pathologists: Jacksonville, FL, USA, 2016; pp. 355–360. [Google Scholar]
- Wise, M.G.; Suarez, D.L.; Seal, B.S.; Pedersen, J.C.; Senne, D.A.; King, D.J.; Kapczynski, D.R.; Spackman, E. Development of a real-time reverse-transcription PCR for detection of Newcastle disease virus RNA in clinical samples. J. Clin. Microbiol. 2004, 42, 329–338. [Google Scholar] [CrossRef]
- Ferreira, H.L.; Taylor, T.L.; Dimitrov, K.M.; Sabra, M.; Afonso, C.L.; Suarez, D.L. Virulent Newcastle disease viruses from chicken origin are more pathogenic and transmissible to chickens than viruses normally maintained in wild birds. Vet. Microbiol. 2019, 235, 25–34. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Lee, C.W.; Kc, M.; Ngunjiri, J.M.; Ghorbani, A.; Lee, K. TLR3 and MDA5 knockout DF-1 cells enhance replication of avian orthoavulavirus 1. Avian Dis. 2023, 67, 94–101. [Google Scholar] [CrossRef]
- Patel, B.A.; Gomis, S.; Dar, A.; Willson, P.J.; Babiuk, L.A.; Potter, A.; Mutwiri, G.; Tikoo, S.K. Oligodeoxynucleotides containing CpG motifs (CpG-ODN) predominantly induce Th1-type immune response in neonatal chicks. Dev. Comp. Immunol. 2008, 32, 1041–1049. [Google Scholar] [CrossRef]
- Nan, F.L.; Zhang, H.; Nan, W.L.; Xie, C.Z.; Ha, Z.; Chen, X.; Xu, X.H.; Qian, J.; Qiu, X.S.; Ge, J.Y.; et al. Lentogenic NDV V protein inhibits IFN responses and represses cell apoptosis. Vet. Microbiol. 2021, 261, 109181. [Google Scholar] [CrossRef]
- Liu, W.; Xu, Z.; Qiu, Y.; Qiu, X.; Tan, L.; Song, C.; Sun, Y.; Liao, Y.; Liu, X.; Ding, C. Single-cell transcriptome atlas of Newcastle disease virus in chickens both in vitro and in vivo. Microbiol. Spectr. 2023, 11, e0512122. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Vaccine | Treatment Adjuvant |
|---|---|---|
| 1. PBS control | none | none |
| 2. Adjuvant only (ODN-1826) | none | ODN-1826 |
| 3. Adjuvant only (Imiquimod) | none | Imiquimod |
| 4. Vaccine + Adjuvant (ODN-1826) | LaSota | ODN-1826 |
| 5. Vaccine + Adjuvant (Imiquimod) | LaSota | Imiquimod |
| 6. Vaccine only | LaSota | none |
| Primer Name | Primer Sequence | Reference 1 |
|---|---|---|
| GAPDH_F5 | 5′-CTGAATGGGAAGCTTACTGGAATG-3′ | NM_204305 |
| GAPDH_R5 | 5′-CGCATCAAAGGTGGAGGAATG-3′ | |
| IFNω1_F4 | 5′-ACAAGAAGCAAGCAGCCATC-3′ | NM_001024836 |
| IFNω1_R4 | 5′-GTGCGGTCAATCCAGTGTTT-3′ | |
| MX1_F6 | 5′-CCAGACCTAGTGAACGAAGGAA-3′ | XM_046901195.1 |
| MX1_R6 | 5′-CAGAAGTCCATTTGCCCATAACA-3′ | |
| IFNG_F1 | 5′-GTAGCTGACGGTGGACCTATT-3′ | NM_205149 |
| IFNG_R1 | 5’-AGAGTTCATTCGCGGCTTTG-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, C.-W.; Bakre, A.; Olivier, T.L.; Alvarez-Narvaez, S.; Harrell, T.L.; Conrad, S.J. Toll-like Receptor Ligands Enhance Vaccine Efficacy against a Virulent Newcastle Disease Virus Challenge in Chickens. Pathogens 2023, 12, 1230. https://doi.org/10.3390/pathogens12101230
Lee C-W, Bakre A, Olivier TL, Alvarez-Narvaez S, Harrell TL, Conrad SJ. Toll-like Receptor Ligands Enhance Vaccine Efficacy against a Virulent Newcastle Disease Virus Challenge in Chickens. Pathogens. 2023; 12(10):1230. https://doi.org/10.3390/pathogens12101230
Chicago/Turabian StyleLee, Chang-Won, Abhijeet Bakre, Timothy L. Olivier, Sonsiray Alvarez-Narvaez, Telvin L. Harrell, and Steven J. Conrad. 2023. "Toll-like Receptor Ligands Enhance Vaccine Efficacy against a Virulent Newcastle Disease Virus Challenge in Chickens" Pathogens 12, no. 10: 1230. https://doi.org/10.3390/pathogens12101230
APA StyleLee, C.-W., Bakre, A., Olivier, T. L., Alvarez-Narvaez, S., Harrell, T. L., & Conrad, S. J. (2023). Toll-like Receptor Ligands Enhance Vaccine Efficacy against a Virulent Newcastle Disease Virus Challenge in Chickens. Pathogens, 12(10), 1230. https://doi.org/10.3390/pathogens12101230