
The In Vitro Antimicrobial Susceptibility of Borrelia burgdorferi sensu lato: Shedding Light on the Known Unknowns


Abstract
1. Introduction
2. The Causative Agents
3. Clinical Manifestations
4. In Vitro Susceptibility Testing Methods for B. burgdorferi s.l.
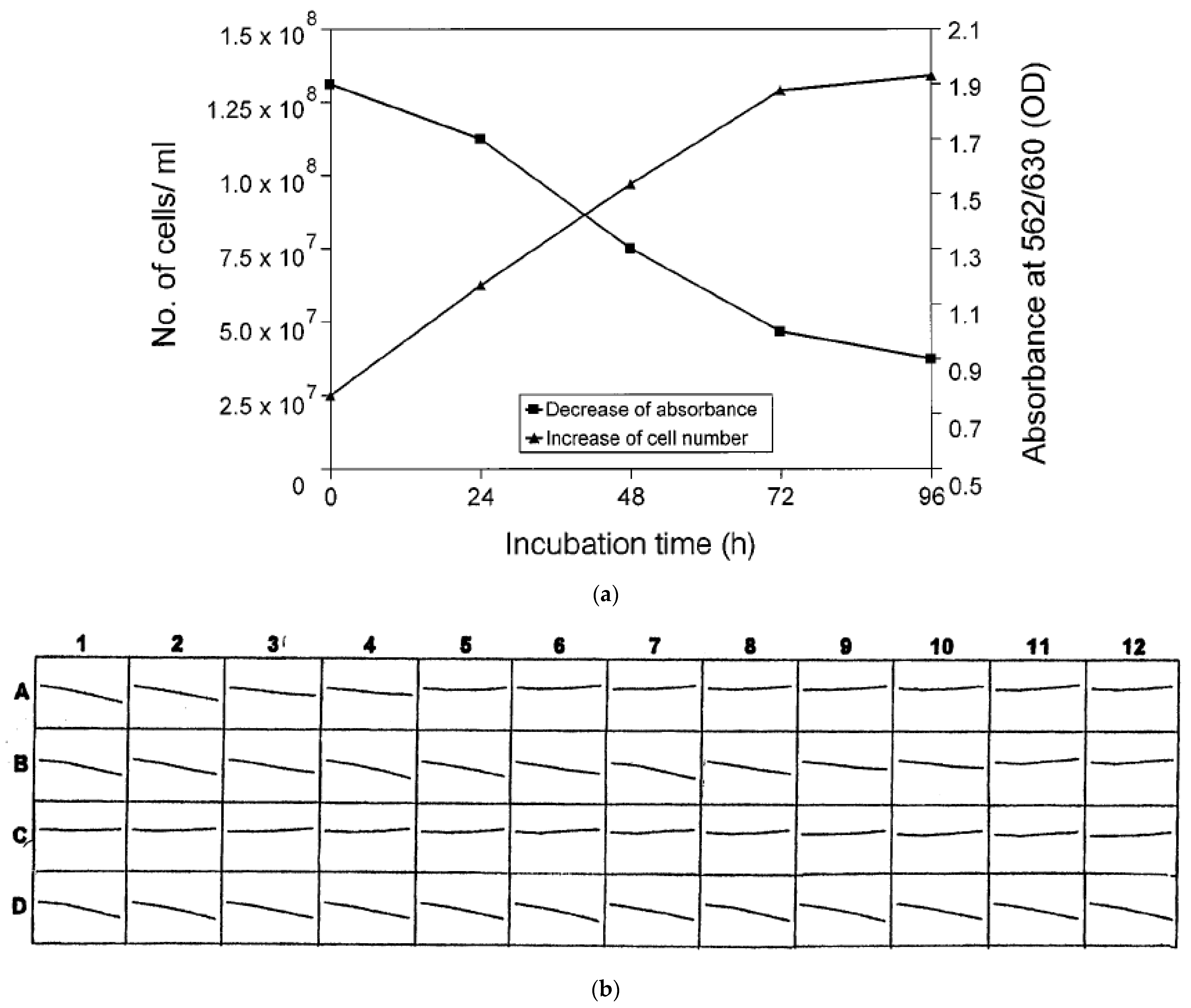
4.1. Determination of Minimal Inhibitory Concentrations (MIC)
4.2. Microdilution in Combination with Microscopic MIC Determination
4.3. Microdilution in Combination with Colorimetric MIC Determination
4.4. Approaches for MBC Determination Using Macro- and Microdilution Methods
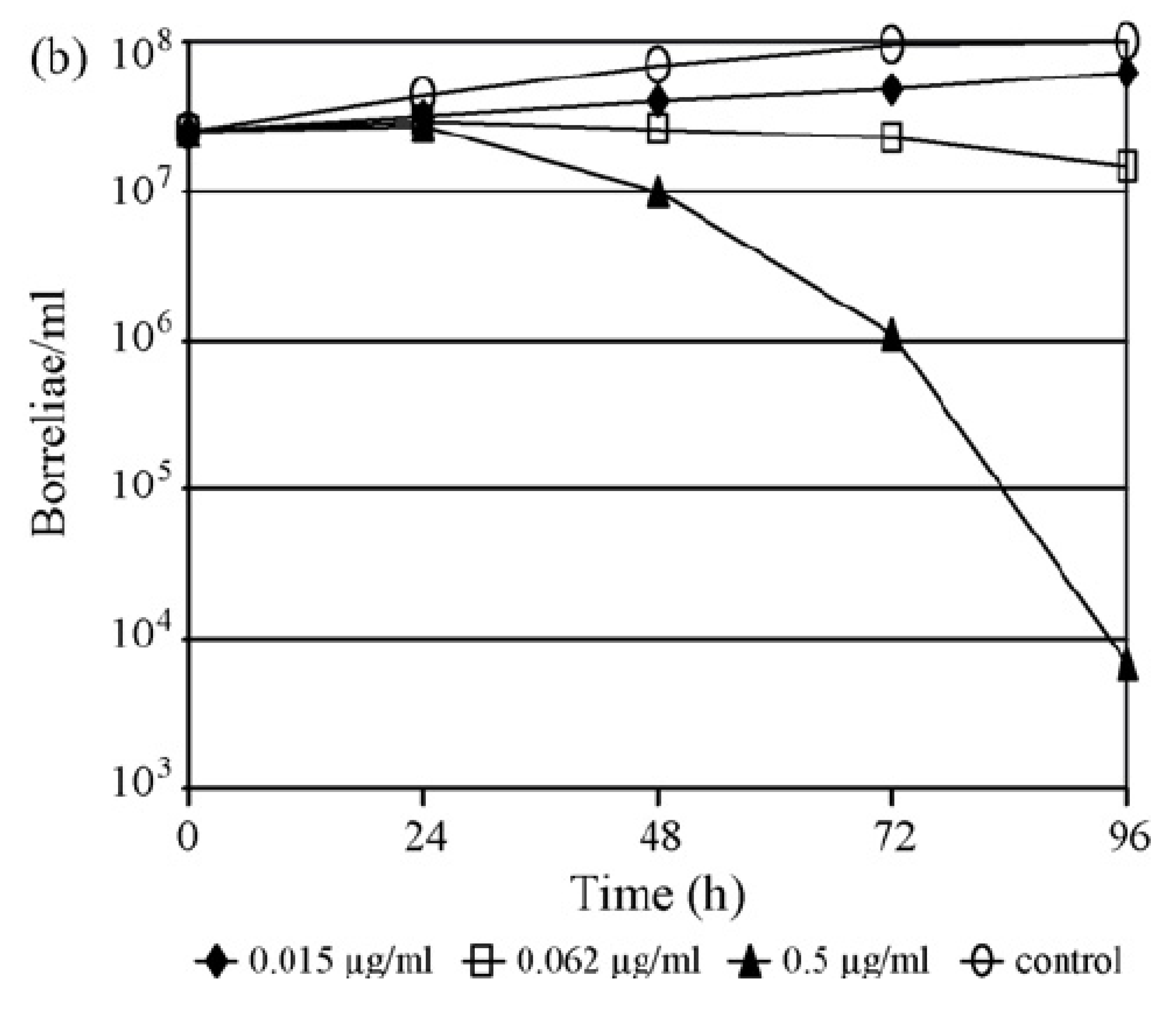
4.5. Time-Kill Experiments Using Macro- and Microdilution Methods
4.6. Time-Kill Experiments Using the Subsurface Plating Technique (SPT)
4.7. Approaches for MIC Determination Using a Dialyses Culture Method
5. In Vitro Antimicrobial Susceptibility Pattern of the B. burgdorferi Complex
5.1. Spectrum of In Vitro Susceptibility against Antimicrobial Agents
5.2. ß-Lactam Agents
5.3. Macrolides, Azalides and Ketolides
5.4. Tetracyclines and Glycylcyclines
5.5. Fluoroquinolones
5.6. Aminoglycosides, Glycopeptides, Streptogramins, Fusidic Acid and Nalidixic Acid
6. Differences in the Antimicrobial Susceptibility of Borrelia burgdorferi Genospecies
7. Drawbacks and Challenges for Susceptibility Testing of Borreliae In Vitro
Discrepancies in In Vitro and In Vivo Activities: Interactions of Antimicrobials with BSK Medium
8. Possible Mechanisms of Resistance of Borrelia burgdorferi s.l. In Vitro
8.1. Currently Known Phenotypic and Genotypic Mechanisms of Resistance
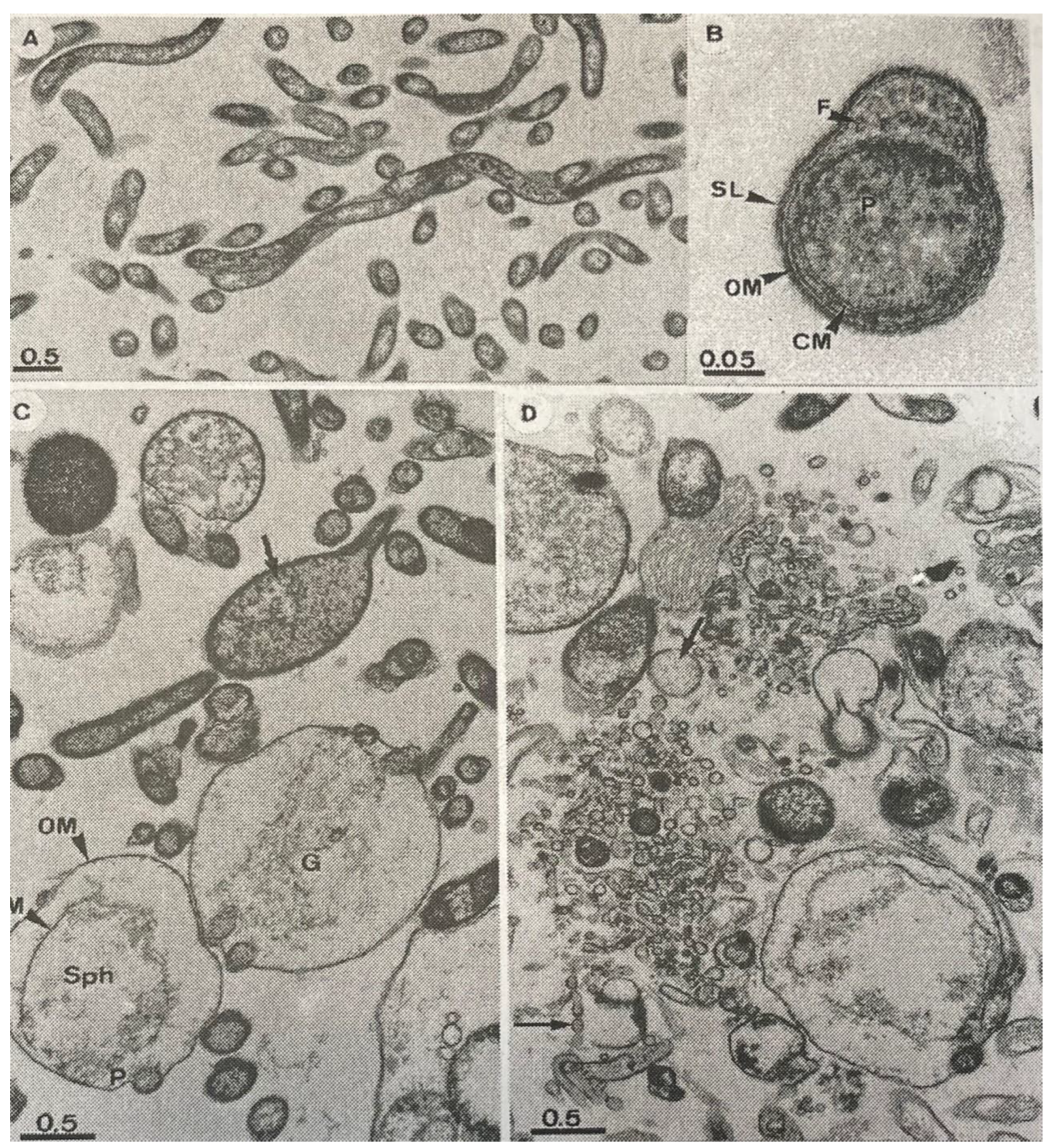
8.2. Tolerance of Borrelia burgdorferi s.l. When Exposed to Antimicrobial Agents
8.3. Changes in Protein Expression in Borreliae Exposed to Antimicrobial Agents
8.4. Evidence from Interactions between Borreliae and Antimicrobial Agents and Eukaryotic Cells
9. Evidence from Isolates from Patients and Primates with Spirochetal Persistence
10. Summary and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Steere, A.C. Lyme disease. N. Engl. J. Med. 2001, 345, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Burgdorfer, W.; Barbour, A.G.; Hayes, S.F.; Benach, J.L.; Grunwaldt, E.; Davis, J.P. Lyme disease-a tick-borne spirochetosis? Science 1982, 216, 1317–1319. [Google Scholar] [CrossRef] [PubMed]
- Gern, L.; Aeschlimann, A.; Schwan, T.G.; Lane, R.S. A tribute to Dr. Willy Burgdorfer, Medical Entomologist Extraordinaire. Ticks Tick Borne Dis. 2016, 7, 66–67. [Google Scholar] [CrossRef]
- CDC. Lyme disease—United States, 2000. MMWR Morb. Mortal. Wkly. Rep. 2002, 51, 29–31. [Google Scholar]
- O’Connell, S.; Granström, M.; Gray, J.S.; Stanek, G. Epidemiology of European Lyme borreliosis. Zentralbl. Bakteriol. 1998, 287, 229–240. [Google Scholar] [CrossRef]
- Stanek, G.; O’Connell, S.; Cimmino, M.; Aberer, E.; Kristoferitsch, W.; Granström, M.; Guy, E.; Gray, J. European Union concerted action on risk assessment in Lyme borreliosis: Clinical case definitions for Lyme borreliosis. Wien. Klin. Wochenschr. 1996, 108, 741–747. [Google Scholar]
- Hunfeld, K.-P.; Ruzic-Sabljic, E.; Norris, D.E.; Kraiczy, P.; Strle, F. In vitro susceptibility testing of Borrelia burgdorferi sensu lato isolates cultured from patients with erythema migrans before and after antimicrobial chemotherapy. Antimicrob. Agents Chemother. 2005, 49, 1294–1301. [Google Scholar] [CrossRef]
- Sykes, R.A.; Makiello, P. An estimate of Lyme borreliosis incidence in Western Europe. J. Public Health 2017, 39, 74–81. [Google Scholar] [CrossRef]
- Huppertz, H.I.; Böhme, M.; Standaert, S.M.; Karch, H.; Plotkin, S.A. Incidence of Lyme borreliosis in the Würzburg region of Germany. Eur. J. Clin. Microbiol. Infect. Dis. 1999, 18, 697–703. [Google Scholar] [CrossRef]
- Nelson, C.A.; Saha, S.; Kugeler, K.J.; Delorey, M.J.; Shankar, M.B.; Hinckley, A.F.; Mead, P.S. Incidence of clinician-diagnosed Lyme disease, United States, 2005–2010. Emerg. Infect. Dis. 2015, 21, 1625–1631. [Google Scholar] [CrossRef]
- Müller, I.; Freitag, M.H.; Poggensee, G.; Scharnetzky, E.; Straube, E.; Schoerner, C.; Hlobil, H.; Hagedorn, H.-J.; Stanek, G.; Schubert-Unkmeir, A.; et al. Evaluating frequency, diagnostic quality, and cost of Lyme borreliosis testing in Germany: A retrospective model analysis. Clin. Dev. Immunol. 2012, 2012, 595427. [Google Scholar] [CrossRef] [PubMed]
- Svartz, N. Penicillin behandling vid dermatitis atrophicans Herxheimer. Nord. Med. 1941, 32, 2783. [Google Scholar]
- Thyresson, N. The penicillin treatment of acrodermatitis atrophicans chronica (Herxheimer). Acta Derm. Venereol. 1949, 29, 572–621. [Google Scholar]
- Steere, A.C.; Grodzicki, R.L.; Kornblatt, A.N.; Craft, J.E.; Barbour, A.G.; Burgdorfer, W.; Schmid, G.P.; Johnson, E.; Malawista, S.E. The spirochetal etiology of Lyme disease. N. Engl. J. Med. 1983, 308, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Oschmann, P.; Kaiser, R. Therapy and prognosis. In Lyme Borreliosis and Tick-Borne Encephalitis; Oschmann, P., Ed.; UNI-MED-Verl.: Bremen, Germany, 1999; ISBN 3895994499. [Google Scholar]
- Wormser, G.P.; Nadelman, R.B.; Dattwyler, R.J.; Dennis, D.T.; Shapiro, E.D.; Steere, A.C.; Rush, T.J.; Rahn, D.W.; Coyle, P.K.; Persing, D.H.; et al. Practice guidelines for the treatment of Lyme disease. The Infectious Diseases Society of America. Clin. Infect. Dis. 2000, 31 (Suppl. S1), S1–S14. [Google Scholar] [CrossRef] [PubMed]
- Hunfeld, K.-P.; Kraiczy, P.; Kekoukh, E.; Schäfer, V.; Brade, V. Standardised in vitro susceptibility testing of Borrelia burgdorferi against well-known and newly developed antimicrobial agents—Possible implications for new therapeutic approaches to Lyme disease. Int. J. Med. Microbiol. 2002, 291 (Suppl. S33), S125–S137. [Google Scholar] [CrossRef] [PubMed]
- Preac-Mursic, V.; Weber, K.; Pfister, H.W.; Wilske, B.; Gross, B.; Baumann, A.; Prokop, J. Survival of Borrelia burgdorferi in antibiotically treated patients with Lyme borreliosis. Infection 1989, 17, 355–359. [Google Scholar] [CrossRef]
- Hassler, D.; Zöller, L.; Haude, M.; Hufnagel, H.D.; Heinrich, F.; Sonntag, H.G. Cefotaxime versus penicillin in the late stage of Lyme disease—Prospective, randomized therapeutic study. Infection 1990, 18, 16–20. [Google Scholar] [CrossRef]
- Pfister, H.W.; Preac-Mursic, V.; Wilske, B.; Schielke, E.; Sörgel, F.; Einhäupl, K.M. Randomized comparison of ceftriaxone and cefotaxime in Lyme neuroborreliosis. J. Infect. Dis. 1991, 163, 311–318. [Google Scholar] [CrossRef]
- Strle, F.; Preac-Mursic, V.; Cimperman, J.; Ruzic, E.; Maraspin, V.; Jereb, M. Azithromycin versus doxycycline for treatment of erythema migrans: Clinical and microbiological findings. Infection 1993, 21, 83–88. [Google Scholar] [CrossRef]
- Nowakowski, J.; McKenna, D.; Nadelman, R.B.; Cooper, D.; Bittker, S.; Holmgren, D.; Pavia, C.; Johnson, R.C.; Wormser, G.P. Failure of treatment with cephalexin for Lyme disease. Arch. Fam. Med. 2000, 9, 563–567. [Google Scholar] [CrossRef]
- Burgdorfer, W.; Lane, R.S.; Barbour, A.G.; Gresbrink, R.A.; Anderson, J.R. The western black-legged tick, Ixodes pacificus: A vector of Borrelia burgdorferi. Am. J. Trop. Med. Hyg. 1985, 34, 925–930. [Google Scholar] [CrossRef]
- Ružić-Sabljić, E.; Cerar, T. Progress in the molecular diagnosis of Lyme disease. Expert Rev. Mol. Diagn. 2017, 17, 19–30. [Google Scholar] [CrossRef]
- Stanek, G.; Reiter, M. The expanding Lyme Borrelia complex—Clinical significance of genomic species? Clin. Microbiol. Infect. 2011, 17, 487–493. [Google Scholar] [CrossRef] [PubMed]
- Margos, G.; Hepner, S.; Fingerle, V. Characteristics of Borrelia burgdorferi sensu lato. In Lyme Borreliosis; Hunfeld, K.-P., Gray, J., Eds.; Springer International Publishing: Cham, Switzerland, 2022; pp. 1–29. ISBN 978-3-030-93679-2. [Google Scholar]
- Margos, G.; Sing, A.; Fingerle, V. Published data do not support the notion that Borrelia valaisiana is human pathogenic. Infection 2017, 45, 567–569. [Google Scholar] [CrossRef] [PubMed]
- Margos, G.; Lane, R.S.; Fedorova, N.; Koloczek, J.; Piesman, J.; Hojgaard, A.; Sing, A.; Fingerle, V. Borrelia bissettiae sp. nov. and Borrelia californiensis sp. nov. prevail in diverse enzootic transmission cycles. Int. J. Syst. Evol. Microbiol. 2016, 66, 1447–1452. [Google Scholar] [CrossRef]
- Pritt, B.S.; Respicio-Kingry, L.B.; Sloan, L.M.; Schriefer, M.E.; Replogle, A.J.; Bjork, J.; Liu, G.; Kingry, L.C.; Mead, P.S.; Neitzel, D.F.; et al. Borrelia mayonii sp. nov., a member of the Borrelia burgdorferi sensu lato complex, detected in patients and ticks in the upper midwestern United States. Int. J. Syst. Evol. Microbiol. 2016, 66, 4878–4880. [Google Scholar] [CrossRef] [PubMed]
- Golovchenko, M.; Vancová, M.; Clark, K.; Oliver, J.H.; Grubhoffer, L.; Rudenko, N. A divergent spirochete strain isolated from a resident of the southeastern United States was identified by multilocus sequence typing as Borrelia bissettii. Parasit. Vectors 2016, 9, 68. [Google Scholar] [CrossRef]
- Fingerle, V.; Schulte-Spechtel, U.C.; Ruzic-Sabljic, E.; Leonhard, S.; Hofmann, H.; Weber, K.; Pfister, K.; Strle, F.; Wilske, B. Epidemiological aspects and molecular characterization of Borrelia burgdorferi s.l. from southern Germany with special respect to the new species Borrelia spielmanii sp. nov. Int. J. Med. Microbiol. 2008, 298, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Lohr, B.; Fingerle, V.; Norris, D.E.; Hunfeld, K.-P. Laboratory diagnosis of Lyme borreliosis: Current state of the art and future perspectives. Crit. Rev. Clin. Lab. Sci. 2018, 55, 219–245. [Google Scholar] [CrossRef]
- Stanek, G.; Strle, F. The history, epidemiology, clinical manifestations and treatment of Lyme borreliosis. In Lyme Borreliosis; Hunfeld, K.-P., Gray, J., Eds.; Springer International Publishing: Cham, Switzerland, 2022; pp. 77–105. ISBN 978-3-030-93679-2. [Google Scholar]
- Marcinkiewicz, A.L.; Dupuis, A.P.; Zamba-Campero, M.; Nowak, N.; Kraiczy, P.; Ram, S.; Kramer, L.D.; Lin, Y.-P. Blood treatment of Lyme borreliae demonstrates the mechanism of CspZ-mediated complement evasion to promote systemic infection in vertebrate hosts. Cell. Microbiol. 2019, 21, e12998. [Google Scholar] [CrossRef] [PubMed]
- Wilske, B.; Preac-Mursic, V.; Göbel, U.B.; Graf, B.; Jauris, S.; Soutschek, E.; Schwab, E.; Zumstein, G. An OspA serotyping system for Borrelia burgdorferi based on reactivity with monoclonal antibodies and OspA sequence analysis. J. Clin. Microbiol. 1993, 31, 340–350. [Google Scholar] [CrossRef] [PubMed]
- Boerner, J.; Failing, K.; Wittenbrink, M.M. In vitro antimicrobial susceptibility testing of Borrelia burgdorferi: Influence of test conditions on minimal inhibitory concentration (MIC) values. Zentralbl. Bakteriol. 1995, 283, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Preac-Mursic, V.; Wilske, B.; Schierz, G. European Borrelia burgdorferi isolated from humans and ticks culture conditions and antibiotic susceptibility. Zentralbl. Bakteriol. Mikrobiol. Hyg. A 1986, 263, 112–118. [Google Scholar] [CrossRef]
- Mursic, V.P.; Wilske, B.; Schierz, G.; Holmburger, M.; Süss, E. In vitro and in vivo susceptibility of Borrelia burgdorferi. Eur. J. Clin. Microbiol. 1987, 6, 424–426. [Google Scholar] [CrossRef]
- Johnson, R.C.; Kodner, C.B.; Jurkovich, P.J.; Collins, J.J. Comparative in vitro and in vivo susceptibilities of the Lyme disease spirochete Borrelia burgdorferi to cefuroxime and other antimicrobial agents. Antimicrob. Agents Chemother. 1990, 34, 2133–2136. [Google Scholar] [CrossRef]
- Dever, L.L.; Jorgensen, J.H.; Barbour, A.G. In vitro antimicrobial susceptibility testing of Borrelia burgdorferi: A microdilution MIC method and time-kill studies. J. Clin. Microbiol. 1992, 30, 2692–2697. [Google Scholar] [CrossRef]
- Agger, W.A.; Callister, S.M.; Jobe, D.A. In vitro susceptibilities of Borrelia burgdorferi to five oral cephalosporins and ceftriaxone. Antimicrob. Agents Chemother. 1992, 36, 1788–1790. [Google Scholar] [CrossRef]
- Dever, L.L.; Jorgensen, J.H.; Barbour, A.G. In vitro activity of vancomycin against the spirochete Borrelia burgdorferi. Antimicrob. Agents Chemother. 1993, 37, 1115–1121. [Google Scholar] [CrossRef]
- Levin, J.M.; Nelson, J.A.; Segreti, J.; Harrison, B.; Benson, C.A.; Strle, F. In vitro susceptibility of Borrelia burgdorferi to 11 antimicrobial agents. Antimicrob. Agents Chemother. 1993, 37, 1444–1446. [Google Scholar] [CrossRef]
- Baradaran-Dilmaghani, R.; Stanek, G. In vitro susceptibility of thirty Borrelia strains from various sources against eight antimicrobial chemotherapeutics. Infection 1996, 24, 60–63. [Google Scholar] [CrossRef] [PubMed]
- Schorter, L.; Straubinger, R.K. Impact of four local anaesthetics on growth and viability of in vitro cultured Borrelia burgdorferi sensu stricto, Borrelia bavariensis and Borrelia afzelii. Ticks Tick Borne Dis. 2021, 12, 101735. [Google Scholar] [CrossRef] [PubMed]
- Veinović, G.; Cerar, T.; Strle, F.; Lotrič-Furlan, S.; Maraspin, V.; Cimperman, J.; Ružić-Sabljić, E. In vitro susceptibility of European human Borrelia burgdorferi sensu stricto strains to antimicrobial agents. Int. J. Antimicrob. Agents 2013, 41, 288–291. [Google Scholar] [CrossRef] [PubMed]
- Caol, S.; Divers, T.; Crisman, M.; Chang, Y.-F. In vitro susceptibility of Borrelia burgdorferi isolates to three antibiotics commonly used for treating equine Lyme disease. BMC Vet. Res. 2017, 13, 293. [Google Scholar] [CrossRef] [PubMed]
- Koetsveld, J.; Draga, R.O.P.; Wagemakers, A.; Manger, A.; Oei, A.; Visser, C.E.; Hovius, J.W. In Vitro Susceptibility of the Relapsing-Fever Spirochete Borrelia miyamotoi to Antimicrobial Agents. Antimicrob. Agents Chemother. 2017, 61, 10–1128. [Google Scholar] [CrossRef]
- Koetsveld, J.; Manger, A.; Hoornstra, D.; Draga, R.O.; Oei, A.; Kolyasnikova, N.M.; Toporkova, M.G.; Sarksyan, D.S.; Wagemakers, A.; Platonov, A.E.; et al. In Vitro Antimicrobial Susceptibility of Clinical Isolates of Borrelia miyamotoi. Antimicrob. Agents Chemother. 2018, 62, 10–1128. [Google Scholar] [CrossRef]
- Dever, L.L.; Jorgensen, J.H.; Barbour, A.G. Comparative in vitro activities of clarithromycin, azithromycin, and erythromycin against Borrelia burgdorferi. Antimicrob. Agents Chemother. 1993, 37, 1704–1706. [Google Scholar] [CrossRef]
- Hunfeld, K.P.; Kraiczy, P.; Wichelhaus, T.A.; Schäfer, V.; Brade, V. New colorimetric microdilution method for in vitro susceptibility testing of Borrelia burgdorferi against antimicrobial substances. Eur. J. Clin. Microbiol. Infect. Dis. 2000, 19, 27–32. [Google Scholar] [CrossRef]
- Hunfeld, K.-P. Contributions to seroepidemiology, diagnosis, and antimicrobial susceptibility of Borrelia, Ehrlichia, and Babesia as indigenous tick conducted pathogens; Shaker: Aachen, Germany, 2004; ISBN 978-3-8322-3347-1. [Google Scholar]
- Hunfeld, K.-P.; Brade, V. Antimicrobial susceptibility of Borrelia burgdorferi sensu lato: What we know, what we don’t know, and what we need to know. Wien. Klin. Wochenschr. 2006, 118, 659–668. [Google Scholar] [CrossRef]
- Maurin, M. Borrelia. In Antibiogram; Courvalin, P., Leclercq, R., Rice, L., Eds.; ESKA Publ. ASM Press: Portland, OR, USA, 2010; pp. 618–624. ISBN 978-2747215459. [Google Scholar]
- Hunfeld, K.P.; Kraiczy, P.; Wichelhaus, T.A.; Schäfer, V.; Brade, V. Colorimetric in vitro susceptibility testing of penicillins, cephalosporins, macrolides, streptogramins, tetracyclines, and aminoglycosides against Borrelia burgdorferi isolates. Int. J. Antimicrob. Agents 2000, 15, 11–17. [Google Scholar] [CrossRef]
- Morgenstern, K.; Baljer, G.; Norris, D.E.; Kraiczy, P.; Hanssen-Hübner, C.; Hunfeld, K.-P. In vitro susceptibility of Borrelia spielmanii to antimicrobial agents commonly used for treatment of Lyme disease. Antimicrob. Agents Chemother. 2009, 53, 1281–1284. [Google Scholar] [CrossRef]
- Kraiczy, P.; Weigand, J.; Wichelhaus, T.A.; Heisig, P.; Backes, H.; Schäfer, V.; Acker, G.; Brade, V.; Hunfeld, K.P. In vitro activities of fluoroquinolones against the spirochete Borrelia burgdorferi. Antimicrob. Agents Chemother. 2001, 45, 2486–2494. [Google Scholar] [CrossRef]
- Ates, L.; Hanssen-Hübner, C.; Norris, D.E.; Richter, D.; Kraiczy, P.; Hunfeld, K.-P. Comparison of in vitro activities of tigecycline, doxycycline, and tetracycline against the spirochete Borrelia burgdorferi. Ticks Tick Borne Dis. 2010, 1, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Hunfeld, K.-P.; Wichelhaus, T.A.; Rödel, R.; Acker, G.; Brade, V.; Kraiczy, P. Comparison of in vitro activities of ketolides, macrolides, and an azalide against the spirochete Borrelia burgdorferi. Antimicrob. Agents Chemother. 2004, 48, 344–347. [Google Scholar] [CrossRef] [PubMed]
- Hunfeld, K.-P.; Ruzić-Sabljić, E.; Norris, D.E.; Kraiczy, P.; Strle, F. Risk of culture-confirmed borrelial persistence in patients treated for erythema migrans and possible mechanisms of resistance. Int. J. Med. Microbiol. 2006, 296 (Suppl. S40), S233–S241. [Google Scholar] [CrossRef]
- Hansen, K.; Hovmark, A.; Lebech, A.M.; Lebech, K.; Olsson, I.; Halkier-Sørensen, L.; Olsson, E.; Asbrink, E. Roxithromycin in Lyme borreliosis: Discrepant results of an in vitro and in vivo animal susceptibility study and a clinical trial in patients with erythema migrans. Acta Derm. Venereol. 1992, 72, 297–300. [Google Scholar] [CrossRef] [PubMed]
- Weber, K. Treatment failure in erythema migrans—A review. Infection 1996, 24, 73–75. [Google Scholar] [CrossRef]
- Strle, F.; Cheng, Y.; Cimperman, J.; Maraspin, V.; Lotric-Furlan, S.; Nelson, J.A.; Picken, M.M.; Ruzic-Sabljic, E.; Picken, R.N. Persistence of Borrelia burgdorferi sensu lato in resolved erythema migrans lesions. Clin. Infect. Dis. 1995, 21, 380–389. [Google Scholar] [CrossRef]
- Luft, B.J.; Volkman, D.J.; Halperin, J.J.; Dattwyler, R.J. New chemotherapeutic approaches in the treatment of Lyme borreliosis. Ann. N. Y. Acad. Sci. 1988, 539, 352–361. [Google Scholar] [CrossRef]
- Hunfeld, K.P.; Weigand, J.; Wichelhaus, T.A.; Kekoukh, E.; Kraiczy, P.; Brade, V. In vitro activity of mezlocillin, meropenem, aztreonam, vancomycin, teicoplanin, ribostamycin and fusidic acid against Borrelia burgdorferi. Int. J. Antimicrob. Agents 2001, 17, 203–208. [Google Scholar] [CrossRef]
- Alder, J.; Mitten, M.; Jarvis, K.; Gupta, P.; Clement, J. Efficacy of clarithromycin for treatment of experimental Lyme disease in vivo. Antimicrob. Agents Chemother. 1993, 37, 1329–1333. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Berger, B.W.; Kaplan, M.H.; Rothenberg, I.R.; Barbour, A.G. Isolation and characterization of the Lyme disease spirochete from the skin of patients with erythema chronicum migrans. J. Am. Acad. Dermatol. 1985, 13, 444–449. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.C.; Schmid, G.P.; Hyde, F.W.; Steigerwalt, A.G.; Brenner, D.J. Borrelia burgdorferi sp. nov.: Etiologic Agent of Lyme Disease. Int. J. Syst. Bacteriol. 1984, 34, 496–497. [Google Scholar] [CrossRef]
- Leimer, N.; Wu, X.; Imai, Y.; Morrissette, M.; Pitt, N.; Favre-Godal, Q.; Iinishi, A.; Jain, S.; Caboni, M.; Leus, I.V.; et al. A selective antibiotic for Lyme disease. Cell 2021, 184, 5405–5418.e16. [Google Scholar] [CrossRef] [PubMed]
- Morshed, M.G.; Konishi, H.; Nishimura, T.; Nakazawa, T. Evaluation of agents for use in medium for selective isolation of Lyme disease and relapsing fever Borrelia species. Eur. J. Clin. Microbiol. Infect. Dis. 1993, 12, 512–518. [Google Scholar] [CrossRef] [PubMed]
- Stiernstedt, S.H.; Tadesse, T.; Wretlind, B. Dialysis culture for determination of MIC and MBC of benzylpenicillin against Borrelia burgdorferi. APMIS 1999, 107, 380–382. [Google Scholar] [CrossRef]
- Sharma, B.; Brown, A.V.; Matluck, N.E.; Hu, L.T.; Lewis, K. Borrelia burgdorferi, the causative agent of Lyme disease, forms drug-tolerant persister cells. Antimicrob. Agents Chemother. 2015, 59, 4616–4624. [Google Scholar] [CrossRef]
- Bayer, A.S.; Lam, K. Efficacy of vancomycin plus rifampin in experimental aortic-valve endocarditis due to methicillin-resistant Staphylococcus aureus: In vitro-in vivo correlations. J. Infect. Dis. 1985, 151, 157–165. [Google Scholar] [CrossRef]
- Chandrasekar, P.H.; Crane, L.R.; Bailey, E.J. Comparison of the activity of antibiotic combinations in vitro with clinical outcome and resistance emergence in serious infection by Pseudomonas aeruginosa in non-neutropenic patients. J. Antimicrob. Chemother. 1987, 19, 321–329. [Google Scholar] [CrossRef]
- Preac Mursic, V.; Marget, W.; Busch, U.; Pleterski Rigler, D.; Hagl, S. Kill kinetics of Borrelia burgdorferi and bacterial findings in relation to the treatment of Lyme borreliosis. Infection 1996, 24, 9–16. [Google Scholar] [CrossRef]
- Callister, S.M.; Schell, R.F.; Lovrich, S.D. Lyme disease assay which detects killed Borrelia burgdorferi. J. Clin. Microbiol. 1991, 29, 1773–1776. [Google Scholar] [CrossRef] [PubMed]
- Rödel, R.; Freyer, A.; Bittner, T.; Schäfer, V.; Hunfeld, K.-P. In vitro activities of faropenem, ertapenem, imipenem and meropenem against Borrelia burgdorferi s.l. Int. J. Antimicrob. Agents 2007, 30, 83–86. [Google Scholar] [CrossRef] [PubMed]
- Preac-Mursic, V.; Wilske, B.; Reinhardt, S. Culture of Borrelia burgdorferi on six solid media. Eur. J. Clin. Microbiol. Infect. Dis. 1991, 10, 1076–1079. [Google Scholar] [CrossRef] [PubMed]
- Stiernstedt, S.H.; Wretlind, B. Dialysis culture enables more accurate determination of MIC of benzylpenicillin for Borrelia burgdorferi than does conventional procedure. Antimicrob. Agents Chemother. 1996, 40, 2882–2883. [Google Scholar] [CrossRef] [PubMed]
- Dever, L.L.; Jorgensen, J.H.; Barbour, A.G. In vitro susceptibility testing of Borrelia burgdorferi by a dialysis culture method. Antimicrob. Agents Chemother. 1997, 41, 1208–1209. [Google Scholar] [CrossRef]
- Hunfeld, K.-P.; Rödel, R.; Wichelhaus, T.A. In vitro activity of eight oral cephalosporins against Borrelia burgdorferi. Int. J. Antimicrob. Agents 2003, 21, 313–318. [Google Scholar] [CrossRef]
- Strle, F.; Maraspin, V.; Lotric-Furlan, S.; Ruzić-Sabljić, E.; Cimperman, J. Azithromycin and doxycycline for treatment of Borrelia culture-positive erythema migrans. Infection 1996, 24, 64–68. [Google Scholar] [CrossRef]
- Sicklinger, M.; Wienecke, R.; Neubert, U. In vitro susceptibility testing of four antibiotics against Borrelia burgdorferi: A comparison of results for the three genospecies Borrelia afzelii, Borrelia garinii, and Borrelia burgdorferi sensu stricto. J. Clin. Microbiol. 2003, 41, 1791–1793. [Google Scholar] [CrossRef]
- Sohaskey, C.D.; Barbour, A.G. Esterases in serum-containing growth media counteract chloramphenicol acetyltransferase activity in vitro. Antimicrob. Agents Chemother. 1999, 43, 655–660. [Google Scholar] [CrossRef]
- Capobianco, J.O.; Cao, Z.; Shortridge, V.D.; Ma, Z.; Flamm, R.K.; Zhong, P. Studies of the novel ketolide ABT-773: Transport, binding to ribosomes, and inhibition of protein synthesis in Streptococcus pneumoniae. Antimicrob. Agents Chemother. 2000, 44, 1562–1567. [Google Scholar] [CrossRef]
- Balfour, J.A.; Figgitt, D.P. Telithromycin. Drugs 2001, 61, 815–829. [Google Scholar] [CrossRef] [PubMed]
- Dougherty, T.J.; Barrett, J.F. ABT-773: A new ketolide antibiotic. Expert Opin. Investig. Drugs 2001, 10, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Hunfeld, K.P.; Wichelhaus, T.A.; Kekoukh, E.; Molitor, M.; Kraiczy, P.; Brade, V. In vitro susceptibility of the Borrelia burgdorferi sensu lato complex to ABT-773, a novel ketolide. J. Antimicrob. Chemother. 2001, 48, 447–449. [Google Scholar] [CrossRef] [PubMed]
- Yassin, H.M.; Dever, L.L. Telithromycin: A new ketolide antimicrobial for treatment of respiratory tract infections. Expert Opin. Investig. Drugs 2001, 10, 353–367. [Google Scholar] [CrossRef]
- Jamal, W.Y.; Al Hashem, G.; Khodakhast, F.; Rotimi, V.O. Comparative in vitro activity of tigecycline and nine other antibiotics against gram-negative bacterial isolates, including ESBL-producing strains. J. Chemother. 2009, 21, 261–266. [Google Scholar] [CrossRef]
- Zhanel, G.G.; DeCorby, M.; Adam, H.; Mulvey, M.R.; McCracken, M.; Lagacé-Wiens, P.; Nichol, K.A.; Wierzbowski, A.; Baudry, P.J.; Tailor, F.; et al. Prevalence of antimicrobial-resistant pathogens in Canadian hospitals: Results of the Canadian Ward Surveillance Study (CANWARD 2008). Antimicrob. Agents Chemother. 2010, 54, 4684–4693. [Google Scholar] [CrossRef]
- Bodmann, K.-F. Kalkulierte parenterale Initialtherapie bakterieller Erkrankungen bei Erwachsenen—Update 2018: PEG S2k Leitlinie (AWMF-Registernummer 082-006); 2. aktualisierte Version, erstellt am 25. Juli 2019; Paul-Ehrlich-Gesellschaft für Chemotherapie e.V: Rheinbach, Germany, 2019; ISBN 978-3-9820853-0-2. [Google Scholar]
- Johnson, R.C. Isolation techniques for spirochetes and their sensitivity to antibiotics in vitro and in vivo. Rev. Infect. Dis. 1989, 11 (Suppl. S6), S1505–S1510. [Google Scholar] [CrossRef]
- Lakos, A.; Nagy, G. Effect of an antibiotic combination on the propagation of Borrelia burgdorferi causative agent Lyme disease. Orv. Hetil. 1999, 140, 1529–1532. [Google Scholar]
- Murgia, R.; Marchetti, F.; Cinco, M. Comparative bacteriostatic and bactericidal activities of cefodizime against Borrelia burgdorferi sensu lato. Antimicrob. Agents Chemother. 1999, 43, 3030–3032. [Google Scholar] [CrossRef]
- Galbraith, K.M.; Ng, A.C.; Eggers, B.J.; Kuchel, C.R.; Eggers, C.H.; Samuels, D.S. parC mutations in fluoroquinolone-resistant Borrelia burgdorferi. Antimicrob. Agents Chemother. 2005, 49, 4354–4357. [Google Scholar] [CrossRef]
- Samuels, D.S.; Garon, C.F. Coumermycin A1 inhibits growth and induces relaxation of supercoiled plasmids in Borrelia burgdorferi, the Lyme disease agent. Antimicrob. Agents Chemother. 1993, 37, 46–50. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Knight, S.W.; Samuels, D.S. Natural synthesis of a DNA-binding protein from the C-terminal domain of DNA gyrase A in Borrelia burgdorferi. EMBO J. 1999, 18, 4875–4881. [Google Scholar] [CrossRef] [PubMed]
- Dever, L.L.; Torigian, C.V.; Barbour, A.G. In vitro activities of the everninomicin SCH 27899 and other newer antimicrobial agents against Borrelia burgdorferi. Antimicrob. Agents Chemother. 1999, 43, 1773–1775. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Feng, J.; Zhang, S.; Shi, W.; Zhang, Y. Activity of sulfa drugs and their combinations against stationary phase B. burgdorferi in vitro. Antibiotics 2017, 6, 10. [Google Scholar] [CrossRef] [PubMed]
- Pothineni, V.R.; Potula, H.-H.S.K.; Ambati, A.; Mallajosyula, V.V.A.; Sridharan, B.; Inayathullah, M.; Ahmed, M.S.; Rajadas, J. Azlocillin can be the potential drug candidate against drug-tolerant Borrelia burgdorferi sensu stricto JLB31. Sci. Rep. 2020, 10, 3798. [Google Scholar] [CrossRef]
- van Dam, A.P.; Kuiper, H.; Vos, K.; Widjojokusumo, A.; de Jongh, B.M.; Spanjaard, L.; Ramselaar, A.C.; Kramer, M.D.; Dankert, J. Different genospecies of Borrelia burgdorferi are associated with distinct clinical manifestations of Lyme borreliosis. Clin. Infect. Dis. 1993, 17, 708–717. [Google Scholar] [CrossRef]
- Hodzic, E. Lyme Borreliosis: Is there a preexisting (natural) variation in antimicrobial susceptibility among Borrelia burgdorferi strains? Bosn. J. Basic Med. Sci. 2015, 15, 1–13. [Google Scholar] [CrossRef][Green Version]
- Peter, O.; Bretz, A.G. In vitro susceptibility of B. burgdorferi, B. garinii and B. afzelii to 7 antimicrobial agents. In Proceedings of the VIth International Conference Lyme borreliosis, Bologna, Italy, 19–22 June 1994; pp. 167–170. [Google Scholar]
- Fingerle, V.; Eiffert, H.; Gessner, A.; Göbel, B.; Hofmann, H.; Hunfeld, K.P.; Krause, A.; Pfister, H.W.; Reischl, U.; Sing, A.; et al. Lyme-Borreliose. 2. Auflage; Elsevier Urban & Fischer: München, Germany, 2017; ISBN 9783437226052. [Google Scholar]
- Ruzić-Sabljić, E.; Podreka, T.; Maraspin, V.; Strle, F. Susceptibility of Borrelia afzelii strains to antimicrobial agents. Int. J. Antimicrob. Agents 2005, 25, 474–478. [Google Scholar] [CrossRef]
- Boerner, J. Empfindlichkeitsprüfung von Borrelia burgdorferi gegen Antibiotika und Chemotherapeutika. Ph.D. Thesis, Justus-Liebig Universität, Gießen, Germany, 1994. [Google Scholar]
- Reisinger, E.C.; Wendelin, I.; Gasser, R. Inactivation of diaminopyrimidines and sulfonamides in Barbour-Stoenner-Kelly medium for isolation of Borrelia burgdorferi. Eur. J. Clin. Microbiol. Infect. Dis. 1995, 14, 732–733. [Google Scholar] [CrossRef]
- Karlsson, M.; Hammers, S.; Nilsson-Ehle, I.; Malmborg, A.S.; Wretlind, B. Concentrations of doxycycline and penicillin G in sera and cerebrospinal fluid of patients treated for neuroborreliosis. Antimicrob. Agents Chemother. 1996, 40, 1104–1107. [Google Scholar] [CrossRef]
- Reisinger, E.; Wendelin, I.; Gasser, R.; Halwachs, G.; Wilders-Truschnig, M.; Krejs, G. Antibiotics and increased temperature against Borrelia burgdorferi in vitro. Scand. J. Infect. Dis. 1996, 28, 155–157. [Google Scholar] [CrossRef] [PubMed]
- Reisinger, E.C.; Wendelin, I.; Gasser, R. In vitro activity of trimethoprim against Borrelia burgdorferi. Eur. J. Clin. Microbiol. Infect. Dis. 1997, 16, 458–460. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrik, F.B. Sexually transmitted diseases. In Dermatology in General Medicine: Textbook and Atlas; Fitzpatrick, T.B., Ed.; McGraw-Hill: New York, NY, USA, 1987; ISBN 0070796890. [Google Scholar]
- Fujita, K.; Baba, T.; Isono, K. Genomic Analysis of the Genes Encoding Ribosomal Proteins in Eight Eubacterial Species and Saccharomyces cerevisiae. Genome Inform. Ser. Workshop Genome Inform. 1998, 9, 3–12. [Google Scholar]
- Terekhova, D.; Sartakova, M.L.; Wormser, G.P.; Schwartz, I.; Cabello, F.C. Erythromycin resistance in Borrelia burgdorferi. Antimicrob. Agents Chemother. 2002, 46, 3637–3640. [Google Scholar] [CrossRef] [PubMed]
- Bunikis, I.; Denker, K.; Ostberg, Y.; Andersen, C.; Benz, R.; Bergström, S. An RND-type efflux system in Borrelia burgdorferi is involved in virulence and resistance to antimicrobial compounds. PLoS Pathog. 2008, 4, e1000009. [Google Scholar] [CrossRef]
- Samuels, D.S.; Mach, K.E.; Garon, C.F. Genetic transformation of the Lyme disease agent Borrelia burgdorferi with coumarin-resistant gyrB. J. Bacteriol. 1994, 176, 6045–6049. [Google Scholar] [CrossRef]
- Samuels, D.S.; Marconi, R.T.; Huang, W.M.; Garon, C.F. gyrB mutations in coumermycin A1-resistant Borrelia burgdorferi. J. Bacteriol. 1994, 176, 3072–3075. [Google Scholar] [CrossRef][Green Version]
- Morrissey, I.; George, J.T. Purification of pneumococcal type II topoisomerases and inhibition by gemifloxacin and other quinolones. J. Antimicrob. Chemother. 2000, 45 (Suppl. S1), S101–S106. [Google Scholar] [CrossRef][Green Version]
- Lewin, C.S.; Morrissey, I.; Smith, J.T. The bactericidal activity of sparfloxacin. J. Antimicrob. Chemother. 1992, 30, 625–632. [Google Scholar] [CrossRef]
- Cabello, F.C.; Embers, M.E.; Newman, S.A.; Godfrey, H.P. Borreliella burgdorferi Antimicrobial-Tolerant Persistence in Lyme Disease and Posttreatment Lyme Disease Syndromes. mBio 2022, 13, e0344021. [Google Scholar] [CrossRef]
- Caskey, J.R.; Hasenkampf, N.R.; Martin, D.S.; Chouljenko, V.N.; Subramanian, R.; Cheslock, M.A.; Embers, M.E. The functional and molecular effects of doxycycline treatment on Borrelia burgdorferi phenotype. Front. Microbiol. 2019, 10, 690. [Google Scholar] [CrossRef] [PubMed]
- Greer, N.D. Tigecycline (Tygacil): The first in the glycylcycline class of antibiotics. In Baylor University Medical Center Proceedings; Taylor & Francis: Abingdon-on-Thames, UK, 2006; Volume 19, pp. 155–161. [Google Scholar] [CrossRef]
- Zhanel, G.G.; Homenuik, K.; Nichol, K.; Noreddin, A.; Vercaigne, L.; Embil, J.; Gin, A.; Karlowsky, J.A.; Hoban, D.J. The glycylcyclines: A comparative review with the tetracyclines. Drugs 2004, 64, 63–88. [Google Scholar] [CrossRef] [PubMed]
- Chau, S.-L.; Chu, Y.-W.; Houang, E.T.S. Novel resistance-nodulation-cell division efflux system AdeDE in Acinetobacter genomic DNA group 3. Antimicrob. Agents Chemother. 2004, 48, 4054–4055. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Weitner, M.; Shi, W.; Zhang, S.; Sullivan, D.; Zhang, Y. Identification of additional anti-persister activity against Borrelia burgdorferi from an FDA drug library. Antibiotics 2015, 4, 397–410. [Google Scholar] [CrossRef]
- Feng, J.; Shi, W.; Zhang, S.; Sullivan, D.; Auwaerter, P.G.; Zhang, Y. A drug combination screen identifies drugs active against amoxicillin-induced round bodies of in vitro Borrelia burgdorferi persisters from an FDA drug library. Front. Microbiol. 2016, 7, 743. [Google Scholar] [CrossRef]
- Feng, J.; Auwaerter, P.G.; Zhang, Y. Drug combinations against Borrelia burgdorferi persisters in vitro: Eradication achieved by using daptomycin, cefoperazone, and doxycycline. PLoS ONE 2015, 10, e0117207. [Google Scholar] [CrossRef]
- Feng, J.; Wang, T.; Shi, W.; Zhang, S.; Sullivan, D.; Auwaerter, P.G.; Zhang, Y. Identification of novel activity against Borrelia burgdorferi persisters using an FDA approved drug library. Emerg. Microbes Infect. 2014, 3, e49. [Google Scholar] [CrossRef]
- Hunfeld, K.-P.; Burg, S.; Hanssen-Hübner, C.; Karas, M.; Brade, V.; Kraiczy, P. Changes in the expression pattern of structural proteins after exposure of Borrelia burgdorferi to penicillin G and doxycycline. Int. J. Med. Microbiol. 2008, 298, 325–332. [Google Scholar] [CrossRef]
- Jungblut, P.R.; Grabher, G.; Stöffler, G. Comprehensive detection of immune relevant Borrelia garinii antigens by two-dimensional electrophoresis. Electrophoresis 1999, 20, 3611–3622. [Google Scholar] [CrossRef]
- Klose, J.; Kobalz, U. Two-dimensional electrophoresis of proteins: An updated protocol and implications for a functional analysis of the genome. Electrophoresis 1995, 16, 1034–1059. [Google Scholar] [CrossRef]
- Barbour, A.G.; Hayes, S.F. Biology of Borrelia species. Microbiol. Rev. 1986, 50, 381–400. [Google Scholar] [CrossRef]
- Brorson, O.; Brorson, S.H. A rapid method for generating cystic forms of Borrelia burgdorferi, and their reversal to mobile spirochetes. APMIS 1998, 106, 1131–1141. [Google Scholar] [CrossRef] [PubMed]
- Brorson, O.; Brorson, S.H. Transformation of cystic forms of Borrelia burgdorferi to normal, mobile spirochetes. Infection 1997, 25, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Alban, P.S.; Johnson, P.W.; Nelson, D.R. Serum-starvation-induced changes in protein synthesis and morphology of Borrelia burgdorferi. Microbiology 2000, 146 Pt 1, 119–127. [Google Scholar] [CrossRef]
- Nitzan, Y.; Deutsch, E.B.; Pechatnikov, I. Diffusion of beta-lactam antibiotics through oligomeric or monomeric porin channels of some gram-negative bacteria. Curr. Microbiol. 2002, 45, 446–455. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Martínez, L.; Hernández-Allés, S.; Albertí, S.; Tomás, J.M.; Benedi, V.J.; Jacoby, G.A. In vivo selection of porin-deficient mutants of Klebsiella pneumoniae with increased resistance to cefoxitin and expanded-spectrum-cephalosporins. Antimicrob. Agents Chemother. 1996, 40, 342–348. [Google Scholar] [CrossRef] [PubMed]
- El Fakhry, Y.; Ouellette, M.; Papadopoulou, B. A proteomic approach to identify developmentally regulated proteins in Leishmania infantum. Proteomics 2002, 2, 1007. [Google Scholar] [CrossRef]
- Seyfzadeh, M.; Keener, J.; Nomura, M. spoT-dependent accumulation of guanosine tetraphosphate in response to fatty acid starvation in Escherichia coli. Proc. Natl. Acad. Sci. USA 1993, 90, 11004–11008. [Google Scholar] [CrossRef]
- Georgilis, K.; Peacocke, M.; Klempner, M.S. Fibroblasts protect the Lyme disease spirochete, Borrelia burgdorferi, from ceftriaxone in vitro. J. Infect. Dis. 1992, 166, 440–444. [Google Scholar] [CrossRef]
- Brouqui, P.; Badiaga, S.; Raoult, D. Eucaryotic cells protect Borrelia burgdorferi from the action of penicillin and ceftriaxone but not from the action of doxycycline and erythromycin. Antimicrob. Agents Chemother. 1996, 40, 1552–1554. [Google Scholar] [CrossRef]
- Sapi, E.; Kaur, N.; Anyanwu, S.; Luecke, D.F.; Datar, A.; Patel, S.; Rossi, M.; Stricker, R.B. Evaluation of in-vitro antibiotic susceptibility of different morphological forms of Borrelia burgdorferi. Infect. Drug Resist. 2011, 4, 97–113. [Google Scholar] [CrossRef] [PubMed]
- Sapi, E.; Theophilus, P.A.S.; Pham, T.V.; Burugu, D.; Luecke, D.F. Effect of RpoN, RpoS and LuxS Pathways on the Biofilm Formation and Antibiotic Sensitivity of Borrelia burgdorferi. Eur. J. Microbiol. Immunol. 2016, 6, 272–286. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.P.; Schoen, R.T.; Rahn, D.W.; Sikand, V.K.; Nowakowski, J.; Parenti, D.L.; Holman, M.S.; Persing, D.H.; Steere, A.C. Clinical characteristics and treatment outcome of early Lyme disease in patients with microbiologically confirmed erythema migrans. Ann. Intern. Med. 2002, 136, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Thanassi, W.T.; Schoen, R.T. The Lyme disease vaccine: Conception, development, and implementation. Ann. Intern. Med. 2000, 132, 661–668. [Google Scholar] [CrossRef]
- Panconesi, E.; Zuccati, G.; Cantini, A. Treatment of syphilis: A short critical review. Sex. Transm. Dis. 1981, 8, 321–325. [Google Scholar]
- Viljanen, M.K.; Oksi, J.; Salomaa, P.; Skurnik, M.; Peltonen, R.; Kalimo, H. Cultivation of Borrelia burgdorferi from the blood and a subcutaneous lesion of a patient with relapsing febrile nodular nonsuppurative panniculitis. J. Infect. Dis. 1992, 165, 596–597. [Google Scholar] [CrossRef]
- Wormser, G.P.; Ramanathan, R.; Nowakowski, J.; McKenna, D.; Holmgren, D.; Visintainer, P.; Dornbush, R.; Singh, B.; Nadelman, R.B. Duration of antibiotic therapy for early Lyme disease. A randomized, double-blind, placebo-controlled trial. Ann. Intern. Med. 2003, 138, 697–704. [Google Scholar] [CrossRef]
- Straubinger, R.K.; Summers, B.A.; Chang, Y.F.; Appel, M.J. Persistence of Borrelia burgdorferi in experimentally infected dogs after antibiotic treatment. J. Clin. Microbiol. 1997, 35, 111–116. [Google Scholar] [CrossRef]
- Oksi, J.; Marjamäki, M.; Nikoskelainen, J.; Viljanen, M.K. Borrelia burgdorferi detected by culture and PCR in clinical relapse of disseminated Lyme borreliosis. Ann. Med. 1999, 31, 225–232. [Google Scholar] [CrossRef]
- Lomholt, H.; Lebech, A.M.; Hansen, K.; Brandrup, F.; Halkier-Sørensen, L. Long-term serological follow-up of patients treated for chronic cutaneous borreliosis or culture-positive erythema migrans. Acta Derm. Venereol. 2000, 80, 362–366. [Google Scholar] [CrossRef]
- Logar, M.; Lotric-Furlan, S.; Maraspin, V.; Cimperman, J.; Jurca, T.; Ruzić-Sabljić, E.; Strle, F. Has the presence or absence of Borrelia burgdorferi sensu lato as detected by skin culture any influence on the course of erythema migrans? Wien. Klin. Wochenschr. 1999, 111, 945–950. [Google Scholar] [PubMed]
- Bockenstedt, L.K.; Mao, J.; Hodzic, E.; Barthold, S.W.; Fish, D. Detection of attenuated, noninfectious spirochetes in Borrelia burgdorferi-infected mice after antibiotic treatment. J. Infect. Dis. 2002, 186, 1430–1437. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, E.L.; Gastanaduy, A.S.; Huwe, B.B. The role of the carrier in treatment failures after antibiotic for group A streptococci in the upper respiratory tract. J. Lab. Clin. Med. 1981, 98, 326–335. [Google Scholar] [PubMed]
- van den Brule, A.J.C.; Munk, C.; Winther, J.F.; Kjaer, S.K.; Jørgensen, H.O.; Meijer, C.J.L.M.; Morré, S.A. Prevalence and persistence of asymptomatic Chlamydia trachomatis infections in urine specimens from Danish male military recruits. Int. J. STD AIDS 2002, 13 (Suppl. S2), S19–S22. [Google Scholar] [CrossRef] [PubMed]
- Embers, M.E.; Hasenkampf, N.R.; Jacobs, M.B.; Tardo, A.C.; Doyle-Meyers, L.A.; Philipp, M.T.; Hodzic, E. Variable manifestations, diverse seroreactivity and post-treatment persistence in non-human primates exposed to Borrelia burgdorferi by tick feeding. PLoS ONE 2017, 12, e0189071. [Google Scholar] [CrossRef] [PubMed]
- Crossland, N.A.; Alvarez, X.; Embers, M.E. Late disseminated lyme disease: Associated pathology and spirochete persistence posttreatment in Rhesus Macaques. Am. J. Pathol. 2018, 188, 672–682. [Google Scholar] [CrossRef]
- Criswell, D.; Tobiason, V.L.; Lodmell, J.S.; Samuels, D.S. Mutations conferring aminoglycoside and spectinomycin resistance in Borrelia burgdorferi. Antimicrob. Agents Chemother. 2006, 50, 445–452. [Google Scholar] [CrossRef]
- Lukehart, S.A.; Godornes, C.; Molini, B.J.; Sonnett, P.; Hopkins, S.; Mulcahy, F.; Engelman, J.; Mitchell, S.J.; Rompalo, A.M.; Marra, C.M.; et al. Macrolide resistance in Treponema pallidum in the United States and Ireland. N. Engl. J. Med. 2004, 351, 154–158. [Google Scholar] [CrossRef]
- Breier, F.; Khanakah, G.; Stanek, G.; Kunz, G.; Aberer, E.; Schmidt, B.; Tappeiner, G. Isolation and polymerase chain reaction typing of Borrelia afzelii from a skin lesion in a seronegative patient with generalized ulcerating bullous lichen sclerosus et atrophicus. Br. J. Dermatol. 2001, 144, 387–392. [Google Scholar] [CrossRef]
- Aberer, E.; Kersten, A.; Klade, H.; Poitschek, C.; Jurecka, W. Heterogeneity of Borrelia burgdorferi in the skin. Am. J. Dermatopathol. 1996, 18, 571–579. [Google Scholar] [CrossRef]
- Pachner, A.R.; Basta, J.; Delaney, E.; Hulinska, D. Localization of Borrelia burgdorferi in murine Lyme borreliosis by electron microscopy. Am. J. Trop. Med. Hyg. 1995, 52, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Comstock, L.E.; Thomas, D.D. Penetration of endothelial cell monolayers by Borrelia burgdorferi. Infect. Immun. 1989, 57, 1626–1628. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Sturrock, A.; Weis, J.J. Intracellular localization of Borrelia burgdorferi within human endothelial cells. Infect. Immun. 1991, 59, 671–678. [Google Scholar] [CrossRef] [PubMed]
- Strle, F.; Nelson, J.A.; Ruzic-Sabljic, E.; Cimperman, J.; Maraspin, V.; Lotric-Furlan, S.; Cheng, Y.; Picken, M.M.; Trenholme, G.M.; Picken, R.N. European Lyme borreliosis: 231 culture-confirmed cases involving patients with erythema migrans. Clin. Infect. Dis. 1996, 23, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, I.; Wormser, G.P.; Schwartz, J.J.; Cooper, D.; Weissensee, P.; Gazumyan, A.; Zimmermann, E.; Goldberg, N.S.; Bittker, S.; Campbell, G.L.; et al. Diagnosis of early Lyme disease by polymerase chain reaction amplification and culture of skin biopsies from erythema migrans lesions. J. Clin. Microbiol. 1992, 30, 3082–3088. [Google Scholar] [CrossRef]
- Zore, A.; Ruzić-Sabljić, E.; Maraspin, V.; Cimperman, J.; Lotric-Furlan, S.; Pikelj, A.; Jurca, T.; Logar, M.; Strle, F. Sensitivity of culture and polymerase chain reaction for the etiologic diagnosis of erythema migrans. Wien. Klin. Wochenschr. 2002, 114, 606–609. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference | MIC a Definition | MBC b Definition | Inoculum | Exposure to Antimicrobial Agent | Method |
|---|---|---|---|---|---|
| Agger et al., 1992 [41] | The lowest concentration of antibiotic without motile borreliae (microscopic reading mode) | The lowest concentration of antibiotic without growth of borreliae after 7 days of subculture in a liquid medium (microscopic reading mode) | 105/mL | 7 d | Macrodilution |
| Alder et al., 1993 [66] | The lowest dilution of antibiotics showed less than 5% vital borreliae (microscopic reading mode) | Not performed | 5 × 106/mL | 48 h | Macrodilution |
| Berger et al., 1985 [67] | The lowest concentration of antibiotic showed no greater number of motile borreliae than the initial inoculum and more than 90% motility of cells (microscopic reading mode) | The lowest concentration of antibiotic without growth of borreliae 11 days of subculture in liquid medium (microscopic reading mode) | 105/mL | 72 h | Macrodilution |
| Dever et al., 1992 [40] | The lowest concentration of antibiotic without macroscopic growth (pellet) or indicator color shift (macroscopic reading mode) | The lowest concentration of antibiotic with a 99.9% reduction of initial inoculum in subculture after 10–12 days (subsurface plating) | 105/mL | 72 h | Microdilution |
| Dever et al., 1992 [40] | The lowest concentration of antibiotic without macroscopic growth (pellet) or indicator color shift (macroscopic reading mode) | The lowest concentration of antibiotic without growth of borreliae in a liquid medium after 10–12 days (macroscopic reading mode) | 105/mL | 72 h | Macrodilution |
| Hansen et al., 1992 [61] | Not performed | The lowest concentration of antibiotic with complete growth inhibition of borreliae in subculture after 7 days (microscopic reading mode) | 105/mL | 7 d | Macrodilution |
| Hunfeld et al., 2000 [51] | The lowest concentration of antibiotic without detectable indicator color shift (photometric reading mode) | The lowest concentration of antibiotic without re-growth of borreliae after 3 weeks of subculture in a liquid medium (microscopic reading mode) | 2.5 × 107/mL | 72 h | Microdilution |
| Koetsveld et.al., 2017 [48] | The lowest concentration of antibiotic without detectable indicator color shift (photometric reading mode) | The lowest concentration of antibiotic without re-growth of borreliae after 3 weeks of subculture in a liquid medium (microscopic reading mode) | 5 × 107/mL | 72 h | Microdilution |
| Johnson et al., 1984 [68] | The lowest concentration of antibiotic showed no greater number of motile borreliae than the initial inoculum (microscopic reading mode) and no sediment | Not performed | 105/mL | 5 d | Macrodilution |
| Luft et al., 1988 [64] | The lowest concentration of antibiotic shows no greater number of motile borreliae than in the initial inoculum (microscopic reading mode) | The lowest concentration of antibiotic without growth of borreliae after 3 weeks of subculture in a liquid medium (microscopic reading mode) | 2 × 105/mL | 72 h | Macrodilution |
| Leimer et al., 2021 [69] | The lowest concentration of antibiotics that inhibited growth | The lowest concentration of antibiotic with 99.9% reduction of initial inoculum in subculture after 20 days (semi-agar plates) | 106/mL | 7 d | Microdilution |
| Morsehd et al., 1993 [70] | The lowest concentration of antibiotic with complete growth inhibition (microscopic reading mode) | Not performed | 104/mL | 8–10 d | Macrodilution |
| Mursic et al., 1987 [38] | The lowest concentration of antibiotic without detectable re-culturable organisms (microscopic reading mode) | Not performed | 105/mL | 5 d | Macrodilution |
| Stiernstedt et al., 1999 [71] | The lowest dilution of antibiotic shows the same number of motile borreliae as the initial inoculum (microscopic reading mode) | The lowest concentration of antibiotic without re-growth of borreliae after 2 weeks of subculture in liquid medium (microscopic reading mode) | 106/mL | 7 d | Dialysis culture |
| Sharma B et al., 2015 [72] | The lowest concentration of antibiotics that inhibited growth | The lowest concentration of antibiotic with 99.9% reduction of initial inoculum in subculture after 20 days (semi-agar plates) | 106/mL | 72 h | Microdilution |
| Antimicrobials | Colorimetric MIC90 a (µg/mL) | MIC Range (µg/mL) (Adopted from the Literature) | MBC90 b (µg/mL) (100% Killing after 72 h) | MBC Range (µg/mL) (Adopted from the Literature) |
|---|---|---|---|---|
| ß-lactames | ||||
| Penicillin G | 1 | 0.03–8 | 16 | 0.05–>50 |
| Amoxycillin | 1 | 0.03–2 | 32 | <0.03–3.2 |
| Piperacillin | 0.06 | <0.06–0.125 | 2 | 1.3–2.6 |
| Mezlocillin | ≤0.06 | <0.06–1 | 2 | 0.125–2 |
| Azlocillin | 0.125 | 0.125 | 4 | N.A. |
| Aztreonam | >64 | 2–>64 | >64 | 64–>256 |
| Cefaclor | 16 | 16–128 | >64 | 64–128 |
| Loracarbef | 32 | 4–32 | >128 | 128–>128 |
| Ceftibuten | 32 | 4–32 | >128 | 128–>128 |
| Cefixime | 1 | 0.25–4 | 32 | 0.8–32 |
| Cefuroxime Cefpodoxim | 0.25 4 | 0.06–0.5 1–8 | 16 32 | 0.25–>16 32–>32 |
| Cefotaxime | 0.125 | 0.01–1 | 8 | 0.02–0.25 |
| Ceftriaxone | 0.03 | <0.01–0.125 | 2 | 0.02–3.81 |
| Sulbactam | 64 | 0.125–256 | >64 | >64 |
| Carbapenems | ||||
| Meropenem | 0.25 | 0.012–0.5 | 16 | 0.5–32 |
| Faropenem Imipenem Ertapenem | 4 0.5 0.125 | 0.03–8 0.06–0.5 0.015–0.125 | 64 32 4 | 8–>64 16–32 0.5–4 |
| Glycopeptides | ||||
| Vancomycin | 1 | 0.25–2 | 16 | 2–32 |
| Teicoplanin Lipopeptides | >8 | 2–>8 | >8 | >8 |
| Daptomycin | N.A. | 0.25–32 | >32 | >32 |
| Tetracyclines | ||||
| Tetracycline | 0.25 | 0.01–20 | N.A. | 0.8–4.1 |
| Minocycline | 0.25 | 0.03–1 | 5.8 (mean MBC) | 3–8 |
| Doxycycline | 0.25 | 0.06–2 | N.A. | 0.25–6.4 |
| Tigecycline | >0.016 | <0.016 | 0.25 | 0.25–1 |
| Macrolides | ||||
| Erythromycin | 0.062 | ≤0.007–1 | >0.5 | 0.05–2.17 |
| Roxithromycin | 0.062 | 0.015–0.12 | >0.5 | 0.0125–1.8 |
| Azithromycin | 0.015 | 0.003–0.03 | 0.5 | 0.007–0.5 |
| Streptogramins | ||||
| Quinopristin/ Dalfopristin | 0.125 | <0.015–0.25 | 8 | 2–8 |
| Ketolides | ||||
| Telithromycin | 0.0078 | <0.0002–0.0078 | 0.25 | 0.0156–0.5 |
| Cethromycin | 0.0019 | <0.0002–0.0078 | 0.125 | 0.0156–0.25 |
| ABT-773 | >0.0020 | >0.0020 | 0.35 | 0.02–1 |
| Cinnamic acids | ||||
| Hygromycin A | N.A. | 0.25 | 1 | N.A. |
| Aminoglycosides | ||||
| Tobramycin | 64 | 8–64 | >64 | N.A. |
| Amikacin | 128 | 32–>128 | >128 | >128 |
| Ribostamycin | >32 | 8–64 | >32 | 4–>32 |
| Spectinomycin | 2 | 0.25–2 | N.A | N.A. |
| Quinolones | ||||
| Nalidixic acid | 256 | 128–>512 | >512 | ≥512 |
| Norfloxacin | 8 | 1–16 | >64 | 8–>64 |
| Ofloxacin | 8 | 1–16 | >16 | 8–>16 |
| Ciprofloxacin | 2 | 0.25–8 | >16 | 4–>16 |
| Sparfloxacin Grepafloxacin Gatifloxacin Sitafloxacin | 1 0.5 1 0.5 | 0.06–8 0.6–2 0.25–2 0.6–4 | >16 16 >16 >16 | 2–>16 2–>16 4–>16 1–>16 |
| Gemifloxacin | 0.12 | 0.03–0.25 | 8 | 0.25–16 |
| Others | ||||
| Fusidic acid | >4 | >4 | >4 | >4 |
| Chloramphenicol Linezolid Sulfamethoxazol | N.A. N.A. N.A. | 1.25–2 >2 8 | N.A. >2 >256 | N.A. N.A. 32–>256. |
| Spot | pI | Mol. Mass (kDa) | Up- or Down | Mol. Mass (kDa) | Identified Protein |
|---|---|---|---|---|---|
| (Est. from Gel) | (Est. from Gel) | Regulation | (From Database) | (By MALDI-TOF MS) | |
| P-2702 | 5.9 | 65 | d | 86.13 | Membrane-associated protein |
| P66 (BB0603) | |||||
| P-3703 | 6.2 | 65 | d | 68.13 | Membrane-associated protein |
| p66 (BB0603) | |||||
| P-4702 | 6.4 | 63 | d | 63.496 | Ribosomal protein S1 (RpsA) |
| (BB0127) | |||||
| P-4706 | 6.7 | 63 | d | NA | No match found |
| P-4802 | 6.4 | 75 | d | NA | No match found |
| P-4803 | 6.5 | 75 | d | 80.298 | Polyribonucleotide |
| nucleotidyltransferase (PnpA) | |||||
| (BB0805) | |||||
| P-4804 | 6.6 | 75 | d | NA | No match found |
| P-5702 | 7 | 57 | d | 50.061 | Pyrophosphate-fructose-6- |
| phosphate-1-phosphortransferase (Pfp) | |||||
| (BB0727) | |||||
| P-7103 | 8.1 | 25 | d | 30.417 | Predicted coding region |
| (BB0238) | |||||
| P-7602 | 7.8 | 50 | d | 55.856 | Glycerol kinase (GlpK) |
| (BB0241) | |||||
| P-7603 | 7.9 | 49 | d | 53.227 | Pyruvate kinase (Pyk) |
| (BB0348) | |||||
| D/P-4703 | 6.5 | 65 | d | NA | No match found |
| D/P-4705 | 6.6 | 63 | d | NA | No match found |
| D/P-7001 | 7.9 | 23 | d | 26.569 | Pfs protein (Pfs-1) |
| (BB0375) | |||||
| D-0504 | 5.2 | 42 | d | 42.372 | Cell division protein (FtsZ) |
| (BB0229) | |||||
| D-4101 | 6.4 | 26 | u | 27.984 | Triosephosphate isomerase |
| EM Patients | |||||
|---|---|---|---|---|---|
| Clinical information | 1 | 2 | 3 | 4 | 5 |
| Sex | F | F | F | F | M |
| Age (yrs.) | 43 | 38 | 53 | 68 | 36 |
| Symptoms at first visit a | MEM | EM | EM | EM, L, PFP | EM, L |
| Recognition of tick bite | Yes | Yes | No | No | Yes |
| Size of EM (cm) | 7 × 2; 6 × 8 | 10 × 16 | 7 × 11 | 9 × 12 | 7 × 12 |
| Systemic complaints b | None | None | Yes | None | None |
| Initial treatment Duration (d) | Ceftriaxone 1 × 2 g i.v. 14 | Amoxicillin 3 × 500 mg p.o. 14 | Cefuroxime 2 × 500 mg p.o. 14 | Cefuroxime 2 × 500 mg p.o. 14 | Azithromycin 2 × 500 mg p.o. 1 1 × 500 mg p.o. 4 |
| IgM-IFT (titre) c | 256 | 128 | Neg. | Neg. | Neg. |
| IgG-IFT (titre) c | 128 | 128 | Neg. | Neg. | Neg. |
| Initial culture | B. afzelii | B. garinii | B. afzelii | B. afzelii | B. afzelii |
| MIC/MBC d | 0.0156/0.25 | 0.0312/0.5 | 0.0625/8 | 0.0625/8 | 0.0039/0.125 |
| Follow-up culture | B. afzelii | B. garinii | B. afzelii | B. afzelii | B. garinii |
| MIC/MBC d | 0.0312/0.5 | 0.0625/1 | 0.125/16 | 0.0625/8 | 0.0019/0.125 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hunfeld, K.-P.; Kraiczy, P.; Norris, D.E.; Lohr, B. The In Vitro Antimicrobial Susceptibility of Borrelia burgdorferi sensu lato: Shedding Light on the Known Unknowns. Pathogens 2023, 12, 1204. https://doi.org/10.3390/pathogens12101204
Hunfeld K-P, Kraiczy P, Norris DE, Lohr B. The In Vitro Antimicrobial Susceptibility of Borrelia burgdorferi sensu lato: Shedding Light on the Known Unknowns. Pathogens. 2023; 12(10):1204. https://doi.org/10.3390/pathogens12101204
Chicago/Turabian StyleHunfeld, Klaus-Peter, Peter Kraiczy, Douglas E. Norris, and Benedikt Lohr. 2023. "The In Vitro Antimicrobial Susceptibility of Borrelia burgdorferi sensu lato: Shedding Light on the Known Unknowns" Pathogens 12, no. 10: 1204. https://doi.org/10.3390/pathogens12101204
APA StyleHunfeld, K.-P., Kraiczy, P., Norris, D. E., & Lohr, B. (2023). The In Vitro Antimicrobial Susceptibility of Borrelia burgdorferi sensu lato: Shedding Light on the Known Unknowns. Pathogens, 12(10), 1204. https://doi.org/10.3390/pathogens12101204