
Antibiofilm Action of Plant Terpenes in Salmonella Strains: Potential Inhibitors of the Synthesis of Extracellular Polymeric Substances

 ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Salmonella Is a Risk to Public Health
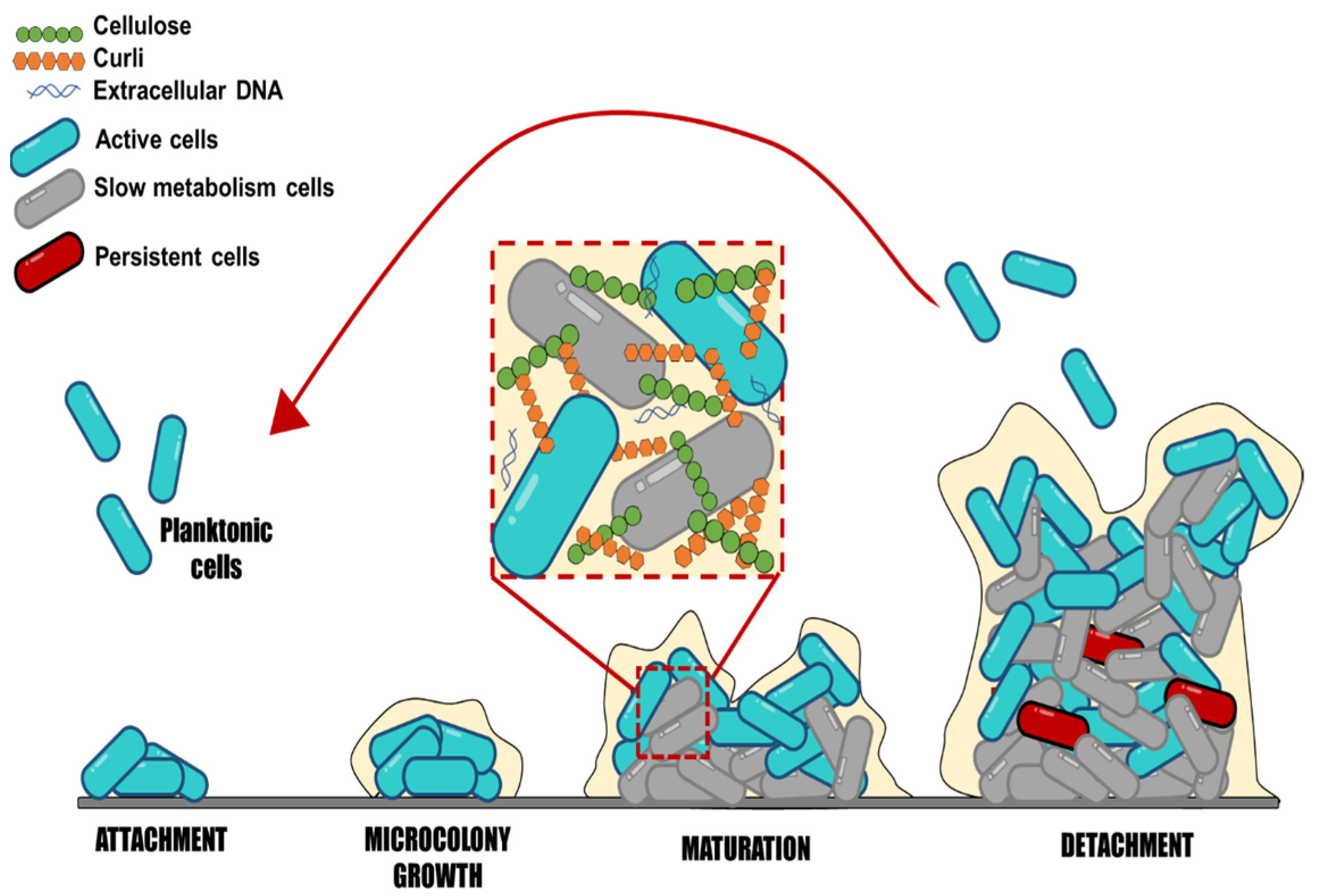
3. Salmonella Biofilm Formation Process
4. Main Structural EPS Produced during Salmonella Biofilm Formation
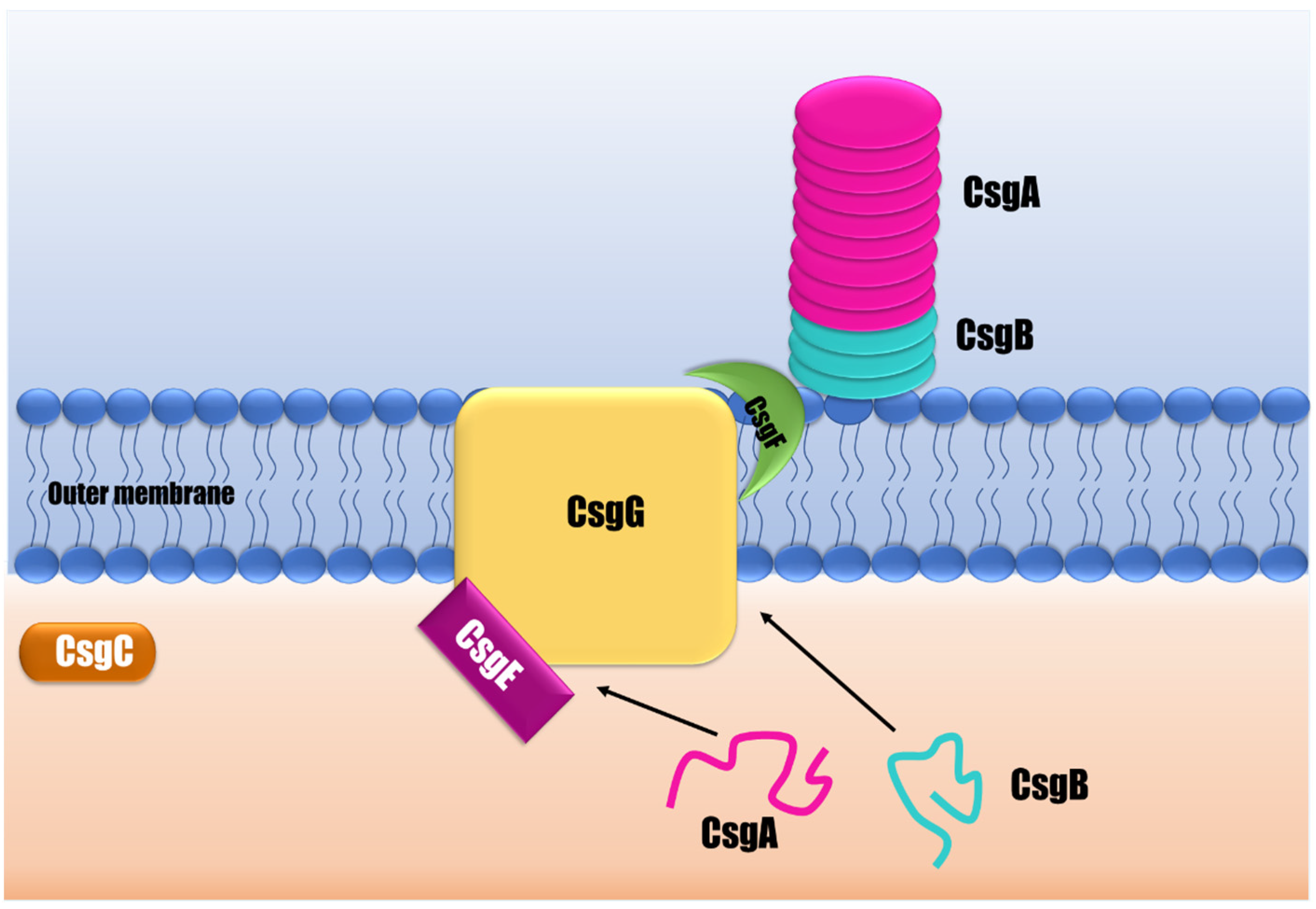
4.1. Curli
4.2. Cellulose
4.3. Colanic Acid
5. Inhibition of Extracellular Polymeric Substances to Reduce Biofilm Formation
6. Terpenes as Potential EPS Inhibitors in Salmonella Biofilms
7. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sell, J.; Dolan, B. Common gastrointestinal infections. Prim. Care Clin. Off. Pract. 2018, 45, 519–532. [Google Scholar] [CrossRef] [PubMed]
- Gordon, M.A. Salmonella infections in immunocompromised adults. J. Infect. 2008, 56, 413–422. [Google Scholar] [CrossRef]
- CDC. Salmonella Homepage. 2016. Available online: https://www.cdc.gov/salmonella/index.html (accessed on 15 January 2021).
- Merino, L.; Procura, F.; Trejo, F.M.; Bueno, D.J.; Golowczyc, M.A. Biofilm formation by Salmonella sp. in the poultry industry: Detection, control and eradication strategies. Food Res. Int. 2019, 119, 530–540. [Google Scholar] [CrossRef] [PubMed]
- Galiè, S.; García-Gutiérrez, C.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Biofilms in the Food Industry: Health Aspects and Control Methods. Front. Microbiol. 2018, 9, 898. [Google Scholar] [CrossRef] [PubMed]
- Van Houdt, R.; Michiels, C.W. Biofilm formation and the food industry, a focus on the bacterial outer surface. J. Appl. Microbiol. 2010, 109, 1117–1131. [Google Scholar] [CrossRef]
- Moraes, J.O.; Cruz, E.A.; Pinheiro, Í.; Oliveira, T.C.; Alvarenga, V.; Sant’Ana, A.S.; Magnani, M. An ordinal logistic regression approach to predict the variability on biofilm formation stages by five Salmonella enterica strains on polypropylene and glass surfaces as affected by pH, temperature and NaCl. Food Microbiol. 2019, 83, 95–103. [Google Scholar] [CrossRef]
- Sadekuzzaman, M.; Yang, S.; Mizan, M.; Ha, S. Current and Recent Advanced Strategies for Combating Biofilms. Compr. Rev. Food Sci. Food Saf. 2015, 14, 491–509. [Google Scholar] [CrossRef]
- Maruzani, R.; Sutton, G.; Nocerino, P.; Marvasi, M. Exopolymeric substances (EPS) from Salmonella enterica: Polymers, proteins and their interactions with plants and abiotic surfaces. J. Microbiol. 2018, 57, 1–8. [Google Scholar] [CrossRef]
- Flemming, H.-C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An emergent form of bacterial life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef]
- Powell, L.; Pritchard, M.F.; Ferguson, E.; Powell, K.A.; Patel, S.U.; Rye, P.; Sakellakou, S.-M.; Buurma, N.J.; Brilliant, C.; Copping, J.M.; et al. Targeted disruption of the extracellular polymeric network of Pseudomonas aeruginosa biofilms by alginate oligosaccharides. Npj Biofilms Microbiomes 2018, 4, 1–10. [Google Scholar] [CrossRef]
- Marshall, J.; Flechtner, A.D.; La Perle, K.M.; Gunn, J.S. Visualization of Extracellular Matrix Components within Sectioned Salmonella Biofilms on the Surface of Human Gallstones. PLoS ONE 2014, 9, e89243. [Google Scholar] [CrossRef] [PubMed]
- Friedman, M. Antibiotic-Resistant Bacteria: Prevalence in Food and Inactivation by Food-Compatible Compounds and Plant Extracts. J. Agric. Food Chem. 2015, 63, 3805–3822. [Google Scholar] [CrossRef] [PubMed]
- Lahiri, D.; Dash, S.; Dutta, R.; Nag, M. Elucidating the effect of anti-biofilm activity of bioactive compounds extracted from plants. J. Biosci. 2019, 44, 52. [Google Scholar] [CrossRef] [PubMed]
- Silva-Espinoza, B.A.; Palomares-Navarro, J.J.; Tapia-Rodriguez, M.R.; Cruz-Valenzuela, M.R.; González-Aguilar, G.A.; Silva-Campa, E.; Pedroza-Montero, M.; Almeida-Lopes, M.; Miranda, R.; Ayala-Zavala, J.F. Combination of ultraviolet light-C and clove essential oil to inactivate Salmonella typhimurium biofilms on stainless steel. J. Food Saf. 2020, 40, e12788. [Google Scholar] [CrossRef]
- Trevisan, D.A.C.; Da Silva, A.F.; Negri, M.; Filho, B.A.D.A.; Junior, M.M.; Patussi, E.V.; Campanerut-Sá, P.A.Z.; Mikcha, J.M.G. Antibacterial and antibiofilm activity of carvacrol against Salmonella enterica serotype Typhimurium. Braz. J. Pharm. Sci. 2018, 54. [Google Scholar] [CrossRef]
- Lira, M.C.; Rodrigues, J.B.; Almeida, E.T.D.C.; Ritter, A.C.; Tondo, E.; Torres, S.M.; Schaffner, D.; de Souza, E.L.; Magnani, M. Efficacy of oregano and rosemary essential oils to affect morphology and membrane functions of noncultivable sessile cells of Salmonella Enteritidis 86 in biofilms formed on stainless steel. J. Appl. Microbiol. 2019, 128, 376–386. [Google Scholar] [CrossRef]
- Miladi, H.; Zmantar, T.; Kouidhi, B.; Chaabouni, Y.; Mahdouani, K.; Bakhrouf, A.; Chaieb, K. Use of carvacrol, thymol, and eugenol for biofilm eradication and resistance modifying susceptibility of Salmonella enterica serovar Typhimurium strains to nalidixic acid. Microb. Pathog. 2017, 104, 56–63. [Google Scholar] [CrossRef]
- Abarkapa, I.; Čolović, R.; Đuragić, O.; Popović, S.; Kokić, B.; Milanov, D.; Pezo, L. Anti-biofilm activities of essential oils rich in carvacrol and thymol against Salmonella Enteritidis. Biofouling 2019, 35, 361–375. [Google Scholar] [CrossRef]
- Liu, F.; Jin, P.; Sun, Z.; Du, L.; Wang, D.; Zhao, T.; Doyle, M.P. Carvacrol oil inhibits biofilm formation and exopolysaccharide production of Enterobacter cloacae. Food Control. 2020, 119, 107473. [Google Scholar] [CrossRef]
- Kim, Y.-G.; Lee, J.-H.; Gwon, G.; Kim, S.-I.; Park, J.G.; Lee, J. Essential Oils and Eugenols Inhibit Biofilm Formation and the Virulence of Escherichia coli O157:H7. Sci. Rep. 2016, 6, 36377. [Google Scholar] [CrossRef]
- Alni, R.H.; Ghorban, K.; Dadmanesh, M. Combined effects of Allium sativum and Cuminum cyminum essential oils on planktonic and biofilm forms of Salmonella typhimurium isolates. 3 Biotech 2020, 10, 315. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Zhang, C.; Li, C.; Lin, L. Inhibition mechanism of cardamom essential oil on methicillin-resistant Staphylococcus aureus biofilm. LWT 2020, 122, 109057. [Google Scholar] [CrossRef]
- Zhang, C.; Li, C.; Abdel-Samie, M.A.; Cui, H.; Lin, L. Unraveling the inhibitory mechanism of clove essential oil against Listeria monocytogenes biofilm and applying it to vegetable surfaces. LWT 2020, 134, 110210. [Google Scholar] [CrossRef]
- Li, M.; Lai, G.; Wang, J.; Ye, D. The inhibition of eugenol on glucan is essential for the biofilm eradication effect on caries-related biofilm in an artificial mouth model. Nat. Prod. Res. 2012, 26, 1152–1155. [Google Scholar] [CrossRef]
- Ortega-Ramirez, L.A.; Gutiérrez-Pacheco, M.M.; Vargas-Arispuro, I.; González-Aguilar, G.A.; Martínez-Téllez, M.A.; Ayala-Zavala, J.F. Inhibition of Glucosyltransferase Activity and Glucan Production as an Antibiofilm Mechanism of Lemongrass Essential Oil against Escherichia coli O157:H7. Antibiotics 2020, 9, 102. [Google Scholar] [CrossRef]
- El Hag, M.; Feng, Z.; Su, Y.; Wang, X.; Yassin, A.; Chen, S.; Peng, D.; Liu, X. Contribution of the csgA and bcsA genes to Salmonella enterica serovar Pullorum biofilm formation and virulence. Avian Pathol. 2017, 46, 541–547. [Google Scholar] [CrossRef]
- Besser, J.M. Salmonella epidemiology: A whirlwind of change. Food Microbiol. 2018, 71, 55–59. [Google Scholar] [CrossRef]
- Brenner, F.; Villar, R.; Angulo, F.; Tauxe, R.; Swaminathan, B. Salmonella nomenclature. J. Clin. Microbiol. 2000, 38, 2465–2467. [Google Scholar] [CrossRef]
- Fàbrega, A.; Vila, J. Salmonella enterica Serovar Typhimurium Skills To Succeed in the Host: Virulence and Regulation. Clin. Microbiol. Rev. 2013, 26, 308–341. [Google Scholar] [CrossRef]
- Reports of Selected Salmonella Outbreak Investigations. Available online: https://www.cdc.gov/salmonella/outbreaks.html (accessed on 7 November 2018).
- USDA. Cost Estimates of Foodborne Illnesses. Available online: https://www.ers.usda.gov/data-products/cost-estimates-of-foodborne-illnesses/ (accessed on 1 September 2021).
- CDC. Infection with Salmonella. Available online: https://www.cdc.gov/training/SIC_CaseStudy/Infection_Salmonella_ptversion.pdf (accessed on 10 September 2021).
- Ehuwa, O.; Jaiswal, A.; Jaiswal, S. Salmonella, Food Safety and Food Handling Practices. Foods 2021, 10, 907. [Google Scholar] [CrossRef]
- European Food Safety Authority; European Centre for Disease Prevention and Control. The European Union one health 2018 zoonoses report. EFSA J. 2019, 17, e05926. [Google Scholar]
- European Food Safety Authority. European Centre for Disease Prevention and Control The European Union One Health 2020 Zoonoses Report. EFSA J. 2021, 19, e06971. [Google Scholar] [CrossRef]
- DGE. Anuario de Morbilidad 1984–2017. Available online: http://www.epidemiologia.salud.gob.mx/anuario/html/anuarios.html (accessed on 15 September 2021).
- Berger, C.N.; Sodha, S.V.; Shaw, R.K.; Griffin, P.M.; Pink, D.; Hand, P.; Frankel, G. Fresh fruit and vegetables as vehicles for the transmission of human pathogens. Environ. Microbiol. 2010, 12, 2385–2397. [Google Scholar] [CrossRef] [PubMed]
- Popa, G.L.; Papa, M.I. Salmonella spp. infection-a continuous threat worldwide. Germs 2021, 11, 88. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, F.B.; Chaieb, K.; Zmantar, T.; Kallel, H.; Bakhrouf, A. Adherence assays and slime production of Vibrio alginolyticus and Vibrio parahaemolyticus. Braz. J. Microbiol. 2009, 40, 394–398. [Google Scholar] [CrossRef]
- Moraes, J.O.; Cruz, E.A.; Souza, E.G.; Oliveira, T.C.; Alvarenga, V.O.; Peña, W.E.; Sant’Ana, A.S.; Magnani, M. Predicting adhesion and biofilm formation boundaries on stainless steel surfaces by five Salmonella enterica strains belonging to different serovars as a function of pH, temperature and NaCl concentration. Int. J. Food Microbiol. 2018, 281, 90–100. [Google Scholar] [CrossRef]
- Dantas, S.T.A.; Rossi, B.F.; Bonsaglia, E.C.R.; Castilho, I.G.; Hernandes, R.T.; Fernandes, A.; Rall, V.L.M. Cross-Contamination and Biofilm Formation by Salmonella enterica Serovar Enteritidis on Various Cutting Boards. Foodborne Pathog. Dis. 2018, 15, 81–85. [Google Scholar] [CrossRef]
- Obe, T.; Richards, A.K.; Shariat, N.W. Differences in biofilm formation of Salmonella serovars on two surfaces under two temperature conditions. J. Appl. Microbiol. 2021, 132, 2410–2420. [Google Scholar] [CrossRef]
- Da Silva, F.F.M.; Monte, F.J.Q.; de Lemos, T.L.G.; Do Nascimento, P.G.G.; de Medeiros Costa, A.K.; De Paiva, L.M.M. Eugenol derivatives: Synthesis, characterization, and evaluation of antibacterial and antioxidant activities. Chem. Cent. J. 2018, 12, 34. [Google Scholar] [CrossRef]
- Kim, S.-H.; Jyung, S.; Kang, D.-H. Comparative study of Salmonella Typhimurium biofilms and their resistance depending on cellulose secretion and maturation temperatures. LWT 2022, 154, 112700. [Google Scholar] [CrossRef]
- Steenackers, H.; Hermans, K.; Vanderleyden, J.; De Keersmaecker, S.C. Salmonella biofilms: An overview on occurrence, structure, regulation and eradication. Food Res. Int. 2012, 45, 502–531. [Google Scholar] [CrossRef]
- Izadi, P.; Eldyasti, A. Holistic insights into extracellular polymeric substance (EPS) in anammosx bacterial matrix and the potential sustainable biopolymer recovery: A review. Chemosphere 2021, 274, 129703. [Google Scholar] [CrossRef] [PubMed]
- Rumbaugh, K.P.; Sauer, K. Biofilm dispersion. Nat. Rev. Microbiol. 2020, 18, 571–586. [Google Scholar] [CrossRef] [PubMed]
- Solano, C.; García, B.; Valle, J.; Berasain, C.; Ghigo, J.M.; Gamazo, C.; Lasa, I. Genetic analysis of Salmonella enteritidis biofilm formation: Critical role of cellulose. Mol. Microbiol. 2002, 43, 793–808. [Google Scholar] [CrossRef] [PubMed]
- Tursi, S.A.; Tükel, Ç. Curli-Containing Enteric Biofilms Inside and Out: Matrix Composition, Immune Recognition, and Disease Implications. Microbiol. Mol. Biol. Rev. 2018, 82, e00028-18. [Google Scholar] [CrossRef] [PubMed]
- Barnhart, M.M.; Chapman, M.R. Curli biogenesis and function. Annu. Rev. Microbiol. 2006, 60, 131–147. [Google Scholar] [CrossRef]
- Römling, U.; Galperin, M.Y. Bacterial cellulose biosynthesis: Diversity of operons, subunits, products, and functions. Trends Microbiol. 2015, 23, 545–557. [Google Scholar] [CrossRef]
- Evans, M.L.; Chapman, M.R. Curli biogenesis: Order out of disorder. Biochim. Biophys. Acta -Mol. Cell Res. 2014, 1843, 1551–1558. [Google Scholar] [CrossRef]
- Hammer, N.D.; Schmidt, J.C.; Chapman, M.R. The curli nucleator protein, CsgB, contains an amyloidogenic domain that directs CsgA polymerization. Proc. Natl. Acad. Sci. USA 2007, 104, 12494–12499. [Google Scholar] [CrossRef]
- Van Gerven, N.; Klein, R.D.; Hultgren, S.J.; Remaut, H. Bacterial Amyloid Formation: Structural Insights into Curli Biogensis. Trends Microbiol. 2015, 23, 693–706. [Google Scholar] [CrossRef]
- Robinson, L.S.; Ashman, E.M.; Hultgren, S.J.; Chapman, M.R. Secretion of curli fibre subunits is mediated by the outer membrane-localized CsgG protein. Mol. Microbiol. 2005, 59, 870–881. [Google Scholar] [CrossRef] [PubMed]
- Nenninger, A.A.; Robinson, L.S.; Hammer, N.D.; Epstein, E.A.; Badtke, M.P.; Hultgren, S.J.; Chapman, M.R. CsgE is a curli secretion specificity factor that prevents amyloid fibre aggregation. Mol. Microbiol. 2011, 81, 486–499. [Google Scholar] [CrossRef]
- Nenninger, A.A.; Robinson, L.S.; Hultgren, S.J. Localized and efficient curli nucleation requires the chaperone-like amyloid assembly protein CsgF. Proc. Natl. Acad. Sci. USA 2009, 106, 900–905. [Google Scholar] [CrossRef] [PubMed]
- Gibson, D.L.; White, A.P.; Rajotte, C.M.; Kay, W.W. AgfC and AgfE facilitate extracellular thin aggregative fimbriae synthesis in Salmonella Enteritidis. Microbiology 2007, 153, 1131–1140. [Google Scholar] [CrossRef] [PubMed]
- Zakikhany, K.; Harrington, C.R.; Nimtz, M.; Hinton, J.C.D.; Römling, U. Unphosphorylated CsgD controls biofilm formation in Salmonella enterica serovar Typhimurium. Mol. Microbiol. 2010, 77, 771–786. [Google Scholar] [CrossRef]
- Sokaribo, A.; Hansen, E.; McCarthy, M.; Desin, T.; Waldner, L.; MacKenzie, K.D.; Mutwiri, G., Jr.; Herman, N.; Herman, D.; Wang, Y.; et al. Metabolic Activation of CsgD in the Regulation of Salmonella Biofilms. Microorganisms 2020, 8, 964. [Google Scholar] [CrossRef]
- Morgan, J.L.W.; McNamara, J.T.; Zimmer, J. Mechanism of activation of bacterial cellulose synthase by cyclic di-GMP. Nat. Struct. Mol. Biol. 2014, 21, 489–496. [Google Scholar] [CrossRef]
- Omadjela, O.; Narahari, A.; Strumillo, J.; Mélida, H.; Mazur, O.; Bulone, V.; Zimmer, J. BcsA and BcsB form the catalytically active core of bacterial cellulose synthase sufficient for in vitro cellulose synthesis. Proc. Natl. Acad. Sci. USA 2013, 110, 17856–17861. [Google Scholar] [CrossRef]
- Ahmad, I.; Cimdins, A.; Beske, T.; Römling, U. Detailed analysis of c-di-GMP mediated regulation of csgD expression in Salmonella typhimurium. BMC Microbiol. 2017, 17, 27. [Google Scholar] [CrossRef]
- Sun, L.; Vella, P.; Schnell, R.; Polyakova, A.; Bourenkov, G.; Li, F.; Cimdins, A.; Schneider, T.; Lindqvist, Y.; Galperin, M.Y.; et al. Structural and Functional Characterization of the BcsG Subunit of the Cellulose Synthase in Salmonella typhimurium. J. Mol. Biol. 2018, 430, 3170–3189. [Google Scholar] [CrossRef]
- MacKenzie, K.D.; Palmer, M.B.; Köster, W.L.; White, A.P. Examining the Link between Biofilm Formation and the Ability of Pathogenic Salmonella Strains to Colonize Multiple Host Species. Front. Veter-Sci. 2017, 4, 138. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Shi, H.; Li, Y.; Shi, Z.; Zhang, X.; Baek, C.-H.; Mothershead, T.; Curtiss, R., 3rd. A colanic acid operon deletion mutation enhances induction of early antibody responses by live attenuated Salmonella vaccine strains. Infect. Immun. 2013, 81, 3148–3162. [Google Scholar] [CrossRef] [PubMed]
- Cowles, K.N.; Willis, D.K.; Engel, T.N.; Jones, J.B.; Barak, J.D. Diguanylate Cyclases AdrA and STM1987 Regulate Salmonella enterica Exopolysaccharide Production during Plant Colonization in an Environment-Dependent Manner. Appl. Environ. Microbiol. 2016, 82, 1237–1248. [Google Scholar] [CrossRef] [PubMed]
- Totani, T.; Nishiuchi, Y.; Tateishi, Y.; Yoshida, Y.; Kitanaka, H.; Niki, M.; Kaneko, Y.; Matsumoto, S. Effects of nutritional and ambient oxygen condition on biofilm formation in Mycobacterium avium subsp. hominissuis via altered glycolipid expression. Sci. Rep. 2017, 7, srep41775. [Google Scholar] [CrossRef] [PubMed]
- Barak, J.D.; Jahn, C.E.; Gibson, D.L.; Charkowski, A.O. The Role of Cellulose and O-Antigen Capsule in the Colonization of Plants by Salmonella enterica. Mol. Plant-Microbe Interact. 2007, 20, 1083–1091. [Google Scholar] [CrossRef]
- Snyder, D.S.; Gibson, D.; Heiss, C.; Kay, W.; Azadi, P. Structure of a capsular polysaccharide isolated from Salmonella enteritidis. Carbohydr. Res. 2006, 341, 2388–2397. [Google Scholar] [CrossRef]
- Giaouris, E.; Heir, E.; Hébraud, M.; Chorianopoulos, N.; Langsrud, S.; Møretrø, T.; Habimana, O.; Desvaux, M.; Renier, S.; Nychas, G.-J. Attachment and biofilm formation by foodborne bacteria in meat processing environments: Causes, implications, role of bacterial interactions and control by alternative novel methods. Meat Sci. 2014, 97, 298–309. [Google Scholar] [CrossRef]
- Moen, B.; Rudi, K.; Bore, E.; Langsrud, S. Subminimal Inhibitory Concentrations of the Disinfectant Benzalkonium Chloride Select for a Tolerant Subpopulation of Escherichia coli with Inheritable Characteristics. Int. J. Mol. Sci. 2012, 13, 4101–4123. [Google Scholar] [CrossRef]
- Wang, H.; Huang, Y.; Wu, S.; Li, Y.; Ye, Y.; Zheng, Y.; Huang, R. Extracellular DNA Inhibits Salmonella enterica Serovar Typhimurium and S. enterica Serovar Typhi Biofilm Development on Abiotic Surfaces. Curr. Microbiol. 2013, 68, 262–268. [Google Scholar] [CrossRef]
- Solomon, E.B.; Niemira, B.A.; Sapers, G.M.; Annous, B.A. Biofilm Formation, Cellulose Production, and Curli Biosynthesis by Salmonella Originating from Produce, Animal, and Clinical Sources†. J. Food Prot. 2005, 68, 906–912. [Google Scholar] [CrossRef]
- González, J.F.; Tucker, L.; Fitch, J.; Wetzel, A.; White, P.; Gunn, J.S. Human Bile-Mediated Regulation of Salmonella Curli Fimbriae. J. Bacteriol. 2019, 201, e00055-19. [Google Scholar] [CrossRef] [PubMed]
- Adcox, H.E.; Vasicek, E.M.; Dwivedi, V.; Hoang, K.V.; Turner, J.; Gunn, J.S. Salmonella Extracellular Matrix Components Influence Biofilm Formation and Gallbladder Colonization. Infect. Immun. 2016, 84, 3243–3251. [Google Scholar] [CrossRef] [PubMed]
- Lapidot, A.; Yaron, S. Transfer of Salmonella enterica Serovar Typhimurium from Contaminated Irrigation Water to Parsley Is Dependent on Curli and Cellulose, the Biofilm Matrix Components. J. Food Prot. 2009, 72, 618–623. [Google Scholar] [CrossRef] [PubMed]
- Brankatschk, K.; Kamber, T.; Pothier, J.F.; Duffy, B.; Smits, T.H. Transcriptional profile of Salmonella enterica subsp. enterica serovar W eltevreden during alfalfa sprout colonization. Microb. Biotechnol. 2014, 7, 528–544. [Google Scholar] [CrossRef]
- Yan, Z.; Yin, M.; Chen, J.; Li, X. Assembly and substrate recognition of curli biogenesis system. Nat. Commun. 2020, 11, 241. [Google Scholar] [CrossRef]
- Yaron, S.; Römling, U. Biofilm formation by enteric pathogens and its role in plant colonization and persistence. Microb. Biotechnol. 2014, 7, 496–516. [Google Scholar] [CrossRef]
- Bhowmick, P.P.; Devegowda, D.; Ruwandeepika, H.D.; Fuchs, T.M.; Srikumar, S.; Karunasagar, I.; Karunasagar, I. gcpA (stm1987) is critical for cellulose production and biofilm formation on polystyrene surface by Salmonella enterica serovar Weltevreden in both high and low nutrient medium. Microb. Pathog. 2011, 50, 114–122. [Google Scholar] [CrossRef]
- Arita-Morioka, K.-I.; Yamanaka, K.; Mizunoe, Y.; Tanaka, Y.; Ogura, T.; Sugimoto, S. Inhibitory effects of Myricetin derivatives on curli-dependent biofilm formation in Escherichia coli. Sci. Rep. 2018, 8, 8452. [Google Scholar] [CrossRef]
- Oh, S.Y.; Yun, W.; Lee, J.H.; Lee, C.H.; Kwak, W.K.; Cho, J.H. Effects of essential oil (blended and single essential oils) on anti-biofilm formation of Salmonella and Escherichia coli. J. Anim. Sci. Technol. 2017, 59, 4. [Google Scholar] [CrossRef]
- Keelara, S.; Thakur, S.; Patel, J. Biofilm Formation by Environmental Isolates of Salmonella and Their Sensitivity to Natural Antimicrobials. Foodborne Pathog. Dis. 2016, 13, 509–516. [Google Scholar] [CrossRef]
- Gill, A.; Holley, R. Disruption of Escherichia coli, Listeria monocytogenes and Lactobacillus sakei cellular membranes by plant oil aromatics. Int. J. Food Microbiol. 2006, 108, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hui, X.; Yan, G.; Tian, F.-L.; Li, H.; Gao, W.-Y. Antimicrobial mechanism of the major active essential oil compounds and their structure–activity relationship. Med. Chem. Res. 2016, 26, 442–449. [Google Scholar] [CrossRef]
- Gutierrez-Pacheco, M.; Gonzalez-Aguilar, G.; Martinez-Tellez, M.; Lizardi-Mendoza, J.; Madera-Santana, T.; Bernal-Mercado, A.; Vazquez-Armenta, F.; Ayala-Zavala, J. Carvacrol inhibits biofilm formation and production of extracellular polymeric substances of Pectobacterium carotovorum subsp. carotovorum. Food Control. 2018, 89, 210–218. [Google Scholar] [CrossRef]
- Morgan, J.L.W.; McNamara, J.T.; Fischer, M.; Rich, J.; Chen, H.-M.; Withers, S.G.; Zimmer, J. Observing cellulose biosynthesis and membrane translocation in crystallo. Nature 2016, 531, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods—A review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Number of Cases | Bacteria | Source of Contamination | Country | Year |
|---|---|---|---|---|
| 9 | Salmonella Thomson, Oraniengbur, Typhimurium, Weltevrede, Enteritidis, Hadar | Salami sticks, seafood, onions, Italian-style meats, prepackaged salads, frozen cooked shrimp, raw froze breaded stuffed chicken products, cashew brie, and ground turkey | USA | 2021 |
| 473 | Salmonella Braenderup, Muenchen, Thompson, and Typhimurium | Various | USA | 2020 |
| 1000 | Salmonella spp. | Backyard poultry | USA | 2019 |
| 80 | S. Typhimurium | Sushi | Chile | 2019 |
| 49 | Salmonella spp. | Chicken-sandwich products | Australia | 2018 |
| 40 | Salmonella Concord | Tahini products | Israel | 2018 |
| 14 | S. Typhimurium | Dehydrated coconut | USA | 2018 |
| 265 | S. Typhimurium | Chicken salad | USA | 2018 |
| 87 | Salmonella spp. | Unknown | Japan | 2017 |
| 24 | S. Typhimurium | Strains used for educational purposes | USA | 2017 |
| 907 | Salmonella Poona serotype | Cucumber | USA | 2016 |
| 230 | S. Typhimurium | Raw mung bean sprouts | South Australia | 2016 |
| 44 | S. Typhimurium | Raw breaded chicken | Canada | 2015 |
| 41 | S. Typhimurium | Strains used for educational purposes | USA | 2014 |
| 22 | S. Typhimurium | Ground beef | USA | 2013 |
| 261 | S. Typhimurium | Cantaloupe melon | USA | 2012 |
| Compound | Concentration Used against Bacteria | Mechanism Action | References |
|---|---|---|---|
| Eugenol | E. coli (800–3000 μg/mL) L. monocytogenes (800–1000 μg/mL) S. enterica serovar Thypimurium (3.18–500 μg/mL) | Inhibition: ATPase, histidine decarboxylase, and extracellular enzyme production (at sublethal concentrations) Membrane permeability ATP and potassium ion leakage | [86] |
| Eugenol | IC50 (µM) 97.31 | Deoxy-D-xylulose 5-phosphate reductoisomerase interaction with amino acids Lys124, Asp149, Ser150, Trp211, Met213, Ile217, Glu230 and Met275 (hydrogen bond with Asn226) Competitive inhibition | [87] |
| Carvacrol | IC50 (µM) 139.24 | Deoxy-D-xylulose 5-phosphate reductoisomerase interaction with amino acids Trp211, Ser212, Met213, Asp274 and Met275 (hydrogen bond with Pro273) Uncompetitive inhibition | [87] |
| Eugenol | 0.625 mg mL−1 CMI | Inhibition of glucan synthesis by Streptococcus sobrinus | [25] |
| Citral | IC50 8.5 µM E. coli | Non-competitive inhibition | [26] |
| Geraniol | IC50 6.5 µM E. coli | Non-competitive inhibition | [26] |
| Cinnamaldehyde | 2 mg/mL S. Typhimurium | Reduced biofilm population by 6 log CFU/cm2 | [85] |
| Carvacrol | 0.66 mM P. carotovorum | Reduced EPS synthesis | [88] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palomares-Navarro, J.J.; Bernal-Mercado, A.T.; González-Aguilar, G.A.; Ortega-Ramirez, L.A.; Martínez-Téllez, M.A.; Ayala-Zavala, J.F. Antibiofilm Action of Plant Terpenes in Salmonella Strains: Potential Inhibitors of the Synthesis of Extracellular Polymeric Substances. Pathogens 2023, 12, 35. https://doi.org/10.3390/pathogens12010035
Palomares-Navarro JJ, Bernal-Mercado AT, González-Aguilar GA, Ortega-Ramirez LA, Martínez-Téllez MA, Ayala-Zavala JF. Antibiofilm Action of Plant Terpenes in Salmonella Strains: Potential Inhibitors of the Synthesis of Extracellular Polymeric Substances. Pathogens. 2023; 12(1):35. https://doi.org/10.3390/pathogens12010035
Chicago/Turabian StylePalomares-Navarro, Julian J., Ariadna T. Bernal-Mercado, Gustavo A. González-Aguilar, Luis A. Ortega-Ramirez, Miguel A. Martínez-Téllez, and Jesús F. Ayala-Zavala. 2023. "Antibiofilm Action of Plant Terpenes in Salmonella Strains: Potential Inhibitors of the Synthesis of Extracellular Polymeric Substances" Pathogens 12, no. 1: 35. https://doi.org/10.3390/pathogens12010035
APA StylePalomares-Navarro, J. J., Bernal-Mercado, A. T., González-Aguilar, G. A., Ortega-Ramirez, L. A., Martínez-Téllez, M. A., & Ayala-Zavala, J. F. (2023). Antibiofilm Action of Plant Terpenes in Salmonella Strains: Potential Inhibitors of the Synthesis of Extracellular Polymeric Substances. Pathogens, 12(1), 35. https://doi.org/10.3390/pathogens12010035