
A Three-Year Look at the Phylogenetic Profile, Antimicrobial Resistance, and Associated Virulence Genes of Uropathogenic Escherichia coli


Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Consideration
2.2. Bacterial Isolates
2.3. Antimicrobial Susceptibility Assay, Detection and Confirmation of ESBLs
2.4. Hemolysin Production and Cell Surface Hydrophobicity Test
2.5. DNA Extraction and Determination of the Phylogenetic Grouping of the Isolates
2.6. Determination of Associated Virulence Factors of Strains’ UPEC Isolates
2.7. Statistical Analysis
3. Results
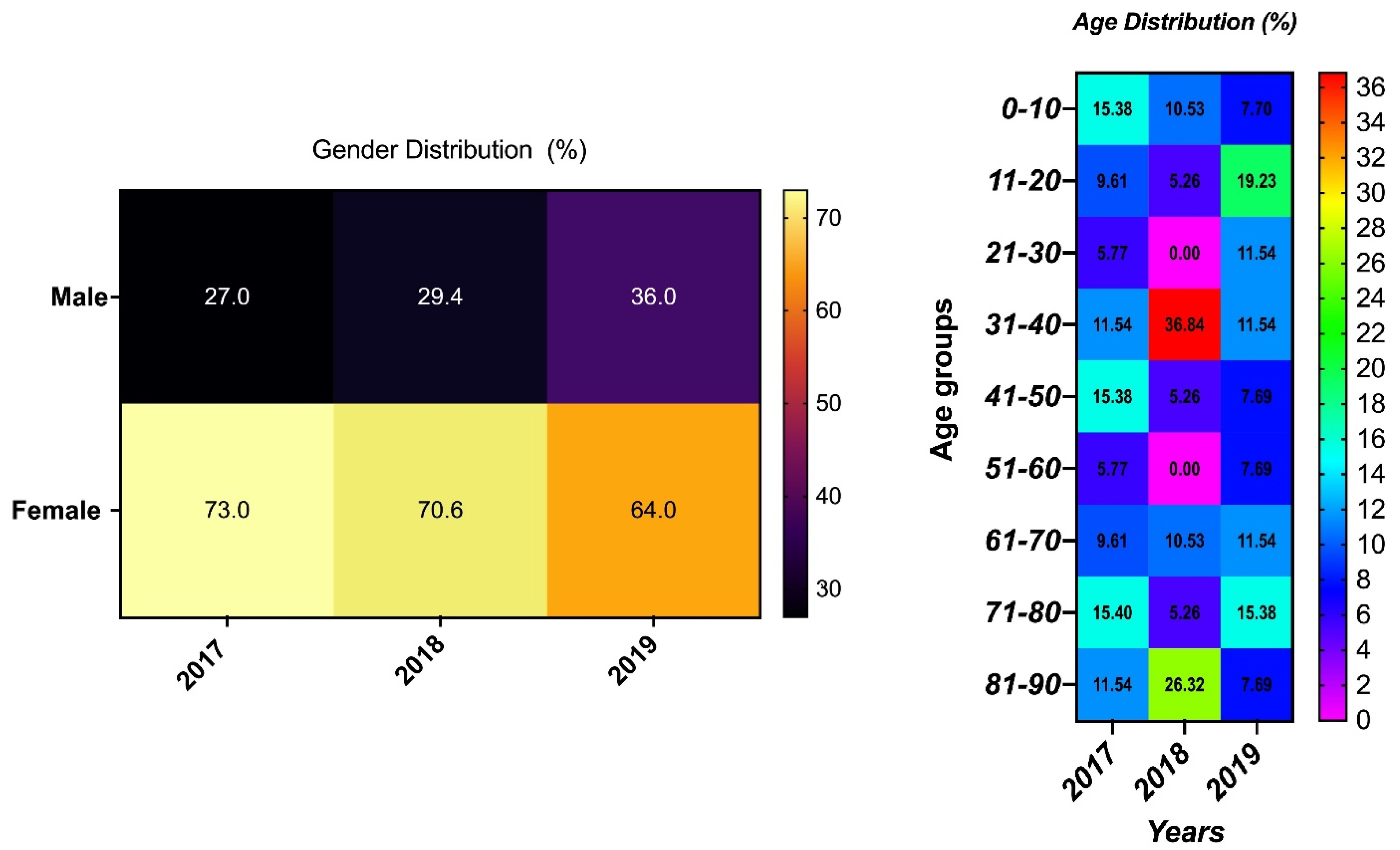
3.1. Demography and Source of Samples
3.2. Hemolysis, Cell Surface Hydrophobicity, and Phylogenetic Groups of the UPEC Isolates
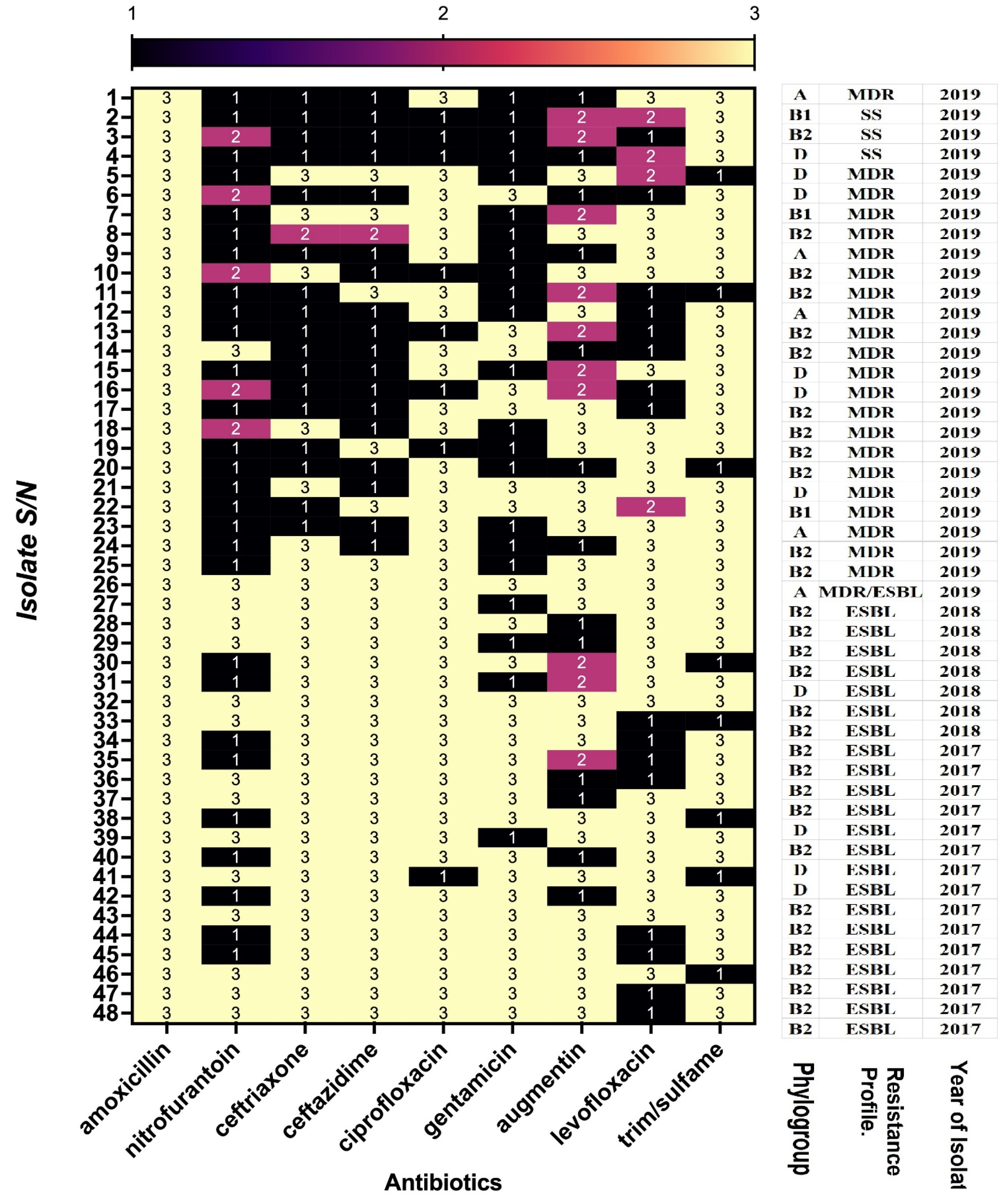
3.3. Antimicrobial Susceptibility of the Isolates
3.4. Antimicrobial Pattern of ESBLs UPEC Strains
3.5. Susceptibility to Antimicrobials According to Phylogenetic Group
3.6. Associated Virulence Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stamm, W.E.; Norrby, S.R. Urinary tract infections: Disease panorama and challenges. J. Infect. Dis. 2001, 183 (Suppl. 1), S1–S4. [Google Scholar] [CrossRef]
- Taur, Y.; Smith, M.A. Adherence to the Infectious Diseases Society of America guidelines in the treatment of uncomplicated urinary tract infection. Clin. Infect. Dis. 2007, 44, 769–774. [Google Scholar] [CrossRef]
- Schappert, S.M.; Burt, C.W. Ambulatory care visits to physician offices, hospital outpatient departments, and emergency de-partments: United States, 2001–2002. Vital. Health Stat. 2006, 13, 1–66. [Google Scholar]
- Kostakioti, M.; Hultgren, S.J.; Hadjifrangiskou, M. Molecular blueprint of uropathogenic Escherichia coli virulence provides clues toward the development of anti-virulence therapeutics. Virulence 2012, 3, 592–594. [Google Scholar] [CrossRef] [PubMed]
- Tandogdu, Z.; Wagenlehner, F.M. Global epidemiology of urinary tract infections. Curr. Opin. Infect. Dis. 2016, 29, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Alanazi, M.Q.; Al-Jeraisy, M.I.; Salam, M. Prevalence and predictors of antibiotic prescription errors in an emergency de-partment, Central Saudi Arabia. Drug Healthc. Patient Saf. 2015, 7, 103–111. [Google Scholar] [CrossRef]
- Car, J. Urinary tract infections in women: Diagnosis and management in primary care. BMJ 2006, 332, 94–97. [Google Scholar] [CrossRef]
- Spurbeck, R.R.; Mobley, H.L.T. Chapter 9—Uropathogenic Escherichia coli. In Donnenberg, Escherichia coli, 2nd ed.; Michael, S., Ed.; Elsevier Inc., Academic Press: Amsterdam, The Netherlands, 2013; pp. 275–304. ISBN 9780123970480. [Google Scholar] [CrossRef]
- Alanazi, M.Q. An evaluation of community-acquired urinary tract infection and appropriateness of treatment in an emergency department in Saudi Arabia. Ther. Clin. Risk Manag. 2018, 14, 2363–2373. [Google Scholar] [CrossRef]
- Akbar, D.H. Urinary tract infection. Diabetics and non-diabetic patients. Saudi Med. J. 2001, 22, 326–329. [Google Scholar]
- General Authority for Statistics. Statistical Yearbook of 2016. In Kingdom of Saudi Arabia: General Authority for Statistics; 2016. Available online: https://www.stats.gov.sa/en/169 (accessed on 3 May 2022).
- Garout, W.A.; Kurdi, H.S.; Shilli, A.H.; Kari, J.A. Urinary tract infection in children younger than 5 years. Etiology and asso-ciated urological anomalies. Saudi Med. J. 2015, 36, 497–501. [Google Scholar] [CrossRef]
- Yadav, K.K.; Adhikari, N.; Khadka, R.; Pant, A.D.; Shah, B. Multidrug resistant Enterobacteriaceae and extended spectrum β-lactamase producing Escherichia coli: A cross-sectional study in National Kidney Center, Nepal. Antimicrob Resist. Infect Control. 2015, 4, 42. [Google Scholar] [CrossRef] [PubMed]
- Gharavi, M.J.; Zarei, J.; Roshani-Asl, P.; Yazdanyar, Z.; Sharif, M.; Rashidi, N. Comprehensive study of antimicrobial suscep-tibility pattern and extended spectrum beta-lactamase (ESBL) prevalence in bacteria isolated from urine samples. Sci. Rep. 2021, 11, 578. [Google Scholar] [CrossRef] [PubMed]
- Norafika, N.A.; Arbianti, N.; Prihatiningsih, S.; Indriani, D.W. A retrospective cross-sectional study of urinary tract infections and prevalence of antibiotic resistant pathogens in patients with diabetes mellitus from a public hospital in Surabaya, Indonesia. Germs 2020, 10, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Alasmary, M.Y. Antimicrobial Resistance Patterns and ESBL of Uropathogens Isolated from Adult Females in Najran Region of Saudi Arabia. Clin. Pract. 2021, 11, 650–658. [Google Scholar] [CrossRef]
- Alqasim, A.; Abu Jaffal, A.; Alyousef, A.A. Prevalence of Multidrug Resistance and Extended-Spectrum β-Lactamase Carriage of Clinical Uropathogenic Escherichia coli Isolates in Riyadh, Saudi Arabia. Int. J. Microbiol. 2018, 2018, 3026851. [Google Scholar] [CrossRef]
- Bien, J.; Sokolova, O.; Bozko, P. Role of Uropathogenic Escherichia coli Virulence Factors in Development of Urinary Tract Infection and Kidney Damage. Int. J. Nephrol 2012, 2012, 681473. [Google Scholar] [CrossRef]
- Schreiber, H.L.; Conover, M.S.; Chou, W.C.; Hibbing, M.E.; Manson, A.L.; Dodson, K.W.; Hannan, T.J.; Roberts, P.L.; Stapleton, A.E.; Hooton, T.M.; et al. Bacterial virulence phenotypes of Escherichia coli and host susceptibility determine risk for urinary tract infections. Sci. Transl. Med. 2017, 9, eaaf1283. [Google Scholar] [CrossRef]
- Asadi, S.; Kargar, M.; Solhjoo, K.; Najafi, A.; Ghorbani-Dalini, S. The Association of Virulence Determinants of Uropathogenic Escherichia coli With Antibiotic Resistance. Jundishapur J. Microbiol. 2014, 7, e9936. [Google Scholar] [CrossRef]
- Shah, C.; Baral, R.; Bartaula, B.; Shrestha, L.B. Virulence factors of uropathogenic Escherichia coli (UPEC) and correlation with antimicrobial resistance. BMC Microbiol. 2019, 19, 204. [Google Scholar] [CrossRef]
- Alghoribi, M.F.; Gibreel, T.M.; Farnham, G.; Al-Johani, S.M.; Balkhy, H.H.; Upton, M. Antibiotic-resistant ST38, ST131 and ST405 strains are the leading uropathogenic Escherichia coli clones in Riyadh, Saudi Arabia. J. Antimicrob. Chemother 2015, 70, 2757–2762. [Google Scholar] [CrossRef]
- Johnson, J.R.; Kuskowski, M.A.; Owens, K.; Gajewski, A.; Winokur, P.L. Phylogenetic origin and virulence genotype in relation to resistance to fluoroquinolones and/or extended-spectrum cephalosporins and cephamycins among Escherichia coli isolates from animals and humans. J. Infect. Dis. 2003, 188, 759–768. [Google Scholar] [CrossRef]
- Alqasim, A.; Abu Jaffal, A.; Alyousef, A.A. Prevalence and molecular characteristics of sequence type 131 clone among clinical uropathogenic Escherichia coli isolates in Riyadh, Saudi Arabia. Saudi J. Biol. Sci. 2020, 27, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Bazaid, A.S.; Saeed, A.; Alrashidi, A.; Alshaghdali, K.; A Hammam, S.; Alreshidi, T.; Alshammary, M.; Alarfaj, A.; Thallab, R.; Aldarhami, A. Antimicrobial Surveillance for Bacterial Uropathogens in Hail, Saudi Arabia: A Five-Year Multi-center Retrospective Study. Infect. Drug Resist. 2021, 14, 1455–1465. [Google Scholar] [CrossRef] [PubMed]
- Al Yousef, S.A.; Younis, S.; Farrag, E.; Moussa, H.S.h.; Bayoumi, F.S.; Ali, A.M. Clinical and Laboratory Profile of Urinary Tract Infections Associated with Extended Spectrum β-Lactamase Producing Escherichia coli and Klebsiella pneumoniae. Ann. Clin. Lab Sci. 2016, 46, 393–400. [Google Scholar] [PubMed]
- Al-Tawfiq, J.A.; Anani, A.A. Antimicrobial susceptibility pattern of bacterial pathogens causing urinary tract infections in a Saudi Arabian hospital. Chemotherapy 2009, 55, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Al-Harthi, A.A.; Al-Fifi, S.H. Antibiotic resistance pattern and empirical therapy for urinary tract infections in children. Saudi Med. J. 2008, 29, 854–858. [Google Scholar]
- Kader, A.A.; Kumar, A. Prevalence and antimicrobial susceptibility of extended-spectrum beta-lactamase-producing Esche-richia coli and Klebsiella pneumoniae in a general hospital. Ann. Saudi. Med. 2005, 25, 239–242. [Google Scholar] [CrossRef]
- Eltahawy, A.T.; Khalaf, R.M.F. Urinary tract infection at a University Hospital in Saudi Arabia: Incidence, microbiology, and antimicrobial susceptibility. Ann. Saudi Med. 1988, 8, 261–266. [Google Scholar] [CrossRef][Green Version]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; CLSI supplement M100; Clinical and La-bor-atory Standards Institute: Pittsburgh, PA, USA, 2020; Available online: https://www.nih.org.pk/wp-content/uploads/2021/02/CLSI2020.pdf (accessed on 1 August 2021).
- Clinical Laboratory Standards Institute. Methods for Dilution Antimicrobial Susceptibility Testing for Bacteria That Grows Aerobically, 9th ed.; (M07-A9); Clinical Laboratory Standards Institute: Wayne, PA, USA, 2012. [Google Scholar]
- Ranjan, K.P.; Ranjan, N.; Chakraborty, A.; Arora, D.R. An approach to uropathogenic Escherichia coli in urinary tract infec-tions. J. Lab Physicians 2010, 2, 70–73. [Google Scholar] [CrossRef]
- Siegfried, L.; Kmetová, M.; Puzová, H.; Molokácová, M.; Filka, J. Virulence-associated factors in Escherichia coli strains isolated from children with urinary tract infections. J. Med. Microbiol. 1994, 41, 127–132. [Google Scholar] [CrossRef]
- Clermont, O.; Bonacorsi, S.; Bingen, E. Rapid and simple determination of the Escherichia coli phylogenetic group. Appl. Environ. Microbiol. 2000, 66, 4555–4558. [Google Scholar] [CrossRef] [PubMed]
- Le Bouguenec, C.; Archambaud, M.; Labigne, A. Rapid and specific detection of the pap, afa, and sfa adhesin-encoding operons in uropathogenic Escherichia coli strains by polymerase chain reaction. J. Clin. Microbiol. 1992, 30, 1189–1193. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; Terai, A.; Yuri, K.; Kurazono, H.; Takeda, Y.; Yoshida, O. Detection of urovirulence factors in Escherichia coli by multiplex polymerase chain reaction. FEMS Immunol. Med. Microbiol. 1995, 12, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.R.; Stell, A.L. Extended virulence genotypes of Escherichia coli strains from patients with urosepsis in relation to phylogeny and host compromise. J. Infect. Dis. 2000, 181, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.R.; Russo, T.A.; Tarr, P.I.; Carlino, U.; Bilge, S.S.; Vary, J.C.; Stell, A.L. Molecular epidemiological and phylogenetic associations of two novel putative virulence genes, iha and iroN (E. coli), among Escherichia coli isolates from patients with urosepsis. Infect. Immun. 2000, 68, 3040–3047. [Google Scholar] [CrossRef]
- Dadi, B.R.; Abebe, T.; Zhang, L.; Mihret, A.; Abebe, W.; Amogne, W. Distribution of virulence genes and phylogenetics of uropathogenic Escherichia coli among urinary tract infection patients in Addis Ababa, Ethiopia. BMC Infect. Dis. 2020, 20, 108. [Google Scholar] [CrossRef] [PubMed]
- Mazumder, R.; Abdullah, A.; Ahmed, D.; Hussain, A. High Prevalence of blaCTX-M-15 Gene among Extended-Spectrum β-Lactamase-Producing Escherichia coli Isolates Causing Extraintestinal Infections in Bangladesh. Antibiotics 2020, 9, 796. [Google Scholar] [CrossRef]
- Lee, J.H.; Subhadra, B.; Son, Y.J.; Kim, D.H.; Park, H.S.; Kim, J.M.; Koo, S.H.; Oh, M.H.; Kim, H.J.; Choi, C.H. Phylogenetic group distributions, virulence factors and antimicrobial resistance properties of uropathogenic Escherichia coli strains isolated from patients with urinary tract infections in South Korea. Lett. Appl. Microbiol. 2016, 62, 84–90. [Google Scholar] [CrossRef]
- Iranpour, D.; Hassanpour, M.; Ansari, H.; Tajbakhsh, S.; Khamisipour, G.; Najafi, A. Phylogenetic groups of Escherichia coli strains from patients with urinary tract infection in Iran based on the new Clermont phylotyping method. BioMed Res. Int. 2015, 2015, 846219. [Google Scholar] [CrossRef]
- Al-Zahrani, J.; Al Dossari, K.; Gabr, A.H.; Ahmed, A.-F.; Al Shahrani, S.A.; Al-Ghamdi, S. Antimicrobial resistance patterns of Uropathogens isolated from adult women with acute uncomplicated cystitis. BMC Microbiol. 2019, 19, 237. [Google Scholar] [CrossRef]
- Al Wutayd, O.; Al Nafeesah, A.; Adam, I.; Babikir, I. The antibiotic susceptibility patterns of uropathogens isolated in Qassim, Saudi Arabia. J. Infect. Dev. Ctries. 2018, 12, 946–952. [Google Scholar] [CrossRef] [PubMed]
- Haseeb, A.; Faidah, H.S.; Al-Gethamy, M.; Iqbal, M.S.; Barnawi, A.M.; Elahe, S.S.; Bukhari, D.N.; Noor Al-Sulaimani, T.M.; Fadaaq, M.; Alghamdi, S.; et al. Evaluation of a Multidisciplinary Antimicrobial Stewardship Program in a Saudi Critical Care Unit: A Quasi-Experimental Study. Front. Pharmacol. 2020, 11, 570238. [Google Scholar] [CrossRef] [PubMed]
- Al-Omari, A.; Al Mutair, A.; Alhumaid, S.; Salih, S.; Alanazi, A.; Albarsan, H.; Abourayan, M.; Al Subaie, M. The impact of antimicrobial stewardship program implementation at four tertiary private hospitals: Results of a five-years pre-post analysis. Antimicrob Resist. Infect. Control 2020, 9, 95. [Google Scholar] [CrossRef] [PubMed]
- Zowawi, H.M. Antimicrobial resistance in Saudi Arabia. An urgent call for an immediate action. Saudi Med. J. 2016, 37, 935–940. [Google Scholar] [CrossRef]
- Alghamdi, S.; Berrou, I.; Aslanpour, Z.; Mutlaq, A.; Haseeb, A.; Albanghali, M.; Hammad, M.A.; Shebl, N. Antimicrobial Stewardship Programmes in Saudi Hospitals: Evidence from a National Survey. Antibiotics 2021, 10, 193. [Google Scholar] [CrossRef]
- AlOtieschan, S.; Alsalim, A.; Albabtain, S.; Almujahid, M.; Obeid, D.; Alhabradi, F.; Alenazi, T. Public Perception Toward Ministry of Health Regulations for Antibiotic Dispensing and Its Impact on Pharmacy and Family Physician Visits. Cureus 2021, 13, e14981. [Google Scholar] [CrossRef]
- Bartoletti, R.; Cai, T.; Wagenlehner, F.M.; Naber, K.; Bjerklund, J.T.E. Treatment of urinary tract infections and antibiotic stewardship. Eur. Urol. Suppl. 2016, 15, 81–87. [Google Scholar] [CrossRef]
- Kot, B. Antibiotic Resistance Among Uropathogenic Escherichia coli. Pol. J. Microbiol 2019, 68, 403–415. [Google Scholar] [CrossRef]
- Adamus-Białek, W.; Baraniak, A.; Wawszczak, M.; Głuszek, S.; Gad, B.; Wróbel, K.; Bator, P.; Majchrzak, M.; Parniewski, P. The genetic background of antibiotic resistance among clinical uropathogenic Escherichia coli strains. Mol. Biol. Rep. 2018, 45, 1055–1065. [Google Scholar] [CrossRef]
- Delpech, G.; Allende, N.G.; Lissarrague, S.; Sparo, M. Antimicrobial resistance of uropathogenic Escherichia coli from elderly patients at a general hospital, Argentina. Open Infect. Dis. J. 2018, 10, 79–87. [Google Scholar] [CrossRef]
- Kresken, M.; Körber-Irrgang, B.; Biedenbach, D.J.; Batista, N.; Besard, V.; Cantón, R.; García-Castillo, M.; Kalka-Moll., W.; Pascual, A.; Schwarz, R.; et al. Comparative in vitro activity of oral antimicrobial agents against Enterobacteriaceae from patients with community acquired urinary tract infections in three European countries. Clin. Microbiol. Infect. 2016, 22, 63.e1–63.e5. [Google Scholar] [CrossRef] [PubMed]
- Cunha, M.A.; Assunção, G.L.M.; Medeiros, I.M.; Freitas, M.R. Antibiotic resistance patterns of urinary tract infections in a north-eastern Brazilian capital. Rev. Inst. Med. Trop São Paulo 2016, 58, 2. [Google Scholar] [CrossRef] [PubMed]
- Abernethy, J.; Guy, R.; Sheridan, E.A.; Hopkins, S.; Kiernan, M.; Wilcox, M.H.; Johnson, A.P.; Hope, R.E. coli bacteraemia sentinel surveillance group. Epidemiology of Escherichia coli bacteraemia in England: Results of an enhanced sentinel surveil-lance programme. J. Hosp. Infect. 2017, 95, 365–375. [Google Scholar] [CrossRef]
- Abduzaimovic, A.; Aljicevic, M.; Rebic, V.; Vranic, S.M.; Abduzaimovic, K.; Sestic, S. Antibiotic Resistance in Urinary Isolates of Escherichia coli. Mater Sociomed 2016, 28, 416–419. [Google Scholar] [CrossRef] [PubMed]
- Yamaji, R.; Rubin, J.; Thys, E.; Friedman, C.R.; Riley, L.W. Persistent Pandemic Lineages of Uropathogenic Escherichia coli in a College Community from 1999 to 2017. J. Clin. Microbiol. 2018, 56, e01834–e17. [Google Scholar] [CrossRef] [PubMed]
- Prasada, S.; Bhat, A.; Bhat, S.; Shenoy Mulki, S.; Tulasidas, S. Changing antibiotic susceptibility pattern in uropathogenic Escherichia coli over a period of 5 years in a tertiary care center. Infect. Drug Resist. 2019, 12, 1439–1443. [Google Scholar] [CrossRef]
- Neuner, E.A.; Sekeres, J.; Hall, G.S.; van Duin, D. Experience with fosfomycin for treatment of urinary tract infections due to multidrug-resistant organisms. Antimicrob. Agents Chemother 2012, 56, 5744–5748. [Google Scholar] [CrossRef]
- Balkhi, B.; Mansy, W.; Alghadeer, S.; Alnuaim, A.; AlShehri, A.; Somily, A. Antimicrobial susceptibility of microorganisms causing Urinary Tract Infections in Saudi Arabia. J. Infect. Dev. Ctries. 2018, 12, 220–227. [Google Scholar] [CrossRef]
- Alyamani, E.J.; Khiyami, A.M.; Booq, R.Y.; Majrashi, M.A.; Bahwerth, F.S.; Rechkina, E. The occurrence of ESBL-producing Escherichia coli carrying aminoglycoside resistance genes in urinary tract infections in Saudi Arabia. Ann. Clin. Microbiol. An-timicrob. 2017, 16, 1–13. [Google Scholar] [CrossRef]
- Hawkey, P.M. Prevalence and clonality of extended-spectrum beta-lactamases in Asia. Clin. Microbiol. Infect. 2008, 14 (Suppl. 1), 159–165. [Google Scholar] [CrossRef]
- Usein, C.R.; Damian, M.; Tatu-Chitoiu, D.; Capusa, C.; Fagaras, R.; Tudorache, D.; Nica, M.; Le Bouguénec, C. Prevalence of virulence genes in Escherichia coli strains isolated from Romanian adult urinary tract infection cases. J. Cell Mol. Med. 2001, 5, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhao, S.; Han, L.; Guo, X.; Chen, M.; Ni, Y.; Zhang, Y.; Cui, Z.; He, P. Drug resistance and virulence of uropa-tho-genic Escherichia coli from Shanghai, China. J. Antibiot (Tokyo) 2014, 67, 799–805. [Google Scholar] [CrossRef] [PubMed]
- Munkhdelger, Y.; Gunregjav, N.; Dorjpurev, A.; Juniichiro, N.; Sarantuya, J. Detection of virulence genes, phylogenetic group and antibiotic resistance of uropathogenic Escherichia coli in Mongolia. J. Infect. Dev. Ctries 2017, 11, 51–57. [Google Scholar] [CrossRef]
- Khairy, R.M.; Mohamed, E.S.; Ghany, H.M.A.; Abdelrahim, S.S. Phylogenic classification and virulence genes profiles of uropathogenic E. coli and diarrheagenic E. coli strains isolated from community acquired infections. PLoS ONE 2019, 14, e0222441. [Google Scholar] [CrossRef]
- Tan, C.W.; Chlebicki, M.P. Urinary tract infections in adults. Singapore Med. J. 2016, 57, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Momtaz, H.; Karimian, A.; Madani, M.; Safarpoor Dehkordi, F.; Ranjbar, R.; Sarshar, M.; Souod, N. Uropathogenic Escherichia coli in Iran: Serogroup distributions, virulence factors and antimicrobial resistance properties. Ann. Clin. Microbiol. Antimicrob 2013, 12, 8. [Google Scholar] [CrossRef] [PubMed]
- Tabasi, M.; Karam, M.R.; Habibi, M.; Mostafavi, E.; Bouzari, S. Genotypic Characterization of Virulence Factors in Escherichia coli Isolated from Patients with Acute Cystitis, Pyelonephritis and Asymptomatic Bacteriuria. J. Clin. Diagn Res. 2016, 10, DC01–DC07. [Google Scholar] [CrossRef]
- Tarchouna, M.; Ferjani, A.; Ben-Selma, W.; Boukadida, J. Distribution of uropathogenic virulence genes in Escherichia coli isolated from patients with urinary tract infection. Int. J. Infect. Dis. 2013, 17, e450–e453. [Google Scholar] [CrossRef]
- Wiles, T.J.; Kulesus, R.R.; Mulvey, M.A. Origins and virulence mechanisms of uropathogenic Escherichia coli. Exp. Mol. Pathol. 2008, 85, 11–19. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Genes | Primer Sequence | BP | Reference |
|---|---|---|---|
| ChuA | F-GACGAACCAACGGTCAGGAT R-TGCCGCCAGTACCAAAGACA | 279 | [35] |
| yjaA | F-TGAAGTGTCAGGAGACGCTG R-ATGGAGAATGCGTTCCTCAAC | 211 | [35] |
| TspE4C2 | F-GAGTAATGTCGGGGCATTCA R-CGCGCCAACAAAGTATTACG | 154 | [35] |
| fimH | F: TGCAGAACGGATAAGCCGTGG R: GCAGTCACCTGCCCTCCGGTA | 506 | [36] |
| Sfa (sfa/foc) | F: CTCCGGAGAACTGGGTGCATCTTAC R: CGGAGGAGTAATTACAAACCTGGCA | 410 | [37] |
| papA | F: ATGGCAGTGGTGTTTTGGTG R: CGTCCCACCATACGTGCTCTTC | 720 | [37] |
| papE/F | F: GCAACAGCAACGCTGGTTGCATCAT R: AGAGAGAGCCACTCTTATACGGACA | 336 | [38] |
| hlyA | F: AACAAGGATAAGCACTGTTCTGGCT R: ACCATATAAGCGGTCATTCCCGTCA | 1170 | [38] |
| iroN | F AAGTCAAAGCAGGGGTTGCCCG R-GACGCCGACATTAAGACGCAG | 665 | [39] |
| S/N | Lab ID | Genes/Phylogenetic Group | Virulence Genes | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| chuA | yjaA | TspE4.C2 | Group | sfa | fimH | papE/F | iroN | papA | hlyA | ||
| 1 | AC12 | − | − | A | − | − | + | − | − | − | |
| 2 | AC13 | − | + | B1 | − | − | − | − | − | − | |
| 3 | AC14 | + | + | B2 | − | + | − | − | − | − | |
| 4 | AC15 | + | − | D | − | − | − | − | − | − | |
| 5 | AC16 | + | − | D | − | − | − | − | − | − | |
| 6 | AC17 | + | − | D | − | − | − | − | − | − | |
| 7 | AC18 | − | + | B1 | − | − | − | − | − | − | |
| 8 | AC19 | + | + | B2 | − | − | + | − | − | − | |
| 9 | AC20 | − | − | A | − | − | − | − | − | ||
| 10 | AC21 | + | + | B2 | − | − | + | − | − | − | |
| 11 | AC22 | + | + | B2 | − | + | − | − | − | − | |
| 12 | AC23 | − | − | A | − | − | − | − | − | − | |
| 13 | AC24 | + | + | B2 | − | + | − | − | − | − | |
| 14 | AC25 | + | + | B2 | − | + | − | − | − | − | |
| 15 | AC26 | + | − | D | − | − | − | − | − | − | |
| 16 | AC27 | + | − | D | − | − | − | − | − | − | |
| 17 | AC28 | + | + | B2 | − | + | − | − | − | − | |
| 18 | AC29 | + | + | B2 | − | − | + | − | − | − | |
| 19 | AC30 | + | + | B2 | − | + | − | − | − | − | |
| 20 | AC31 | + | + | B2 | − | + | − | − | − | − | |
| 21 | AC32 | + | − | D | − | + | − | − | − | − | |
| 22 | AC33 | − | + | B1 | − | − | − | − | − | − | |
| 23 | AC34 | − | − | A | − | − | − | − | − | − | |
| 24 | AC35 | + | + | B2 | − | + | − | − | − | − | |
| 25 | AC36 | + | + | B2 | − | + | − | − | − | − | |
| 26 | AC37 | − | − | A | − | − | − | − | − | − | |
| 27 | LAB07 | + | + | B2 | − | − | + | − | − | − | |
| 28 | LAB31 | + | + | B2 | − | − | − | − | − | − | |
| 29 | LAB40 | + | + | B2 | − | + | − | − | − | − | |
| 30 | LAB63 | + | + | B2 | − | + | − | − | − | − | |
| 31 | LAB71 | + | − | D | − | − | − | − | − | − | |
| 32 | LAB87 | + | + | B2 | − | + | − | − | − | − | |
| 33 | LAB91 | + | + | B2 | − | − | + | − | − | − | |
| 34 | LAB114 | + | + | B2 | − | − | − | − | − | − | |
| 35 | LAB119 | + | + | B2 | − | + | − | − | − | − | |
| 36 | LAB122 | + | + | B2 | − | − | − | − | − | − | |
| 37 | LAB124 | + | + | B2 | − | − | − | − | − | − | |
| 38 | LAB130 | + | − | D | − | + | − | − | − | − | |
| 39 | LAB140 | + | + | B2 | − | − | + | − | − | − | |
| 40 | LAB149 | + | − | D | − | + | − | − | − | − | |
| 41 | LAB155 | + | − | D | − | + | − | − | − | − | |
| 42 | LAB158 | + | + | B2 | − | − | − | − | − | − | |
| 43 | LAB168 | + | + | B2 | − | + | − | − | − | − | |
| 44 | LAB186 | + | + | B2 | − | − | − | − | − | − | |
| 45 | LAB188 | + | + | B2 | − | + | − | − | − | − | |
| 46 | LAB193 | + | + | B2 | − | + | − | − | − | − | |
| 47 | LAB194 | + | + | B2 | − | + | − | − | − | − | |
| 48 | LAB195 | + | + | B2 | − | + | − | − | − | − | |
| Antimicrobial Classes | Antimicrobials | Year of Isolation | % R Pairwise Comparison (p Value) | % I Pairwise Comparison (p Value) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2017 | 2018 | 2019 | 2017 vs. 2018 | 2017 vs. 2019 | 2018 vs. 2019 | 2017 vs. 2018 | 2017 vs. 2019 | 2018 vs. 2019 | |||||
| % R | % I | % R | % I | % R | % I | ||||||||
| Penicillins | Aml | np | np | 88 | 0 | 100 | 0 | na | na | <0.01 | na | na | - |
| Am | 100 | 0 | 100 | 0 | 100 | 0 | - | - | - | - | - | - | |
| Aug | 73.08 | 0 | 64.7 | 0 | 46 | 27 | 0.20 | <0.01 | 0.01 | - | <0.01 | <0.01 | |
| Ptz | np | np | 35.3 | 0 | 23 | 4 | na | na | 0.06 | na | na | 0.04 | |
| Cephalosporins | Cfz | 100 | 0 | 100 | 0 | np | np | - | na | na | - | na | na |
| Cft | 100 | 0 | 94.1 | 0 | 30.8 | 7.7 | 0.01 | <0.01 | <0.01 | - | <0.01 | <0.01 | |
| Cxm | 100 | 0 | 100 | 0 | 89 | 0 | - | <0.01 | <0.01 | - | - | - | |
| Kf | 100 | 0 | 100 | 0 | 57.7 | 27 | - | <0.01 | <0.01 | - | <0.01 | <0.01 | |
| Fox | np | np | np | np | 38.46 | 0 | na | na | na | na | na | na | |
| Caz | 100 | 0 | 94.1 | 0 | 26.92 | 0 | 0.01 | <0.01 | <0.01 | - | - | - | |
| Cax | 100 | 0 | 96 | 0 | 27 | 0 | 0.04 | <0.01 | <0.01 | - | - | - | |
| Pime | 100 | 0 | 94.1 | 0 | 15.4 | 0 | 0.01 | <0.01 | <0.01 | - | - | - | |
| Carbapenems | Imi | 4.6 | 0 | 12 | 0 | 0 | 8 | 0.06 | 0.03 | <0.01 | - | <0.01 | <0.01 |
| Mer | 2 | 0 | 0 | 0 | 4 | 4 | 0.16 | 0.41 | 0.04 | - | 0.04 | 0.04 | |
| Aminoglycosides | Amk | 11 | 2 | 23.5 | 0 | 4 | 4 | 0.02 | 0.06 | <0.01 | 0.16 | 0.41 | 0.04 |
| Gm | 66 | 0 | 53 | 0 | 27 | 0 | 0.06 | <0.01 | <0.01 | - | - | - | |
| Fluroquinolones | Cp | 85.7 | 0 | 70.5 | 0 | 73 | 0 | 0.01 | 0.03 | 0.69 | - | - | - |
| Levo | 81.4 | 0 | 76.5 | 0 | 90 | 0 | 0.40 | 0.08 | 0.01 | - | - | - | |
| Others | Tig | 0 | 0 | 0 | 0 | 0 | 0 | - | - | - | - | - | - |
| Fd | 35 | 0 | 59 | 0 | 8 | 19 | <0.01 | <0.01 | <0.01 | - | <0.01 | <0.01 | |
| Sxt | 72.55 | 0 | 75 | 0 | 84.6 | 0 | 0.69 | 0.04 | 0.09 | - | - | - | |
| Azt | 73 | 0 | 100 | 0 | 75 | 0 | <0.01 | 0.75 | <0.01 | - | - | - | |
| C | 25 | 7 | 7 | 25 | np | np | <0.01 | na | na | <0.01 | na | na | |
| Pip | 100 | 0 | 100 | 0 | np | np | - | na | na | - | na | na | |
| Te | 79.6 | 0 | 64.7 | 0 | 65 | 0 | 0.02 | 0.02 | 0.96 | - | - | - | |
| ESBL Test | Year of Isolation | ||
|---|---|---|---|
| 2017 | 2018 | 2019 | |
| N = 52 (%) | N = 50 (%) | N = 50 (%) | |
| CTX/CLA | 46 (88.5) | 20 (40) | 2 (4) |
| CAZ/CLA | 44 (85) | 15 (30) | 1 (2) |
| Negative for CTX/CAZ/CLA | 6 (11.5) | 0 (0) | 0 (0) |
| Associated extended-spectrum beta-lactamases (ESBL) genes | |||
| ctx | 44 (85) | 15 (30) | 0 (0) |
| shv | 13 (25) | 10 (20) | 0 (0) |
| tem | 29 (56) | 6 (12) | 0 (0) |
| ctx/shv | 13 (25) | 3 (6) | 1 (2) |
| ctx/tem | 18 (34.6) | 5 (10) | 0 (0) |
| shv/tem | 1 (2) | 0 (0) | 0 (0) |
| ctx/shv/tem | 8 (15.4) | 3 (6) | 1 (2) |
| None | 8 (15.4) | 1 (2) | 0 (0) |
| Antibiotics | Phylogroups, N (%) | Comparison of % R across Phylogroups X2, p Value | |||||||
|---|---|---|---|---|---|---|---|---|---|
| A N = 18 | B1 N = 11 | B2 N = 106 | D N = 35 | ||||||
| R % | I % | R % | I % | R % | I % | R % | I % | ||
| Amoxicillin | 18 (100) | 0 (0) | 11 (100) | 0 (0) | 106 (100) | 0 (0) | 35 (100) | 0 (0) | na |
| Augmentin | 11 (60) | 0 (0) | 4 (36) | 7 (64) | 64 (60.4) | 14 (13.2) | 14 (40) | 11 (31) | 58.32, <0.001 |
| Ceftriaxone | 14 (78) | 4 (22) | 8 (73) | 0 (0) | 74 (70) | 6 (6) | 21 (60) | 2 (6) | 2.07, 0.558 |
| Ceftazidime | 0 (0) | 0 (0) | 8 (73) | 0 (0) | 67 (63) | 7 (7) | 14 (40) | 0 (0) | 28.76, <0.001 |
| Ciprofloxacin | 18 (100) | 0 (0) | 7 (64) | 0 (0) | 82 (77) | 0 (0) | 25 (71) | 0 (0) | 0.001 |
| Gentamicin | 4 (22) | 0 (0) | 8 (73) | 0 (0) | 53 (50) | 0 (0) | 25 (71) | 0 (0) | 13.72, 0.003 |
| Nitrofurantoin | 4 (22) | 0 (0) | 0 (0) | 0 (0) | 25 (24) | 11 (10) | 11 (31) | 0 (0) | 0.15 |
| Levofloxacin | 11 (61) | 0 (0) | 7 (64) | 4 (36) | 53 (50) | 14 (13) | 14 (40) | 14 (40) | 3.11, 0.375 |
| Trimethoprim/ Sulfamethoxazole | 11 (61) | 0 (0) | 11 (100) | 0 (0) | 96 (91) | 0 (0) | 25 (71) | 0 (0) | 0.01 |
| Tobramycin | 17 (94) | 0 (0) | 8 (73) | 0 (0) | 98 (93) | 0 (0) | 29 (83) | 0 (0) | 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Badger-Emeka, L.I.; Kausar, N.; Estrella, E.; Angeles, G.B. A Three-Year Look at the Phylogenetic Profile, Antimicrobial Resistance, and Associated Virulence Genes of Uropathogenic Escherichia coli. Pathogens 2022, 11, 631. https://doi.org/10.3390/pathogens11060631
Badger-Emeka LI, Kausar N, Estrella E, Angeles GB. A Three-Year Look at the Phylogenetic Profile, Antimicrobial Resistance, and Associated Virulence Genes of Uropathogenic Escherichia coli. Pathogens. 2022; 11(6):631. https://doi.org/10.3390/pathogens11060631
Chicago/Turabian StyleBadger-Emeka, Lorina I., Naheed Kausar, Edric Estrella, and Glenda Belgira Angeles. 2022. "A Three-Year Look at the Phylogenetic Profile, Antimicrobial Resistance, and Associated Virulence Genes of Uropathogenic Escherichia coli" Pathogens 11, no. 6: 631. https://doi.org/10.3390/pathogens11060631
APA StyleBadger-Emeka, L. I., Kausar, N., Estrella, E., & Angeles, G. B. (2022). A Three-Year Look at the Phylogenetic Profile, Antimicrobial Resistance, and Associated Virulence Genes of Uropathogenic Escherichia coli. Pathogens, 11(6), 631. https://doi.org/10.3390/pathogens11060631