
Genomic and Biological Profile of a Novel Bacteriophage, Vibrio phage Virtus, Which Improves Survival of Sparus aurata Larvae Challenged with Vibrio harveyi



Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Isolation and Purification of Bacteriophages
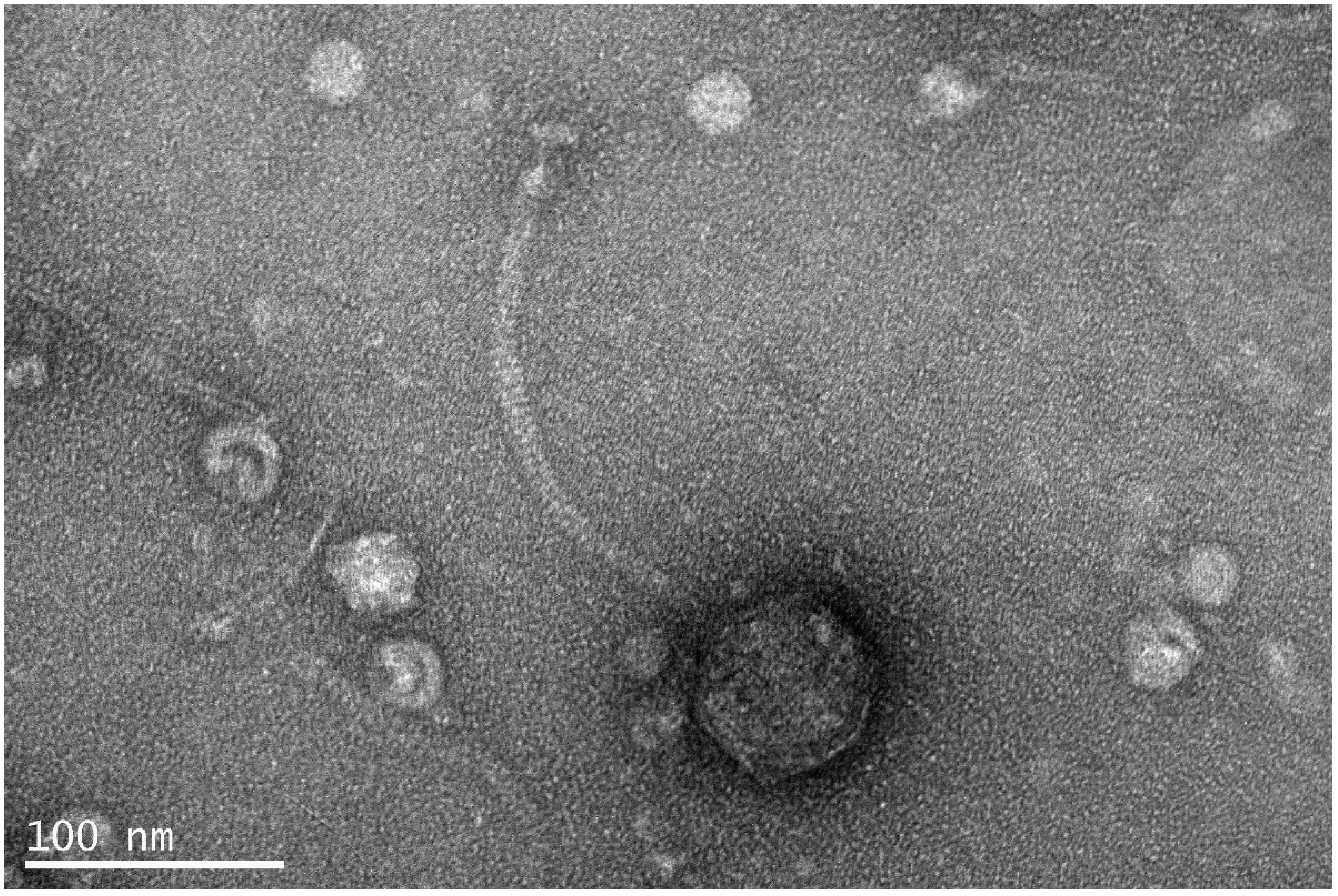
2.3. Transmission Electron Microscopy
2.4. Lysogeny Test
2.5. Host Range Test
2.6. Efficiency of Plating (EOP)
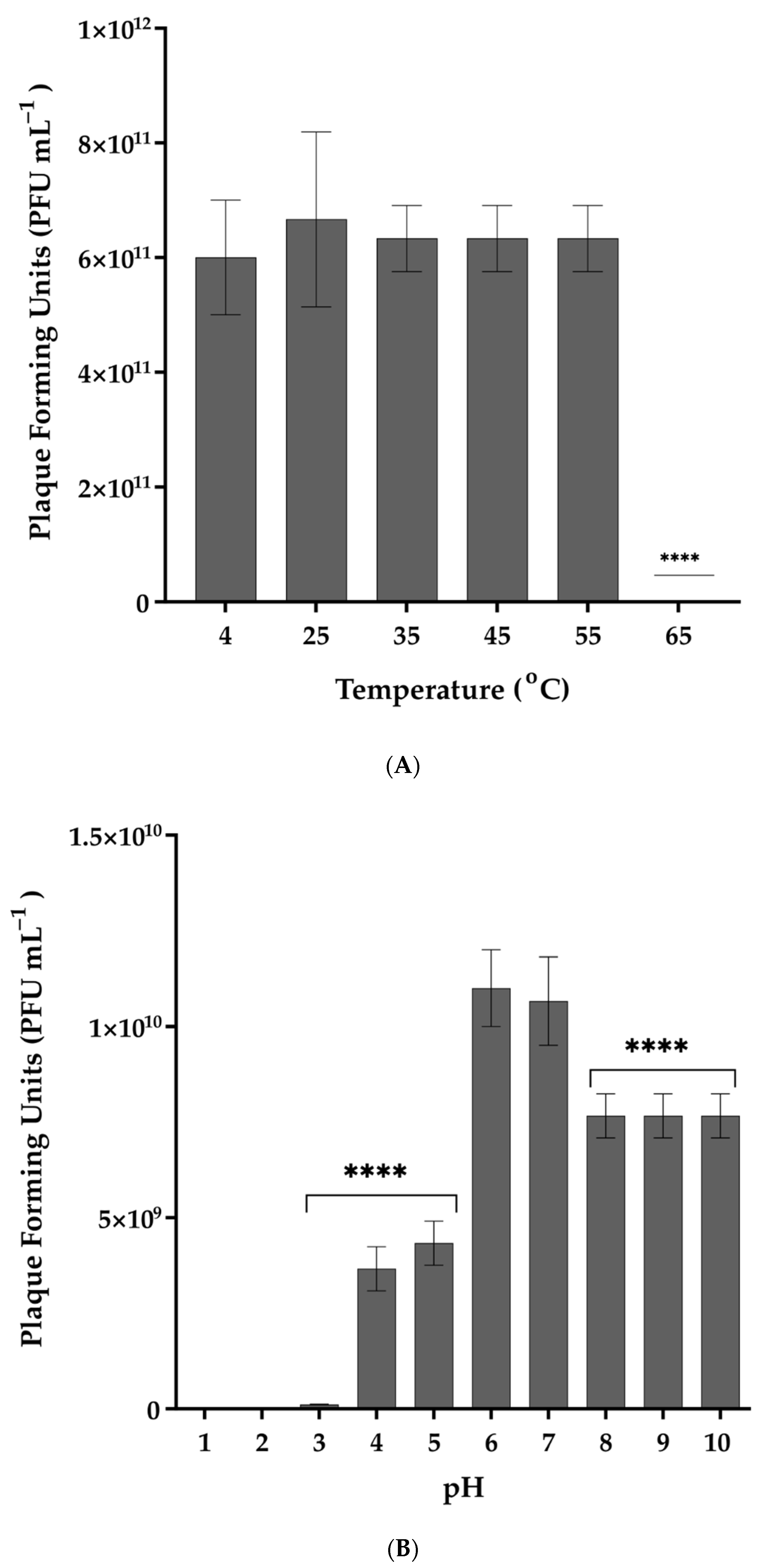
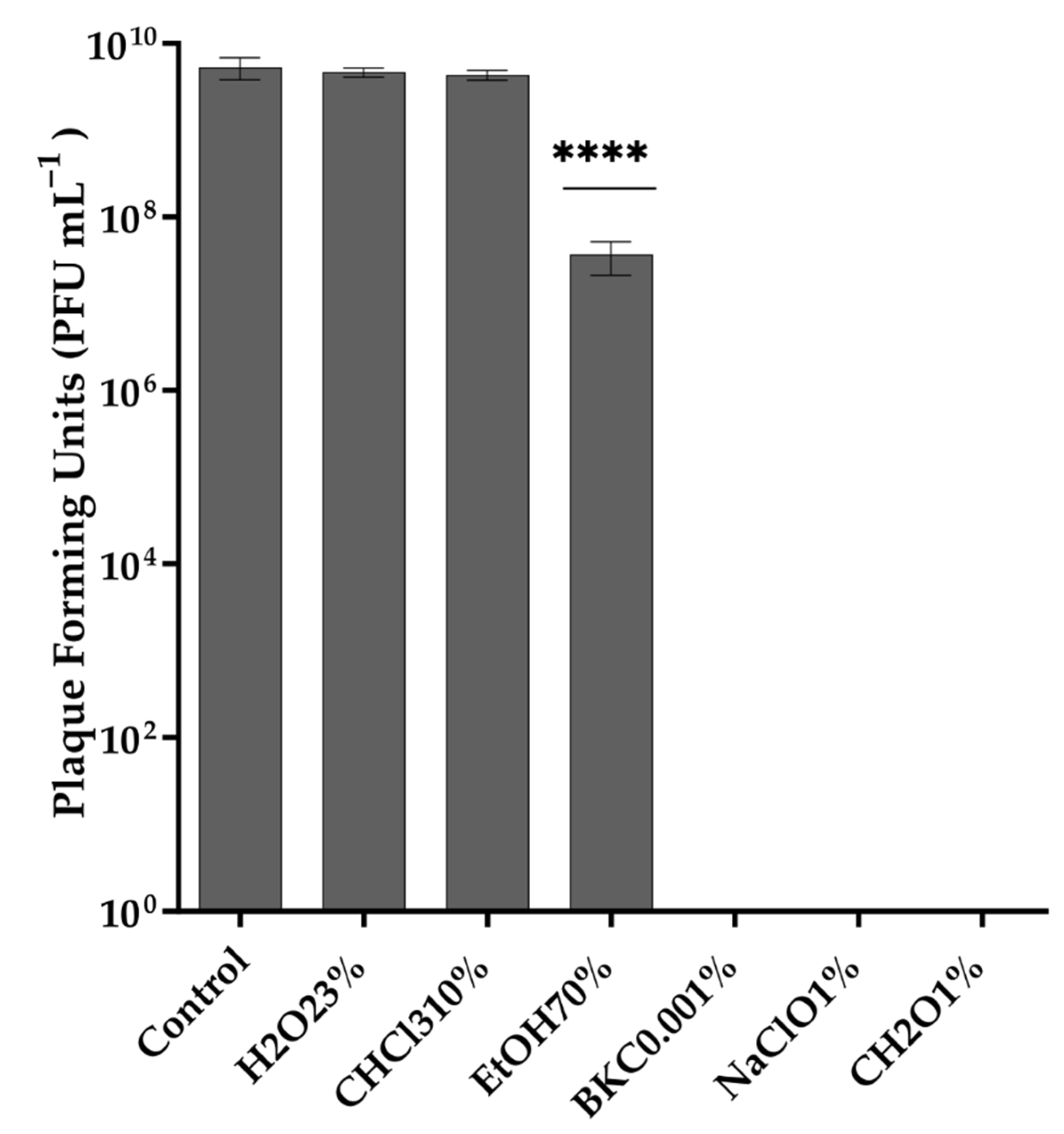
2.7. Stability of Phage in Different Temperatures, pH Values, and Organic Solvents
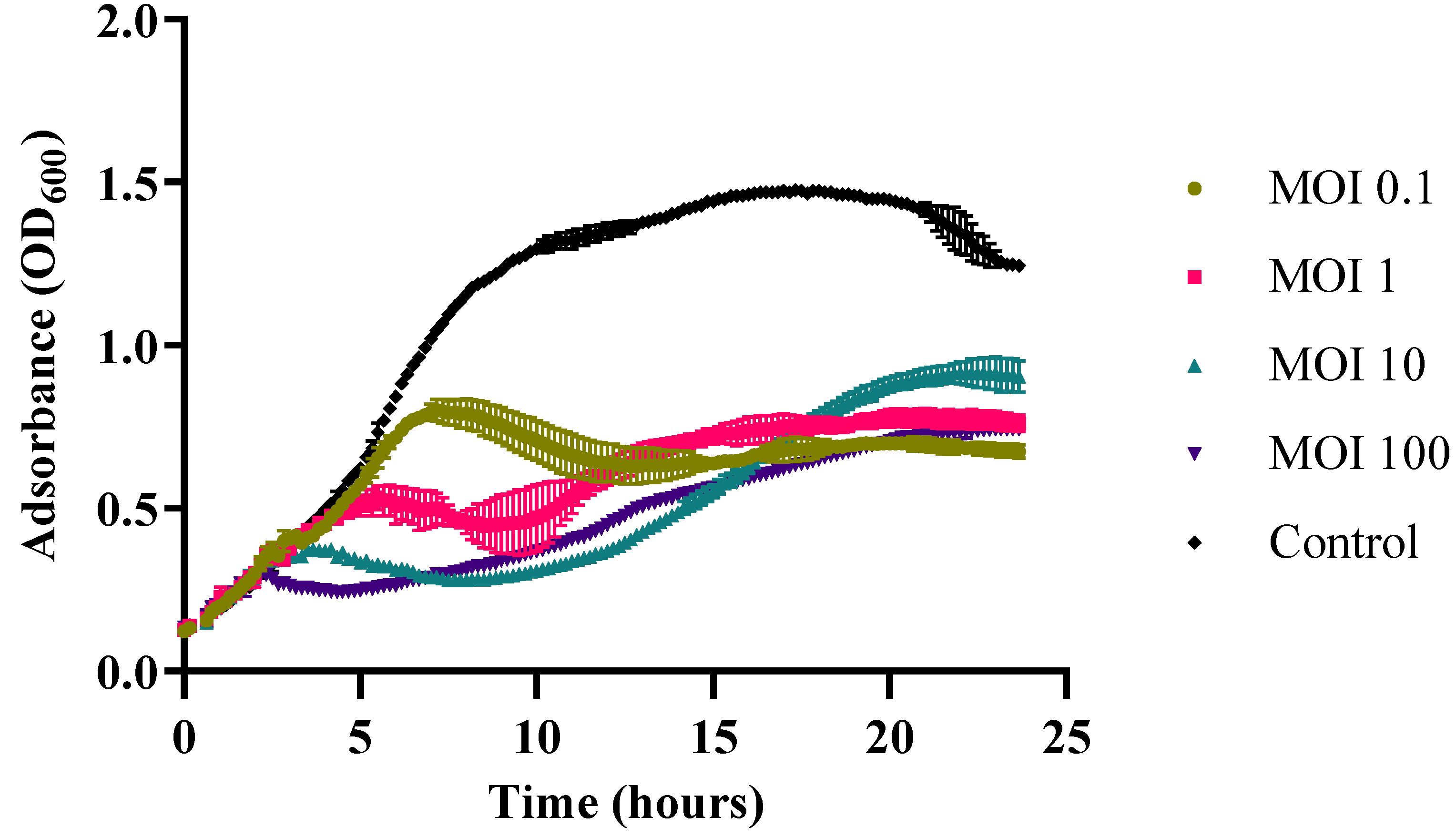
2.8. One-Step Growth
2.9. In Vitro Cell Lysis
2.10. DNA Extraction and Purification
2.11. Genomic Analysis
2.12. Genome Alignment and Phylogenetic Analysis of Vibrio phage Virtus
2.13. In Vivo Phage Therapy Trial in Gilthead Seabream Larvae
2.14. Statistical Analysis
2.15. Data Availability
3. Results
3.1. Isolation and Morphology of Vibrio phage Virtus
3.2. Host Range and Efficiency of Plating (EOP) of Vibrio phage Virtus against Multiple Antibiotic Resistant Strains
3.3. Thermal and pH Stability of Vibrio phage Virtus and Exposure to Organic Solvents and Common Disinfectants
3.4. One-Step Growth of Vibrio phage Virtus
3.5. In Vitro Cell Lysis
3.6. Whole Genome Sequencing and Assembly
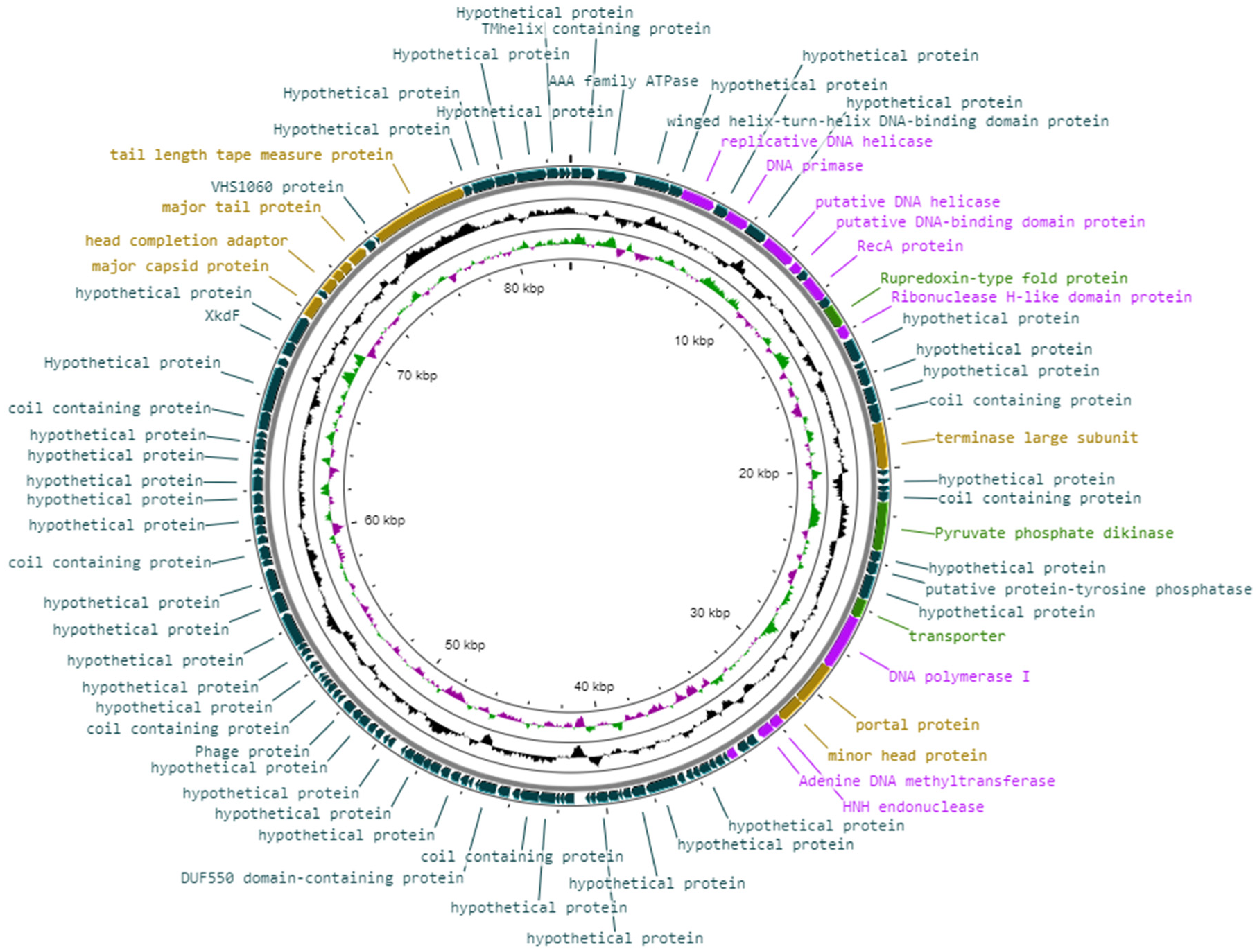
3.7. Genomic Features of Vibrio phage Virtus
3.8. Genomic Arrangement and Functional Annotations of Vibrio phage Virtus
3.8.1. Phage Structural Proteins
3.8.2. DNA Replication, Repair, and Recombination
3.8.3. Miscellaneous Proteins
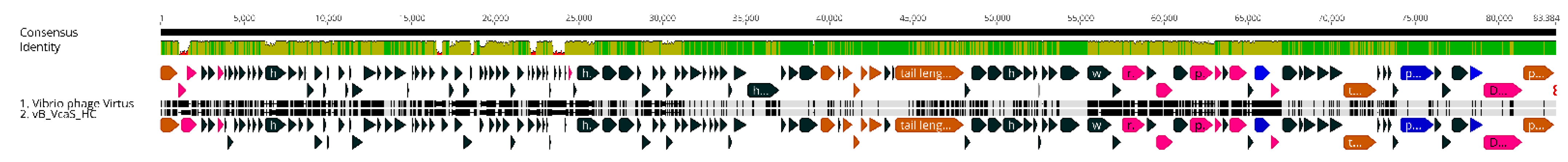
3.9. Genomic Synteny of Vibrio phage Virtus with Other Similar Phages
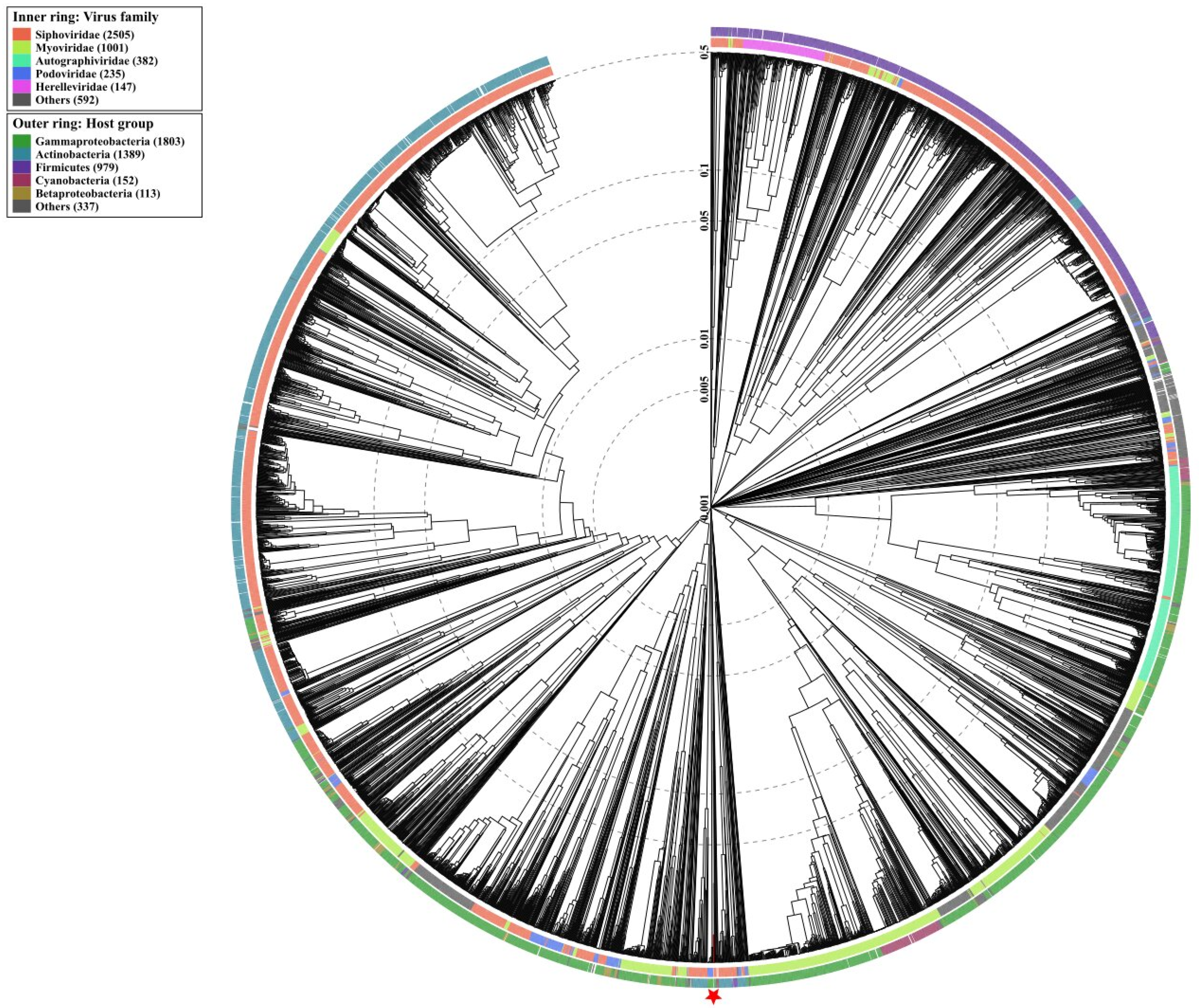
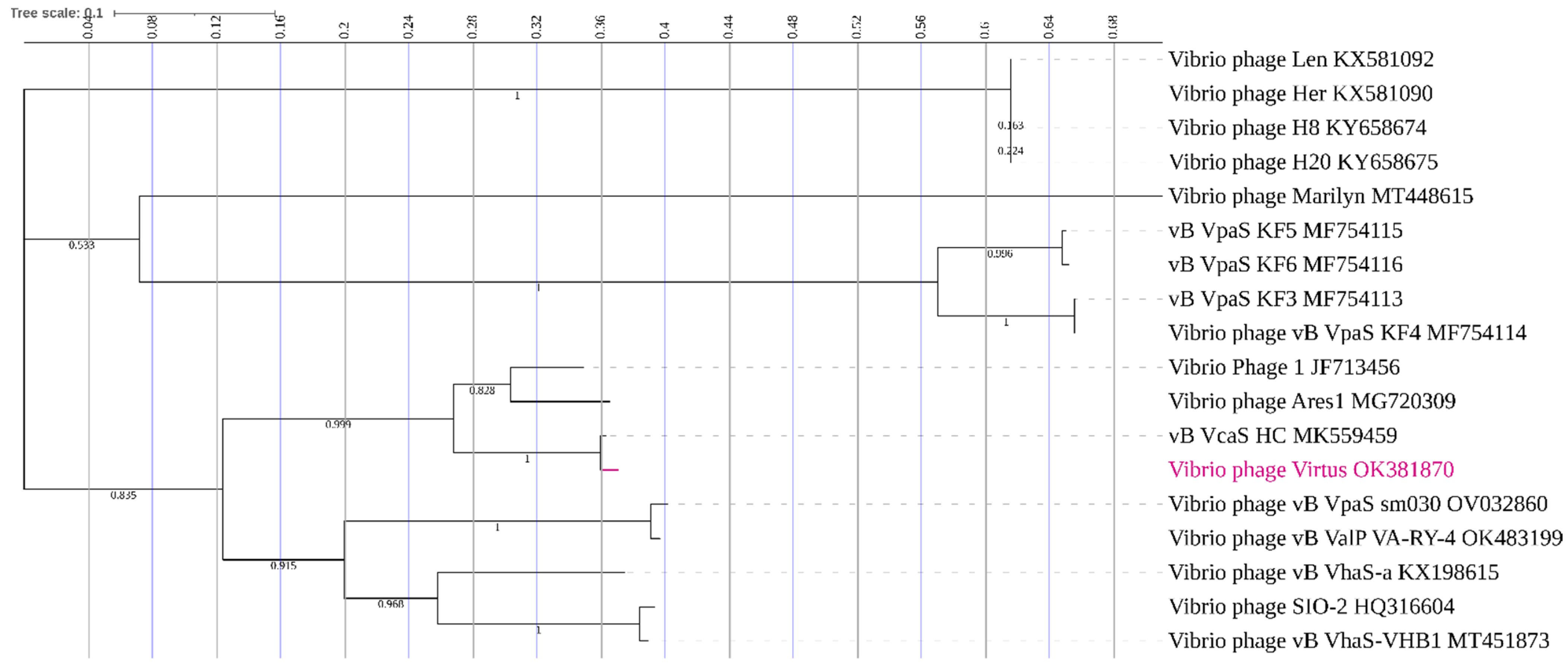
3.10. Phylogenetic Analysis
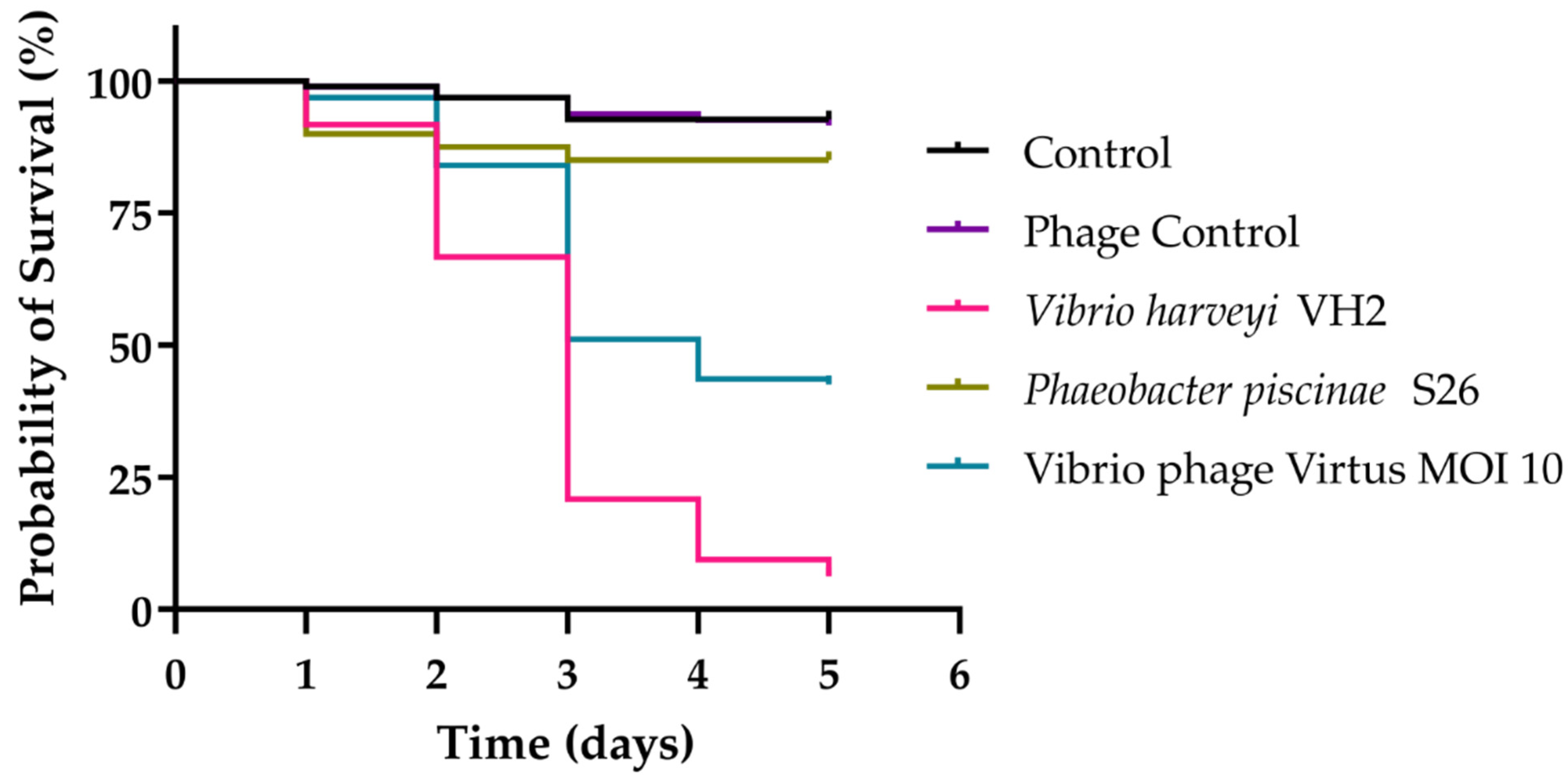
3.11. In Vivo Phage Therapy in Gilthead Seabream Larvae
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, X.H.; He, X.; Austin, B. Vibrio Harveyi: A Serious Pathogen of Fish and Invertebrates in Mariculture. Mar. Life Sci. Technol. 2020, 2, 231–245. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.C.; Chuang, W.H.; Lee, K.K. Infectious Gastroenteritis Caused by Vibrio Harveyi (V. Carchariae) in Cultured Red Drum, Sciaenops Ocellatus. J. Appl. Ichthyol. 2003, 19, 59–61. [Google Scholar] [CrossRef]
- Montánchez, I.; Ogayar, E.; Plágaro, A.H.; Esteve-Codina, A.; Gómez-Garrido, J.; Orruño, M.; Arana, I.; Kaberdin, V.R. Analysis of Vibrio Harveyi Adaptation in Sea Water Microcosms at Elevated Temperature Provides Insights into the Putative Mechanisms of Its Persistence and Spread in the Time of Global Warming. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Vezzulli, L.; Grande, C.; Reid, P.C.; Hélaouët, P.; Edwards, M.; Höfle, M.G.; Brettar, I.; Colwell, R.R.; Pruzzo, C. Climate Influence on Vibrio and Associated Human Diseases during the Past Half-Century in the Coastal North Atlantic. Proc. Natl. Acad. Sci. USA 2016, 113, E5062–E5071. [Google Scholar] [CrossRef]
- Sterk, A.; Schets, F.M.; de Roda Husman, A.M.; de Nijs, T.; Schijven, J.F. Effect of Climate Change on the Concentration and Associated Risks of Vibrio Spp. in Dutch Recreational Waters. Risk Anal. 2015, 35, 1717–1729. [Google Scholar] [CrossRef]
- Montánchez, I.; Kaberdin, V.R. Vibrio Harveyi: A Brief Survey of General Characteristics and Recent Epidemiological Traits Associated with Climate Change. Mar. Environ. Res. 2020, 154, 104850. [Google Scholar] [CrossRef]
- Cascarano, M.C.; Stavrakidis-Zachou, O.; Mladineo, I.; Thompson, K.D.; Papandroulakis, N.; Katharios, P. Mediterranean Aquaculture in a Changing Climate: Temperature Effects on Pathogens and Diseases of Three Farmed Fish Species. Pathogens 2021, 10, 1205. [Google Scholar] [CrossRef]
- Rigos, G.; Kogiannou, D.; Padrós, F.; Cristòfol, C.; Florio, D.; Fioravanti, M.; Zarza, C. Best Therapeutic Practices for the Use of Antibacterial Agents in Finfish Aquaculture: A Particular View on European Seabass (Dicentrarchus Labrax) and Gilthead Seabream (Sparus Aurata) in Mediterranean Aquaculture. Rev. Aquac. 2021, 13, 1285–1323. [Google Scholar] [CrossRef]
- Wee, B.A.; Muloi, D.M.; van Bunnik, B.A.D. Quantifying the Transmission of Antimicrobial Resistance at the Human and Livestock Interface with Genomics. Clin. Microbiol. Infect. 2020, 26, 1612–1616. [Google Scholar] [CrossRef]
- Kalatzis, P.G.; Castillo, D.; Katharios, P.; Middelboe, M. Bacteriophage Interactions with Marine Pathogenic Vibrios: Implications for Phage Therapy. Antibiotics 2018, 7, 15. [Google Scholar] [CrossRef]
- Culot, A.; Grosset, N.; Gautier, M. Overcoming the Challenges of Phage Therapy for Industrial Aquaculture: A Review. Aquaculture 2019, 513, 734423. [Google Scholar] [CrossRef]
- Kalatzis, P.G.; Bastías, R.; Kokkari, C.; Katharios, P. Isolation and Characterization of Two Lytic Bacteriophages, Φst2 and Φgrn1; Phage Therapy Application for Biological Control of Vibrio Alginolyticus in Aquaculture Live Feeds. PLoS ONE 2016, 11, e0151101. [Google Scholar] [CrossRef] [PubMed]
- Silva, Y.J.; Costa, L.; Pereira, C.; Mateus, C.; Cunha, Â.; Calado, R.; Gomes, N.C.M.; Pardo, M.A.; Hernandez, I.; Almeida, A. Phage Therapy as an Approach to Prevent Vibrio Anguillarum Infections in Fish Larvae Production. PLoS ONE 2014, 9, 1–23. [Google Scholar] [CrossRef]
- Martínez-Díaz, S.F.; Hipólito-Morales, A. Efficacy of Phage Therapy to Prevent Mortality during the Vibriosis of Brine Shrimp. Aquaculture 2013, 400–401, 120–124. [Google Scholar] [CrossRef]
- Castillo, D.; D’Alvise, P.; Middelboe, M.; Gram, L.; Liu, S.; Kalatzis, P.G.; Kokkari, C.; Katharios, P. Draft Genome Sequences of the Fish Pathogen Vibrio Harveyi Strains VH2 and VH5. Genome Announc. 2015, 3, 2–3. [Google Scholar] [CrossRef] [PubMed]
- Clokie, M.R.J.; Kropinski, A.M. Bacteriophages: Volume I; Humana: Totowa, NJ, USA, 2009; Volume 1, ISBN 9781627032384. [Google Scholar]
- Misol, G.N.; Kokkari, C.; Katharios, P. Biological and Genomic Characterization of a Novel Jumbo Bacteriophage, Vb_VhaM_pir03 with Broad Host Lytic Activity against Vibrio Harveyi. Pathogens 2020, 9, 1051. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Denes, T.; Den Bakker, H.C.; Tokman, J.I.; Guldimann, C.; Wiedmann, M. Selection and Characterization of Phage-Resistant Mutant Strains of Listeria Monocytogenes Reveal Host Genes Linked to Phage Adsorption. Appl. Environ. Microbiol. 2015, 81, 4295–4305. [Google Scholar] [CrossRef]
- Jäckel, C.; Hertwig, S.; Scholz, H.C.; Nöckler, K.; Reetz, J.; Hammerl, J.A. Prevalence, Host Range, and Comparative Genomic Analysis of Temperate Ochrobactrum Phages. Front. Microbiol. 2017, 8, 1207. [Google Scholar] [CrossRef]
- Kutter, E. Chapter 14—Phage Host Range and Efficiency of Plating. In Bacteriophages: Methods and Protocols, Volume 1: Isolation, Characterization, and Interactions; Humana: Totowa, NJ, USA, 2009; Volume 501, pp. 141–149. [Google Scholar] [CrossRef]
- Pan, L.; Li, D.; Sun, Z.; Lin, W.; Hong, B.; Qin, W.; Xu, L.; Liu, W.; Zhou, Q.; Wang, F.; et al. First Characterization of a Hafnia Phage Reveals Extraordinarily Large Burst Size and Unusual Plaque Polymorphism. Front. Microbiol. 2022, 12, 754331. [Google Scholar] [CrossRef]
- Higuera, G.; Bastías, R.; Tsertsvadze, G.; Romero, J.; Espejo, R.T. Recently Discovered Vibrio Anguillarum Phages Can Protect against Experimentally Induced Vibriosis in Atlantic Salmon, Salmo Salar. Aquaculture 2013, 392–395, 128–133. [Google Scholar] [CrossRef]
- Davis, J.J.; Wattam, A.R.; Aziz, R.K.; Brettin, T.; Butler, R.; Butler, R.M.; Chlenski, P.; Conrad, N.; Dickerman, A.; Dietrich, E.M.; et al. The PATRIC Bioinformatics Resource Center: Expanding Data and Analysis Capabilities. Nucleic Acids Res. 2020, 48, D606–D612. [Google Scholar] [CrossRef] [PubMed]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality Assessment Tool for Genome Assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef] [PubMed]
- Bushnell, B. BBMap: A Fast, Accurate, Splice-Aware Aligner; Lawrence Berkeley National Lab. (LBNL): Berkeley, CA, USA, 2014. [Google Scholar]
- Garneau, J.R.; Depardieu, F.; Fortier, L.C.; Bikard, D.; Monot, M. PhageTerm: A Tool for Fast and Accurate Determination of Phage Termini and Packaging Mechanism Using next-Generation Sequencing Data. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ramsey, J.; Rasche, H.; Maughmer, C.; Criscione, A.; Mijalis, E.; Liu, M.; Hu, J.C.; Young, R.; Gill, J.J. Galaxy and Apollo as a Biologist-Friendly Interface for High-Quality Cooperative Phage Genome Annotation. PLoS Comput. Biol. 2020, 16, e1008214. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Harris, M.A.; Clark, J.; Ireland, A.; Lomax, J.; Ashburner, M.; Foulger, R.; Eilbeck, K.; Lewis, S.; Marshall, B.; Mungall, C.; et al. The Gene Oncology (GO) Database and Informatics Resource. Nucleic Acids Res. 2004, 32, 258–261. [Google Scholar] [CrossRef]
- Mitchell, A.L.; Attwood, T.K.; Babbitt, P.C.; Blum, M.; Bork, P.; Bridge, A.; Brown, S.D.; Chang, H.Y.; El-Gebali, S.; Fraser, M.I.; et al. InterPro in 2019: Improving Coverage, Classification and Access to Protein Sequence Annotations. Nucleic Acids Res. 2019, 47, D351–D360. [Google Scholar] [CrossRef]
- Krogh, A.; Larsson, B.; Von Heijne, G.; Sonnhammer, E.L.L. Predicting Transmembrane Protein Topology with a Hidden Markov Model: Application to Complete Genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef]
- Sonnhammer, E.L.L.; Krogh, A. A Hidden Markov Model for Predicting Transmembrane Helices in Protein Sequence. In Proceedings of the Sixth International Conference on Intelligent Systems for Molecular Biology, Toronto, ON, Canada, 19–23 July 2008; Volume 8, pp. 175–182. [Google Scholar]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The Conserved Domain Database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef]
- Moura, A.; Soares, M.; Pereira, C.; Leitão, N.; Henriques, I.; Correia, A. INTEGRALL: A Database and Search Engine for Integrons, Integrases and Gene Cassettes. Bioinformatics 2009, 25, 1096–1098. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Yang, J.; Yu, J.; Yao, Z.; Sun, L.; Shen, Y.; Jin, Q. VFDB: A Reference Database for Bacterial Virulence Factors. Nucleic Acids Res. 2005, 33, 325–328. [Google Scholar] [CrossRef] [PubMed]
- Kleinheinz, K.A.; Joensen, K.G.; Larsen, M.V. Applying the ResFinder and VirulenceFinder Web-Services for Easy Identification of Acquired Antibiotic Resistance and E. Coli Virulence Genes in Bacteriophage and Prophage Nucleotide Sequences. Bacteriophage 2014, 4, e27943. [Google Scholar] [CrossRef] [PubMed]
- Arndt, D.; Grant, J.R.; Marcu, A.; Sajed, T.; Pon, A.; Liang, Y.; Wishart, D.S. PHASTER: A Better, Faster Version of the PHAST Phage Search Tool. Nucleic Acids Res. 2016, 44, W16–W21. [Google Scholar] [CrossRef]
- Hockenberry, A.J.; Wilke, C.O. BACPHLIP: Predicting Bacteriophage Lifestyle from Conserved Protein Domains. PeerJ 2021, 9, e11396. [Google Scholar] [CrossRef]
- Darling, A.E.; Mau, B.; Perna, N.T. Progressivemauve: Multiple Genome Alignment with Gene Gain, Loss and Rearrangement. PLoS ONE 2010, 5, e11147. [Google Scholar] [CrossRef]
- Needleman, S.B.; Wunsch, C.D. A General Method Applicable to the Search for Similarities in the Amino Acid Sequence of Two Proteins. J. Mol. Biol. 1970, 48, 443–453. [Google Scholar] [CrossRef]
- Smith, T.F.; Waterman, M.S. Identification of Common Molecular Subsequences. J. Mol. Biol. 1981, 147, 195–197. [Google Scholar] [CrossRef]
- Nishimura, Y.; Yoshida, T.; Kuronishi, M.; Uehara, H.; Ogata, H.; Goto, S. ViPTree: The Viral Proteomic Tree Server. Bioinformatics 2017, 33, 2379–2380. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: A Multiple Sequence Alignment Method with Reduced Time and Space Complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The Rapid Generation of Mutation Data Matrices. Bioinformatics 1992, 8, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree of Life (ITOL) v5: An Online Tool for Phylogenetic Tree Display and Annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Panini, E.B.; Mylonas, C.C.; Zanuy, S.; Carrillo, M.; Ramos, J.; Bruce, M.P. Incubation of Embryos and Larvae of Marine Fish Using Microtiter Plates. Aquac. Int. 2001, 9, 189–196. [Google Scholar] [CrossRef]
- Dunnett, C.W. A Multiple Comparison Procedure for Comparing Several Treatments with a Control. J. Am. Stat. Assoc. 1955, 50, 1096–1121. [Google Scholar] [CrossRef]
- Haynes, W. Tukey’s Test. In Encyclopedia of Systems Biology; Dubitzky, W., Wolkenhauer, O., Cho, K.-H., Yokota, H., Eds.; Springer: New York, NY, USA, 2013; pp. 2303–2304. ISBN 978-1-4419-9863-7. [Google Scholar]
- Kishore, J.; Goel, M.; Khanna, P. Understanding Survival Analysis: Kaplan-Meier Estimate. Int. J. Ayurveda Res. 2010, 1, 274. [Google Scholar] [CrossRef]
- Cardinaud, M.; Barbou, A.; Capitaine, C.; Bidault, A.; Dujon, A.M.; Moraga, D.; Paillard, C. Vibrio Harveyi Adheres to and Penetrates Tissues of the European Abalone Haliotis Tuberculata within the First Hours of Contact. Appl. Environ. Microbiol. 2014, 80, 6328–6333. [Google Scholar] [CrossRef]
- Martin, G.G.; Rubin, N.; Swanson, E. Vibrio Parahaemolyticus and V. Harveyi Cause Detachment of the Epithelium from the Midgut Trunk of the Penaeid Shrimp Sicyonia Ingentis. Dis. Aquat. Organ. 2004, 60, 21–29. [Google Scholar] [CrossRef]
- Haldar, S.; Maharajan, A.; Chatterjee, S.; Hunter, S.A.; Chowdhury, N.; Hinenoya, A.; Asakura, M.; Yamasaki, S. Identification of Vibrio Harveyi as a Causative Bacterium for a Tail Rot Disease of Sea Bream Sparus Aurata from Research Hatchery in Malta. Microbiol. Res. 2010, 165, 639–648. [Google Scholar] [CrossRef]
- Luna, G.M.; Bongiorni, L.; Gili, C.; Biavasco, F.; Danovaro, R. Vibrio Harveyi as a Causative Agent of the White Syndrome in Tropical Stony Corals. Environ. Microbiol. Rep. 2010, 2, 120–127. [Google Scholar] [CrossRef]
- Mohd Yazid, S.H.; Mohd Daud, H.; Azmai, M.N.A.; Mohamad, N.; Mohd Nor, N. Estimating the Economic Loss Due to Vibriosis in Net-Cage Cultured Asian Seabass (Lates Calcarifer): Evidence From the East Coast of Peninsular Malaysia. Front. Vet. Sci. 2021, 8, 644009. [Google Scholar] [CrossRef] [PubMed]
- Novriadi, R. Vibriosis in Aquaculture. Omni-Akuatika 2016, 12, 4–5. [Google Scholar] [CrossRef]
- Deng, Y.; Xu, L.; Liu, S.; Wang, Q.; Guo, Z.; Chen, C.; Feng, J. What Drives Changes in the Virulence and Antibiotic Resistance of Vibrio Harveyi in the South China Sea? J. Fish Dis. 2020, 43, 853–862. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, T.; Ito, E.; Nomura, N.; Nomura, N.; Matsumura, M. Comparison of Vibrio Harveyi Strains Isolated from Shrimp Farms and from Culture Collection in Terms of Toxicity and Antibiotic Resistance. FEMS Microbiol. Lett. 2006, 258, 194–199. [Google Scholar] [CrossRef][Green Version]
- Karunasagar, I.; Pai, R.; Malathi, G.R.; Karunasagar, I. Mass Mortality of Penaeus Monodon Larvae Due to Antibiotic-Resistant Vibrio Harveyi Infection. Aquaculture 1994, 128, 203–209. [Google Scholar] [CrossRef]
- Vinod, M.G.; Shivu, M.M.; Umesha, K.R.; Rajeeva, B.C.; Krohne, G.; Karunasagar, I.; Karunasagar, I. Isolation of Vibrio Harveyi Bacteriophage with a Potential for Biocontrol of Luminous Vibriosis in Hatchery Environments. Aquaculture 2006, 255, 117–124. [Google Scholar] [CrossRef]
- Shivu, M.M.; Rajeeva, B.C.; Girisha, S.K.; Karunasagar, I.; Krohne, G.; Karunasagar, I. Molecular Characterization of Vibrio Harveyi Bacteriophages Isolated from Aquaculture Environments along the Coast of India. Environ. Microbiol. 2007, 9, 322–331. [Google Scholar] [CrossRef]
- Wu, L.; Tian, Y.; Pang, M.; Yang, Z.; Bao, H.; Zhou, Y.; Sun, L.; Wang, R.; Zhang, H. A Novel Vibriophage VB_VhaS_PcB-1G Capable of Inhibiting Virulent Vibrio Harveyi Pathogen. Aquaculture 2021, 542, 736854. [Google Scholar] [CrossRef]
- Nurhafizah, W.W.I.; Alia, S.A.; Azna, N.S.; Lee, K.L.; Nadirah, M.; Laith, A.R.; Danish-Daniel, M.; Zainathan, S.C.; Shariff, M.M.D.; Mazlan, A.G.; et al. In-Vitro Characterization of Lytic Bacteriophage PhVh6 as Potential Biocontrol Agent against Pathogenic Vibrio Harveyi. AACL Bioflux 2017, 10, 64–76. [Google Scholar]
- Baudoux, A.-C.; Hendrix, R.W.; Lander, G.C.; Bailly, X.; Podell, S.; Paillard, C.; Johnson, J.E.; Potter, C.S.; Carragher, B.; Azam, F. Genomic and Functional Analysis of Vibrio Phage SIO-2 Reveals Novel Insights into Ecology and Evolution of Marine Siphoviruses. Environ. Microbiol. 2012, 14, 2071–2086. [Google Scholar] [CrossRef]
- Pasharawipas, T.; Thaikua, S.; Sriurairatana, S.; Ruangpan, L.; Direkbusarakum, S.; Manopvisetcharean, J.; Flegel, T.W. Partial Characterization of a Novel Bacteriophage of Vibrio Harveyi Isolated from Shrimp Culture Ponds in Thailand. Virus Res. 2005, 114, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Lal, T.M.; Sano, M.; Ransangan, J. Isolation and Characterization of Large Marine Bacteriophage (Myoviridae), VhKM4 Infecting Vibrio Harveyi. J. Aquat. Anim. Health 2017, 29, 26–30. [Google Scholar] [CrossRef] [PubMed]
- Oakey, H.J.; Owens, L. A New Bacteriophage, VHML, Isolated from a Toxin-Producing Strain of Vibrio Harveyi in Tropical Australia. J. Appl. Microbiol. 2000, 89, 702–709. [Google Scholar] [CrossRef] [PubMed]
- Thiyagarajan, S.; Chrisolite, B.; Alavandi, S.V.; Poornima, M.; Kalaimani, N.; Santiago, T.C. Characterization of Four Lytic Transducing Bacteriophages of Luminescent Vibrio Harveyi Isolated from Shrimp (Penaeus Monodon) Hatcheries. FEMS Microbiol. Lett. 2011, 325, 85–91. [Google Scholar] [CrossRef][Green Version]
- Turner, D.; Kropinski, A.M.; Adriaenssens, E.M. A Roadmap for Genome-Based Phage Taxonomy. Viruses 2021, 13, 506. [Google Scholar] [CrossRef]
- Kerr, B.; West, J.; Bohannan, B.J.M. Bacteriophages: Models for Exploring Basic Principles of Ecology; Cambridge University Press: Cambridge, UK, 2009; ISBN 9780511541483. [Google Scholar]
- Chow, C.E.T.; Suttle, C.A. Biogeography of Viruses in the Sea. Annu. Rev. Virol. 2015, 2, 41–66. [Google Scholar] [CrossRef]
- Weinbauer, M.G. Ecology of Prokaryotic Viruses. FEMS Microbiol. Rev. 2004, 28, 127–181. [Google Scholar] [CrossRef]
- Holmfeldt, K.; Middelboe, M.; Nybroe, O.; Riemann, L. Large Variabilities in Host Strain Susceptibility and Phage Host Range Govern Interactions between Lytic Marine Phages and Their Flavobacterium Hosts. Appl. Environ. Microbiol. 2007, 73, 6730–6739. [Google Scholar] [CrossRef]
- De Jonge, P.A.; Nobrega, F.L.; Brouns, S.J.J.; Dutilh, B.E. Molecular and Evolutionary Determinants of Bacteriophage Host Range. Trends Microbiol. 2019, 27, 51–63. [Google Scholar] [CrossRef]
- Thompson, F.L.; Iida, T.; Swings, J. Biodiversity of Vibrios. Microbiol. Mol. Biol. Rev. 2004, 68, 403–431. [Google Scholar] [CrossRef]
- Baker-Austin, C.; Oliver, J.D.; Alam, M.; Ali, A.; Waldor, M.K.; Qadri, F.; Martinez-Urtaza, J. Vibrio Spp. Infections. Nat. Rev. Dis. Prim. 2018, 4, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.; Huang, Y.; Mi, Z.; Yin, X.; Wang, L.; Fan, H.; Zhang, Z.; An, X.; Chen, J.; Tong, Y. Complete Genome Sequence of IME13, a Stenotrophomonas Maltophilia Bacteriophage with Large Burst Size and Unique Plaque Polymorphism. J. Virol. 2012, 86, 11392–11393. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zeng, H.; Li, C.; Luo, D.; Zhang, J.; Ding, Y.; Chen, M.; Yang, X.; Lei, T.; Wu, S.; Ye, Q.; et al. Novel Phage VB_CtuP_B1 for Controlling Cronobacter Malonaticus and Cronobacter Turicensis in Ready-to-Eat Lettuce and Powered Infant Formula. Food Res. Int. 2021, 143, 110255. [Google Scholar] [CrossRef] [PubMed]
- Endy, D.; Kong, D.; Yin, J. Intracellular Kinetics of a Growing Virus: A Genetically Structured Simulation for Bacteriophage T7. Biotechnol. Bioeng. 1997, 55, 375–389. [Google Scholar] [CrossRef]
- Abedon, S.T.; Herschler, T.D.; Stopar, D. Bacteriophage Latent-Period Evolution as a Response to Resource Availability. Appl. Environ. Microbiol. 2001, 67, 4233–4241. [Google Scholar] [CrossRef]
- Sui, B.; Qi, X.; Wang, X.; Ren, H.; Liu, W.; Zhang, C. Characterization of a Novel Bacteriophage Swi2 Harboring Two Lysins Can Naturally Lyse Escherichia Coli. Front. Microbiol. 2021, 12, 670799. [Google Scholar] [CrossRef]
- Hyman, P.; Abedon, S.T. Practical Methods for Determining Phage Growth Parameters. In BT-Bacteriophages: Methods and Protocols, Volume 1: Isolation, Characterization, and Interactions; Clokie, M.R.J., Kropinski, A.M., Eds.; Humana Press: Totowa, NJ, USA, 2009; pp. 175–202. ISBN 978-1-60327-164-6. [Google Scholar]
- Villa, T.G.; Feijoo-Siota, L.; Rama, J.L.R.; Sánchez-Pérez, A.; Viñas, M. Horizontal Gene Transfer Between Bacteriophages and Bacteria: Antibiotic Resistances and Toxin Production. In Horizontal Gene Transfer: Breaking Borders Between Living Kingdoms; Villa, T.G., Viñas, M., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 97–142. ISBN 978-3-030-21862-1. [Google Scholar]
- Canchaya, C.; Fournous, G.; Chibani-Chennoufi, S.; Dillmann, M.L.; Brüssow, H. Phage as Agents of Lateral Gene Transfer. Curr. Opin. Microbiol. 2003, 6, 417–424. [Google Scholar] [CrossRef]
- Khemayan, K.; Prachumwat, A.; Sonthayanon, B.; Intaraprasong, A.; Sriurairatana, S.; Flegel, T.W. Complete Genome Sequence of Virulence-Enhancing Siphophage VHS1 from Vibrio Harveyi. Appl. Environ. Microbiol. 2012, 78, 2790–2796. [Google Scholar] [CrossRef]
- Li, C.; Wang, Z.; Zhao, J.; Wang, L.; Xie, G.; Huang, J.; Zhang, Y. A Novel Vibriophage VB_VcaS_HC Containing Lysogeny-Related Gene Has Strong Lytic Ability against Pathogenic Bacteria. Virol. Sin. 2021, 36, 281–290. [Google Scholar] [CrossRef]
- Oliveira, L.; Tavares, P.; Alonso, J.C. Headful DNA Packaging: Bacteriophage SPP1 as a Model System. Virus Res. 2013, 173, 247–259. [Google Scholar] [CrossRef]
- Prasad Bhattacharyya, S.; Basaveswara Rao, V. A Novel Terminase Activity Associated with the DNA Packaging Protein Gp17 of Bacteriophage T4. Virology 1993, 196, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.F. Microbial Metabolism. Ref. Modul. Biomed. Sci. 2022, 1, 363–376. [Google Scholar] [CrossRef]
- Warwick-Dugdale, J.; Buchholz, H.H.; Allen, M.J.; Temperton, B. Host-Hijacking and Planktonic Piracy: How Phages Command the Microbial High Seas. Virol. J. 2019, 16, 15. [Google Scholar] [CrossRef] [PubMed]
- Miller, E.S.; Heidelberg, J.F.; Eisen, J.A.; Nelson, W.C.; Durkin, A.S.; Ciecko, A.; Feldblyum, T.V.; White, O.; Paulsen, I.T.; Nierman, W.C.; et al. Complete Genome Sequence of the Broad-Host-Range Vibriophage KVP40: Comparative Genomics of a T4-Related Bacteriophage. J. Bacteriol. 2003, 185, 5220–5233. [Google Scholar] [CrossRef] [PubMed]
- Kelly, L.; Ding, H.; Huang, K.H.; Osburne, M.S.; Chisholm, S.W. Genetic Diversity in Cultured and Wild Marine Cyanomyoviruses Reveals Phosphorus Stress as a Strong Selective Agent. ISME J. 2013, 7, 1827–1841. [Google Scholar] [CrossRef] [PubMed]
- Katharios, P.; Kalatzis, P.G.; Kokkari, C.; Sarropoulou, E.; Middelboe, M. Isolation and Characterization of a N4-like Lytic Bacteriophage Infecting Vibrio Splendidus, a Pathogen of Fish and Bivalves. PLoS ONE 2017, 12, e0190083. [Google Scholar] [CrossRef]
- Hellweger, F.L. Carrying Photosynthesis Genes Increases Ecological Fitness of Cyanophage in Silico. Environ. Microbiol. 2009, 11, 1386–1394. [Google Scholar] [CrossRef]
- Middelboe, M.; Hagström, A.; Blackburn, N.; Sinn, B.; Fischer, U.; Borch, N.H.; Pinhassi, J.; Simu, K.; Lorenz, M.G. Effects of Bacteriophages on the Population Dynamics of Four Strains of Pelagic Marine Bacteria. Microb. Ecol. 2001, 42, 395–406. [Google Scholar] [CrossRef]
- Oechslin, F. Resistance Development to Bacteriophages Occurring during Bacteriophage Therapy. Viruses 2018, 10, 351. [Google Scholar] [CrossRef]
- Labrie, S.J.; Samson, J.E.; Moineau, S. Bacteriophage Resistance Mechanisms. Nat. Rev. Microbiol. 2010, 8, 317–327. [Google Scholar] [CrossRef]
- Cui, H.; Cong, C.; Wang, L.; Li, X.; Li, J.; Yang, H.; Li, S.; Xu, Y. Protective Effectiveness of Feeding Phage Cocktails in Controlling Vibrio Harveyi Infection of Turbot Scophthalmus Maximus. Aquaculture 2021, 535, 736390. [Google Scholar] [CrossRef]
- Quiroz-Guzmán, E.; Peña-Rodriguez, A.; Vázquez-Juárez, R.; Barajas-Sandoval, D.R.; Balcázar, J.L.; Martínez-Díaz, S.F. Bacteriophage Cocktails as an Environmentally-Friendly Approach to Prevent Vibrio Parahaemolyticus and Vibrio Harveyi Infections in Brine Shrimp (Artemia Franciscana) Production. Aquaculture 2018, 492, 273–279. [Google Scholar] [CrossRef]
- Yang, Y.; Shen, W.; Zhong, Q.; Chen, Q.; He, X.; Baker, J.L.; Xiong, K.; Jin, X.; Wang, J.; Hu, F.; et al. Development of a Bacteriophage Cocktail to Constrain the Emergence of Phage-Resistant Pseudomonas Aeruginosa. Front. Microbiol. 2020, 11, 327. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Barton, M.; Elliott, L.; Li, X.; Abraham, S.; O’Dea, M.; Munro, J. Bacteriophage Therapy for the Control of Vibrio Harveyi in Greenlip Abalone (Haliotis Laevigata). Aquaculture 2017, 473, 251–258. [Google Scholar] [CrossRef]
- Levin, B.R.; Bull, J.J. Population and Evolutionary Dynamics of Phage Therapy. Nat. Rev. Microbiol. 2004, 2, 166–173. [Google Scholar] [CrossRef]
- Weber, B.; Chen, C.; Milton, D.L. Colonization of Fish Skin Is Vital for Vibrio Anguillarum to Cause Disease. Environ. Microbiol. Rep. 2010, 2, 133–139. [Google Scholar] [CrossRef]
- Hooton, S.P.T.; Brathwaite, K.J.; Connerton, I.F. The Bacteriophage Carrier State of Campylobacter Jejuni Features Changes in Host Non-Coding RNAs and the Acquisition of New Host-Derived CRISPR Spacer Sequences. Front. Microbiol. 2016, 7, 355. [Google Scholar] [CrossRef]
- Mangalea, M.R.; Duerkop, B.A. Fitness Trade-Offs Resulting from Bacteriophage Resistance Potentiate Synergistic Antibacterial Strategies. Infect. Immun. 2020, 88, e00926-19. [Google Scholar] [CrossRef]
- León, M.; Kokkari, C.; García, K.; Castillo, D.; Katharios, P.; Bastías, R. Diversification of Vibrio Anguillarum Driven by the Bacteriophage CHOED. Front. Microbiol. 2019, 10, 1396. [Google Scholar] [CrossRef]
- Pourcel, C.; Midoux, C.; Vergnaud, G.; Latino, L. A Carrier State Is Established in Pseudomonas Aeruginosa by Phage LeviOr01, a Newly Isolated SsRNA Levivirus. J. Gen. Virol. 2017, 98, 2181–2189. [Google Scholar] [CrossRef]
- Manchado, M.; Planas, J.V.; Cousin, X.; Rebordinos, L.; Claros, M.G. Current Status in Other Finfish Species: Description of Current Genomic Resources for the Gilthead Seabream (Sparus Aurata) and Soles (Solea Senegalensis and Solea Solea). Genom. Aquac. 2016, 195–221. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain ID | Species | Country | Location | Date | Host |
|---|---|---|---|---|---|
| DSM 19623 | V. harveyi | USA | Massachusetts | - | Talochestria capensis |
| SA 2.1 | V. harveyi | Saudi Arabia | Red Sea | July 2019 | Sparus aurata |
| DSM 2171 | V. alginolyticus | Japan | - | - | Trachurus trachurus |
| Gal 90 | V. harveyi | Greece | Central Greece | September 2020 | Sparus aurata |
| Vh No22 | V. harveyi | Greece | Ionian Islands | September 2015 | Dichentrachus labrax |
| Kef 62 | V. harveyi | Greece | Ionian Islands | July 2020 | Dichentrachus labrax |
| Kef 75 | V. harveyi | Greece | Ionian Islands | July 2020 | Dichentrachus labrax |
| Gal 56 | V. harveyi | Greece | Central Greece | May 2020 | Dichentrachus labrax |
| Gal 77 | V. harveyi | Greece | Central Greece | July 2020 | Sparus aurata |
| Gal 72 | V. harveyi | Greece | Central Greece | June 2020 | Dichentrachus labrax |
| Gal 94 | V. harveyi | Greece | Central Greece | September 2020 | Sparus aurata |
| L. SUSI | V. parahaemolyticus | Philippines | Philippines | April 2018 | Shrimp |
| V1 | V. alginolyticus | Greece | - | May 2018 | Sparus aurata |
| LAR194 | V. mediterranei | Greece | Central Greece | May 2020 | Artemia nauplii |
| SM1 | V. harveyi | Greece | Central Greece | - | Seriola dumerili |
| MAN113 | V. splendidus | Greece | Saronikos Gulf | September 2019 | Seriola dumerili |
| Varv A4/1.1 | V. harveyi | Greece | Central Greece | July 2019 | Sparus aurata |
| VH2 | V. harveyi | Greece | Crete | 2007 | Seriola dumerili |
| VhP1 Liv | V. harveyi | Greece | Crete | April 2015 | Seriola dumerili |
| VhP1 Spl | V. harveyi | Greece | Eastern Aegean | September 2015 | Dichentrachus labrax |
| DY05 | V. owensii | Greece | Eastern Aegean | September 2015 | Dichentrachus labrax |
| SA 6.2 | V. owensii | Saudi Arabia | - | July 2019 | Oreochromis niloticus |
| VIB391 | V. campbellii | Thailand | - | October 2016 | Shrimp |
| Kef 56 | V. rotiferianus | Greece | Ionian Islands | May 2020 | Dichentrachus labrax |
| VhSerFre | V. harveyi | Greece | Crete | August 2015 | Seriola dumerili |
| Phaeobacter S26 | Phaeobacter piscinae | Greece | Euboea | July 2013 | Artemia nauplii |
| Efficiency of Plating of Vibrio Phage Virtus | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Species/Strain | Host Range | EOP | |||||||
| 100 | 10−1 | 10−2 | 10−3 | 10−4 | 10−5 | 10−6 | 10−7 | (%) | |
| Vibrio harveyi | |||||||||
| SA 2.1 | ++ | ++ | ++ | + | + | + | + | - | High |
| Varv A4/1.1 | - | - | - | - | - | - | - | - | NF |
| DSM 19623 | ++ | + | + | + | + | - | - | - | Medium |
| Vh No22 | - | - | - | - | - | - | - | - | NF |
| VhP1 Spl | +++ | ++ | ++ | ++ | ++ | ++ | + | + | High |
| VH2 | ++++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | High |
| VhSerFre | ++++ | +++ | +++ | - | - | - | - | - | Medium |
| VhP1 Liv | - | - | - | - | - | - | - | - | NF |
| Kef 75 | +++ | +++ | ++ | ++ | ++ | ++ | + | + | High |
| SM1 | + | - | - | - | - | - | - | - | Low |
| Gal 56 | + | - | - | - | - | - | - | - | Low |
| Gal 77 | - | - | - | - | - | - | - | - | NF |
| Gal 94 | - | - | - | - | - | - | - | - | NF |
| Gal 72 | - | - | - | - | - | - | - | - | NF |
| Kef 62 | - | - | - | - | - | - | - | - | NF |
| Gal 90 | - | - | - | - | - | - | - | - | NF |
| Vibrio parahaemolyticus | |||||||||
| L.SUSI | ++++ | +++ | - | - | - | - | - | - | Medium |
| Vibrio alginolyticus | |||||||||
| DSM2171 | - | - | - | - | - | - | - | - | NF |
| V1 | - | - | - | - | - | - | - | - | NF |
| Vibrio mediterranei | |||||||||
| LAR194 | +++ | + | + | - | - | - | - | - | Low |
| Vibrio splendidus | |||||||||
| MAN113 | - | - | - | - | - | - | - | - | NF |
| Vibrio owensii | |||||||||
| SA 6.2 | ++++ | +++ | +++ | ++ | + | - | - | - | Medium |
| DY05 | - | - | - | - | - | - | - | - | NF |
| Vibrio campbellii | |||||||||
| VIB391 | ++++ | +++ | +++ | ++ | + | - | - | - | Medium |
| Vibrio rotiferianus | |||||||||
| Kef56 | + | - | - | - | - | - | - | - | Low |
| Predicted Functions | Start | End | Length | Direction | InterPro | NCBI CDD Best Hit | E-Value | |
|---|---|---|---|---|---|---|---|---|
| ORF1 | Hypothetical protein | 44 | 478 | 434 | Forward | pfam18925|DUF5675 | 7.64 × 10−30 | |
| ORF2 | TMhelix containing protein | 480 | 1019 | 539 | Forward | |||
| ORF3 | AAA family ATPase | 1143 | 2366 | 1223 | Forward | cl38936|P-loop_NTPase super family | 2.77 × 10−38 | |
| ORF4 | Winged helix-turn-helix DNA-binding domain protein | 2753 | 4246 | 1493 | Forward | cl41463| PspC_subgroup_2 super family | 6.13 × 10−3 | |
| ORF5 | Hypothetical protein | 4246 | 4839 | 593 | Forward | |||
| ORF6 | Replicative DNA helicase | 4824 | 6272 | 1448 | Forward | cl38936| P-loop_NTPase super family | 1.16 × 10−20 | |
| ORF7 | Hypothetical protein | 6344 | 6928 | 584 | Forward | |||
| ORF8 | DNA primase | 6921 | 7907 | 986 | Forward | cl40740| DnaG super family | 1.07 × 10−9 | |
| ORF9 | Hypothetical protein | 7921 | 8841 | 920 | Forward | |||
| ORF10 | Putative DNA helicase | 8889 | 10,316 | 1427 | Forward | cl34083| SSL2 super family | 1.43 × 10−32 | |
| ORF11 | Putative DNA-binding domain protein | 10,415 | 10,867 | 452 | Forward | cl02600 | HTH_MerR-SF super family | 1.22 × 10−3 | |
| ORF12 | Hypothetical protein | 10,874 | 11,257 | 383 | Forward | |||
| ORF13 | RecA protein | 11,268 | 12,344 | 1076 | Forward | IPR013765 | cl38936 | P-loop_NTPase super family | 1.94 × 10−50 |
| ORF14 | Hypothetical protein | 12,325 | 12,738 | 413 | Forward | No hit | ||
| ORF15 | Rupredoxin-type fold protein | 12,728 | 13,723 | 995 | Forward | cl37788 NOB1_Zn_bind super family | 8.11 × 10−3 | |
| ORF16 | Ribonuclease-H-like domain protein | 13,723 | 14,277 | 554 | Forward | No hit | ||
| ORF17 | Hypothetical protein | 14,386 | 15,360 | 974 | Forward | No hit | ||
| ORF18 | Hypothetical protein | 15,427 | 15,696 | 269 | Forward | No hit | ||
| ORF19 | Hypothetical protein | 15,721 | 16,464 | 743 | Forward | No hit | ||
| ORF20 | Hypothetical protein | 16,542 | 17,273 | 731 | Forward | No hit | ||
| ORF21 | Coil containing protein | 17,288 | 18,097 | 809 | Forward | No hit | ||
| ORF22 | Terminase large subunit | 18,084 | 20,057 | 1973 | Forward | No hit | ||
| ORF23 | Hypothetical protein | 20,076 | 20,318 | 242 | Forward | No hit | ||
| ORF24 | Hypothetical protein | 20,434 | 20,697 | 263 | Forward | No hit | ||
| ORF25 | Hypothetical protein | 20,739 | 21,005 | 266 | Forward | No hit | ||
| ORF26 | Coil-containing protein | 20,995 | 21,417 | 422 | Forward | No hit | ||
| ORF27 | Pyruvate, phosphate dikinase | 21,445 | 23,520 | 2075 | Forward | IPR010121 | cl35801 PRK09279 super family | 1.21 × 10−180 |
| ORF28 | Hypothetical protein | 23,525 | 24,013 | 488 | Forward | No hit | ||
| ORF29 | Putative protein-tyrosine phosphatase | 24,006 | 24,551 | 545 | Forward | cl28904 PTP_DSP_cys super family | 8.26 × 10−17 | |
| ORF30 | Hypothetical protein | 24,577 | 25,620 | 1043 | Forward | No hit | ||
| ORF31 | Transporter | 25,631 | 26,473 | 842 | Forward | No hit | ||
| ORF32 | DNA polymerase I | 26,473 | 28,830 | 2357 | Forward | IPR002298 | cl34031 PolA super family | 8.32 × 10−71 |
| ORF33 | Portal proten | 28,832 | 30,697 | 1865 | Forward | IPR006944 | cl19194 Phage_portal super family | 2.47 × 10−39 |
| ORF34 | Minor head protein | 30,701 | 31,789 | 1088 | Forward | cl10072 Phage_Mu_F super family | 1.69 × 10−3 | |
| ORF35 | HNH endonuclease | 31,789 | 32,304 | 515 | Forward | pfam13392 HNH_3 | 5.16 × 10−10 | |
| ORF36 | DNA methylotransferase | 32,304 | 32,954 | 650 | Forward | IPR007757 | cl01947 MT-A70 super family | 2.11 × 10−28 |
| ORF37 | TMhelix containing protein | 33,138 | 33,554 | 416 | Forward | No hit | ||
| ORF38 | Hypothetical protein | 33,566 | 34,021 | 455 | Forward | No hit | ||
| ORF39 | Putative DNA polymerase I | 34,149 | 34,544 | 395 | Forward | cl02626 DNA_pol_A super family | 1.35 × 10−5 | |
| ORF40 | Hypothetical protein | 34,547 | 34,732 | 185 | Forward | No hit | ||
| ORF41 | Hypothetical protein | 34,755 | 35,123 | 368 | Forward | No hit | ||
| ORF42 | Hypothetical protein | 35,134 | 35,394 | 260 | Forward | No hit | ||
| ORF43 | Hypothetical protein | 35,401 | 35,889 | 488 | Forward | No hit | ||
| ORF44 | Hypothetical protein | 35,892 | 36,176 | 284 | Forward | No hit | ||
| ORF45 | Hypothetical protein | 36,182 | 36,481 | 299 | Forward | No hit | ||
| ORF46 | Hypothetical protein | 36,566 | 36,874 | 308 | Forward | No hit | ||
| ORF47 | Hypothetical protein | 36,926 | 38,263 | 1337 | Forward | No hit | ||
| ORF48 | Hypothetical protein | 38,309 | 38,902 | 593 | Forward | No hit | ||
| ORF49 | SEC-C motif protein | 38,928 | 39,290 | 362 | Forward | pfam02810 SEC-C | 2.55 × 10−6 | |
| ORF50 | Hypothetical protein | 39,299 | 39,469 | 170 | Forward | No hit | ||
| ORF51 | putative zinc- or iron-chelating-domain-containing protein | 39,466 | 39,927 | 461 | Forward | No hit | ||
| ORF52 | Hypothetical protein | 39,911 | 40,432 | 521 | Forward | No hit | ||
| ORF53 | Hypothetical protein | 40,432 | 40,674 | 242 | Forward | No hit | ||
| ORF54 | Hypothetical protein | 40,674 | 40,853 | 179 | Forward | No hit | ||
| ORF55 | Hypothetical protein | 41,325 | 41,765 | 440 | Forward | No hit | ||
| ORF56 | Hypothetical protein | 41,753 | 41,956 | 203 | Forward | No hit | ||
| ORF57 | Hypothetical protein | 41,953 | 42,135 | 182 | Forward | No hit | ||
| ORF58 | Hypothetical protein | 42,132 | 42,845 | 713 | Forward | No hit | ||
| ORF59 | Coil containing protein | 42,826 | 43,638 | 812 | Forward | No hit | ||
| ORF60 | Hypothetical protein | 43,644 | 43,952 | 308 | Forward | No hit | ||
| ORF61 | Hypothetical protein | 44,081 | 44,587 | 506 | Forward | No hit | ||
| ORF62 | DUF550-domain-containing protein | 44,651 | 45,433 | 782 | Forward | cl04522 DUF550 super family | 5.51 × 10−7 | |
| ORF63 | Hypothetical protein | 45,417 | 45,575 | 158 | Forward | No hit | ||
| ORF64 | Hypothetical protein | 45,683 | 45,877 | 194 | Forward | No hit | ||
| ORF65 | Hypothetical protein | 45,892 | 46,221 | 329 | Forward | No hit | ||
| ORF66 | Hypothetical protein | 46,274 | 46,669 | 395 | Forward | No hit | ||
| ORF67 | Hypothetical protein | 46,746 | 47,126 | 380 | Forward | No hit | ||
| ORF68 | Hypothetical protein | 47,178 | 47,639 | 461 | Forward | No hit | ||
| ORF69 | Hypothetical protein | 47,636 | 47,962 | 326 | Forward | No hit | ||
| ORF70 | Hypothetical protein | 47,946 | 48,461 | 515 | Forward | No hit | ||
| ORF71 | Hypothetical protein | 48,448 | 48,855 | 407 | Forward | No hit | ||
| ORF72 | Hypothetical protein | 48,861 | 49,031 | 170 | Forward | No hit | ||
| ORF73 | Hypothetical protein | 49,427 | 49,699 | 272 | Forward | No hit | ||
| ORF74 | Hypothetical protein | 49,714 | 49,920 | 206 | Forward | No hit | ||
| ORF75 | Hypothetical protein | 50,017 | 50,349 | 332 | Forward | No hit | ||
| ORF76 | Hypothetical protein | 50,408 | 50,743 | 335 | Forward | No hit | ||
| ORF77 | Hypothetical protein | 50,800 | 51,246 | 446 | Forward | No hit | ||
| ORF78 | Hypothetical protein | 51,236 | 51,598 | 362 | Forward | No hit | ||
| ORF79 | Phage protein (ACLAME 851) | 51,644 | 52,147 | 503 | Forward | No hit | ||
| ORF80 | Hypothetical protein | 52,281 | 52,481 | 200 | Forward | No hit | ||
| ORF81 | VHS1018 | 52,523 | 52,774 | 251 | Forward | No hit | ||
| ORF82 | Coil-containing protein | 52,822 | 53,127 | 305 | Forward | No hit | ||
| ORF83 | Hypothetical protein | 53,131 | 53,367 | 236 | Forward | No hit | ||
| ORF84 | Hypothetical protein | 53,380 | 53,580 | 200 | Forward | No hit | ||
| ORF85 | Hypothetical protein | 53,580 | 53,723 | 143 | Forward | No hit | ||
| ORF86 | Hypothetical protein | 53,847 | 54,092 | 245 | Forward | No hit | ||
| ORF87 | Hypothetical protein | 54,108 | 54,341 | 233 | Forward | No hit | ||
| ORF88 | Hypothetical protein | 54,468 | 54,668 | 200 | Forward | No hit | ||
| ORF89 | DNA Helicase | 54,715 | 54,993 | 278 | Forward | cl28899 DEAD-like_helicase_N super family | 8.48 × 10−3 | |
| ORF90 | Hypothetical protein | 54,990 | 55,247 | 257 | Forward | No hit | ||
| ORF91 | Hypothetical protein | 55,244 | 56,659 | 1415 | Forward | No hit | ||
| ORF92 | Hypothetical protein | 56,753 | 57,649 | 896 | Forward | No hit | ||
| ORF93 | Hypothetical protein | 57,724 | 58,683 | 959 | Forward | No hit | ||
| ORF94 | Hypothetical protein | 58,814 | 59,128 | 314 | Forward | No hit | ||
| ORF95 | Coil-containing protein | 59,106 | 59,690 | 584 | Forward | No hit | ||
| ORF96 | Hypothetical protein | 59,766 | 60,128 | 362 | Forward | No hit | ||
| ORF97 | Hypothetical protein | 60,179 | 60,571 | 392 | Forward | No hit | ||
| ORF98 | Hypothetical protein | 60,568 | 61,002 | 434 | Forward | No hit | ||
| ORF99 | Hypothetical protein | 61,079 | 61,432 | 353 | Forward | No hit | ||
| ORF100 | Hypothetical protein | 61,438 | 61,932 | 494 | Forward | No hit | ||
| ORF101 | Hypothetical protein | 61,998 | 62,693 | 695 | Forward | No hit | ||
| ORF102 | Hypothetical protein | 62,697 | 62,978 | 281 | Forward | No hit | ||
| ORF103 | Hypothetical protein | 63,124 | 63,414 | 290 | Forward | No hit | ||
| ORF104 | Hypothetical protein | 63,428 | 63,730 | 302 | Forward | No hit | ||
| ORF105 | Hypothetical protein | 63,742 | 64,266 | 524 | Forward | No hit | ||
| ORF106 | Hypothetical protein | 64,266 | 64,538 | 272 | Forward | No hit | ||
| ORF107 | Coil-containing protein | 64,607 | 65,395 | 788 | Forward | No hit | ||
| ORF108 | Hypothetical protein | 65,388 | 67,358 | 1970 | Forward | No hit | ||
| ORF109 | Hypothetical protein | 67,424 | 67,774 | 350 | Forward | No hit | ||
| ORF110 | XkdF | 67,903 | 68,514 | 611 | Forward | cl24270 Peptidase_S78_2 super family | 2.76 × 10−19 | |
| ORF111 | Hypothetical protein | 68,517 | 69,686 | 1169 | Forward | cl36772 2A1904 super family | 2.50 × 10−5 | |
| ORF112 | Major capsid protein | 69,767 | 70,717 | 950 | Forward | IPR024455 | cl27082 Phage_capsid super family | 4.87 × 10−5 |
| ORF113 | Coil-containing protein | 70,795 | 71,064 | 269 | Forward | No hit | ||
| ORF114 | Head completion adaptor | 71,080 | 71,724 | 644 | Forward | No hit | ||
| ORF115 | Neck protein | 71,721 | 72,185 | 464 | Forward | No hit | ||
| ORF116 | Tail-completion protein | 72,182 | 72,664 | 482 | Forward | No hit | ||
| ORF117 | Major tail protein | 72,718 | 73,512 | 794 | Forward | IPR016893 | No hit | |
| ORF118 | Hypothetical protein | 73,601 | 74,035 | 434 | Forward | No hit | ||
| ORF119 | VHS1060 protein | 74,122 | 74,235 | 113 | Forward | No hit | ||
| ORF120 | Tail length tape measure protein | 74,241 | 78,374 | 4133 | Forward | pfam10145 PhageMin_Tail | 1.22 × 10−22 | |
| ORF121 | Hypothetical protein | 78,371 | 78,757 | 386 | Forward | No hit | ||
| ORF122 | Hypothetical protein | 78,767 | 79,756 | 989 | Forward | No hit | ||
| ORF123 | Hypothetical protein | 79,766 | 80,659 | 893 | Forward | No hit | ||
| ORF124 | Hypothetical protein | 80,662 | 81,960 | 1298 | Forward | No hit | ||
| ORF125 | Hypothetical protein | 81,963 | 82,487 | 524 | Forward | No hit | ||
| ORF126 | Hypothetical protein | 82,488 | 82,790 | 302 | Forward | No hit | ||
| ORF127 | Hypothetical protein | 82,787 | 82,960 | 173 | Forward | No hit |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Droubogiannis, S.; Katharios, P. Genomic and Biological Profile of a Novel Bacteriophage, Vibrio phage Virtus, Which Improves Survival of Sparus aurata Larvae Challenged with Vibrio harveyi. Pathogens 2022, 11, 630. https://doi.org/10.3390/pathogens11060630
Droubogiannis S, Katharios P. Genomic and Biological Profile of a Novel Bacteriophage, Vibrio phage Virtus, Which Improves Survival of Sparus aurata Larvae Challenged with Vibrio harveyi. Pathogens. 2022; 11(6):630. https://doi.org/10.3390/pathogens11060630
Chicago/Turabian StyleDroubogiannis, Stavros, and Pantelis Katharios. 2022. "Genomic and Biological Profile of a Novel Bacteriophage, Vibrio phage Virtus, Which Improves Survival of Sparus aurata Larvae Challenged with Vibrio harveyi" Pathogens 11, no. 6: 630. https://doi.org/10.3390/pathogens11060630
APA StyleDroubogiannis, S., & Katharios, P. (2022). Genomic and Biological Profile of a Novel Bacteriophage, Vibrio phage Virtus, Which Improves Survival of Sparus aurata Larvae Challenged with Vibrio harveyi. Pathogens, 11(6), 630. https://doi.org/10.3390/pathogens11060630