
Mast-Cell Response to Leishmania mexicana and Sand-Fly Salivary Proteins Is Modulated by Orchiectomy


 ,
,  and
and
Abstract
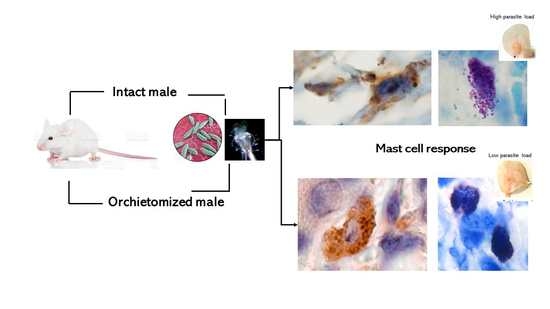
:
1. Introduction
2. Results
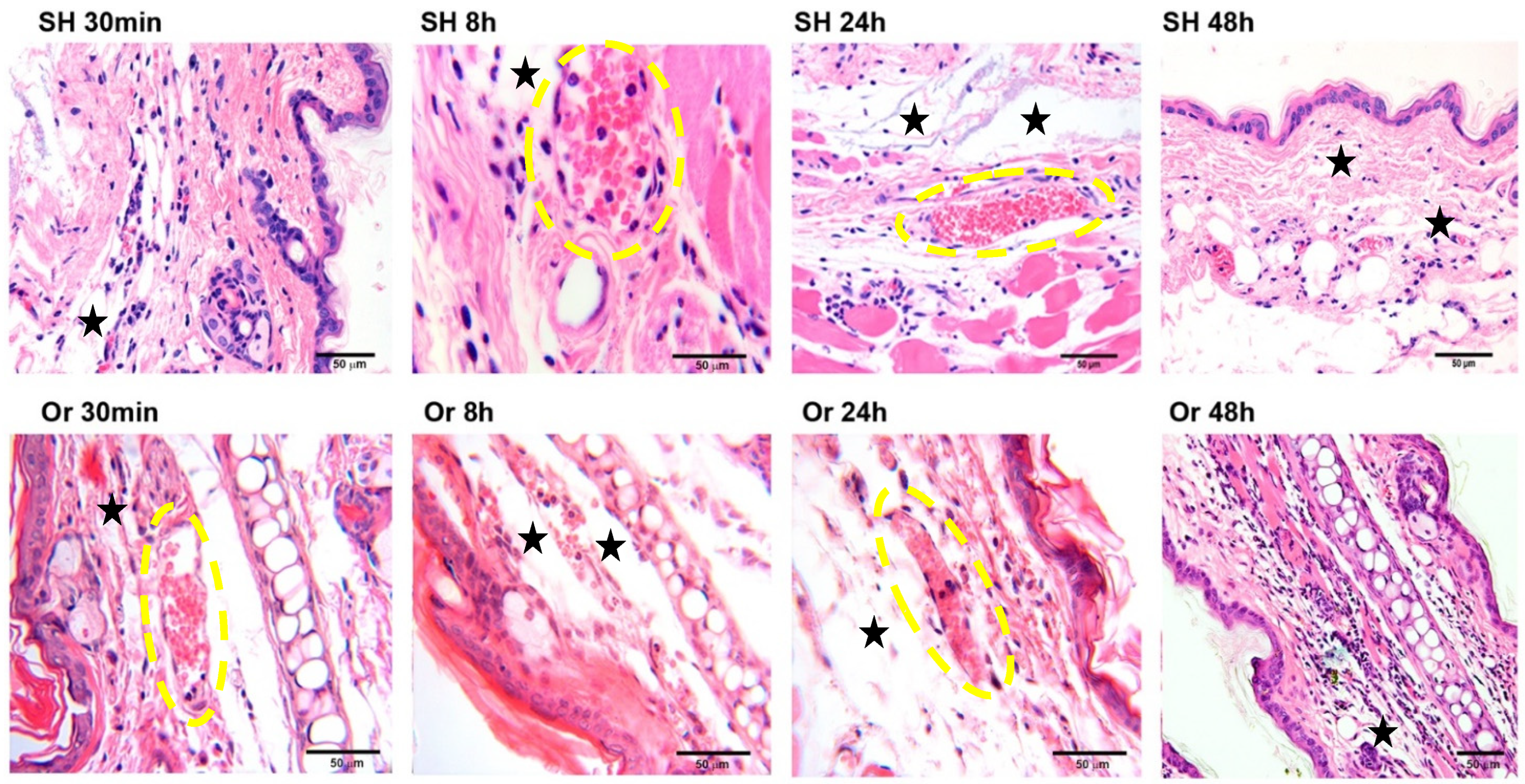
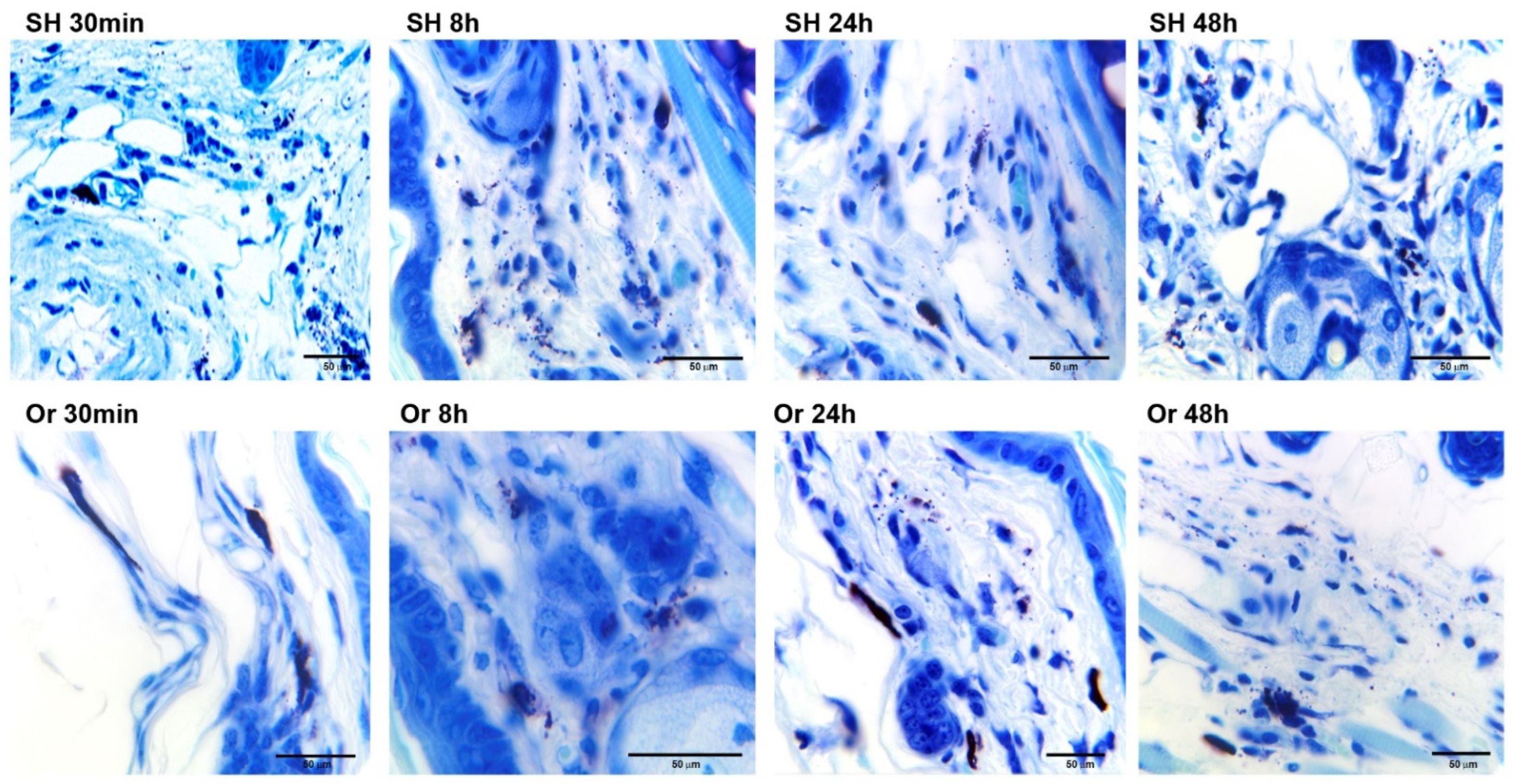
2.1. Ear Tissue Histology after Inoculation of L. mexicana and Vector Salivary Proteins
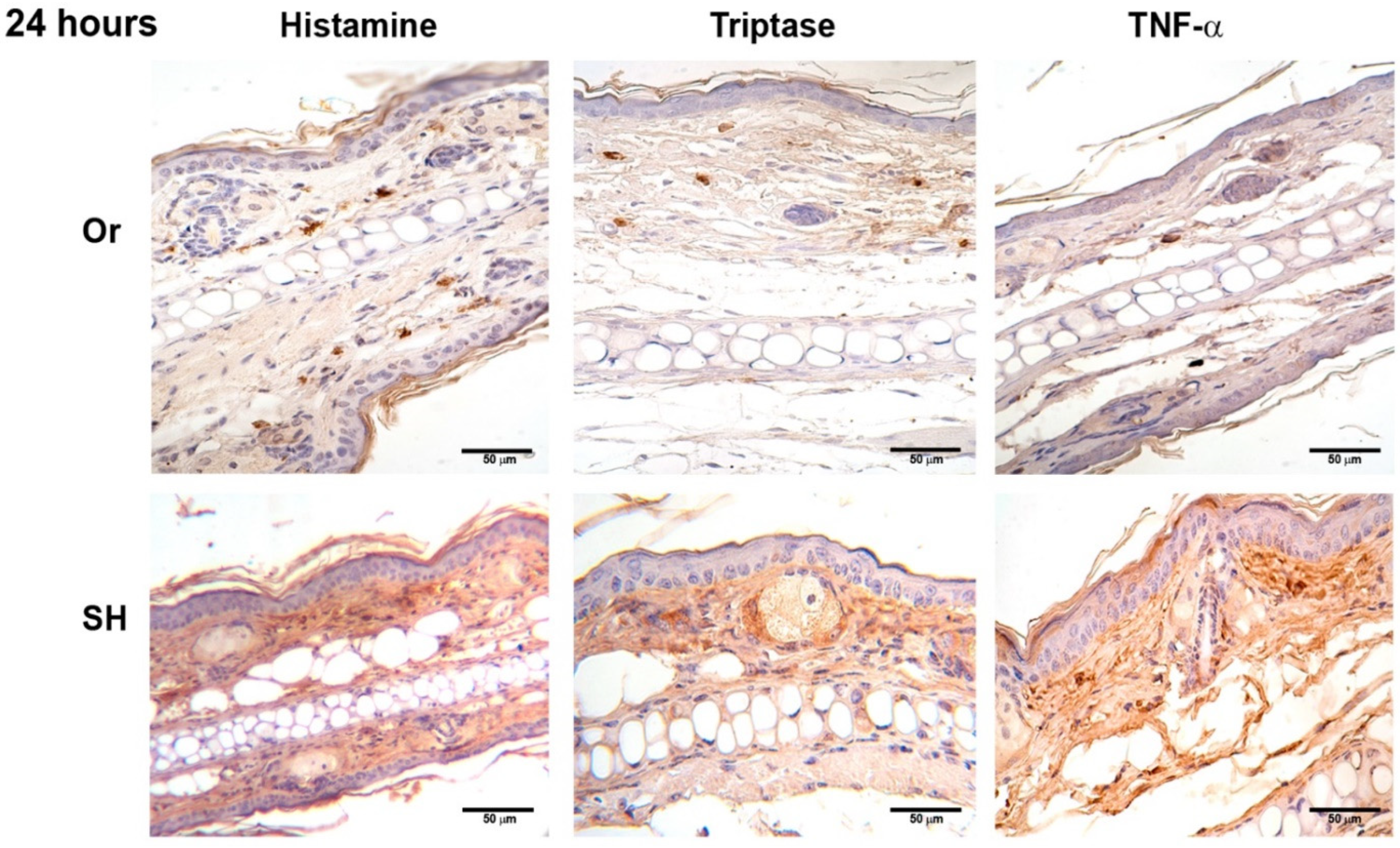
2.2. Immunohistochemistry of Ears after Inoculation of L. mexicana and Vector Salivary Gland Lysates: Inflammatory Mediators Released after MC Activation
2.3. Chronic Infection of L. mexicana Together with Vector Salivary Gland Lysates in Orchiectomized BALB/c Mice
3. Discussion
4. Materials and Methods
4.1. Animals and Experimental Groups
4.2. Surgical Procedure
4.3. Parasite Culture
4.4. Salivary Gland Collection
4.5. Infection Procedure
4.6. Histology and Immunohistochemistry
4.7. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Krystel-Whittemore, M.; Dileepan, K.N.; Wood, J.G. Mast Cell: A Multi-Functional Master Cell. Front. Immunol. 2016, 6, 620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crivellato, E.; Ribatti, D. The mast cell an evolutionary perspective. Biol. Rev. Camb. Philos. Soc. 2010, 85, 347–360. [Google Scholar] [CrossRef]
- Galli, S.; Tsai, M. Mast cells in allergy and infection: Versatile effector and regulatory cells in innate and adaptive immunity. Eur. J. Immunol. 2010, 40, 1843–1851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Silva, M.Z.E.; Jamur, M.C.; Oliver, C. Mast Cell Function: A New Vision of an Old Cell. J. Histochem. Cytochem. 2014, 62, 69–738. [Google Scholar] [CrossRef] [PubMed]
- Soman, N.A.; St John, A.L. Mast cell-orchestrated immunity to pathogens. Nat. Rev. Immunol. 2010, 10, 440–4525. [Google Scholar]
- Harvima, I.; Nilsson, G. Mast cells as regulators of skin inflammation and immunity. Acta Derm. Venereol. 2011, 91, 644–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villaseñor-Cardoso, M.I.; Salaiza, N.; Delgado, J.; Gutiérrez-Kobeh, L.; Pérez-Torres, A.; Becker, I. Mast cells are activated by Leishmania mexicana LPG and regulate the disease outcome depending on the genetic background of the host. Parasite Immunol. 2008, 30, 425–434. [Google Scholar] [CrossRef]
- Romão, P.R.T.; Da Costa Santiago, H.; Ramos, C.D.L.; De Oliveira, C.F.E.; Monteiro, M.C.; De Queiroz Cunha, F.; Vieira, L.Q. Mast cell degranulation contributes to susceptibility to Leishmania major. Parasite Immunol. 2009, 31, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Kaye, P.M.; Beattie, L. Lessons from other diseases: Granulomatous inflammation in leishmaniasis. Semin. Immunopathol. 2016, 38, 249–260. [Google Scholar] [CrossRef] [Green Version]
- Andrade, B.B.; De Oliveira, C.I.; Brodskyn, C.C.; Barral, A.; Barral-Netto, M. Role of Sand Fly Saliva in Human and Experimental Leishmaniasis: Current Insights. Scan. J. Immunol. 2007, 66, 122–127. [Google Scholar] [CrossRef]
- Rohousova, I.; Volf, P. Sand Fly: Effects on host immune response and Leishmania transmission. Folia Parasitol. 2006, 53, 161–171. [Google Scholar] [CrossRef] [Green Version]
- Gomes, R.; Oliveira, F.; Teixeira, C.; Meneses, C.; Gilmore, D.C.; Elnaiem, D.A.; Kamhawi, S.; Valenzuela, J.V. Immunity to Sand Fly Salivary Protein LJM11 Modulates Host Response to Vector-Transmitted Leishmania Conferring Ulcer-Free Protection. Soc. Invest. Dermatol. 2012, 132, 2735–2743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muñoz-Cruz, S.; Togno-Pierce, C.; Morales-Montor, J. Non-reproductive effects of sex steroids: Their immunoregulatory role. Curr. Top. Med. Chem. 2011, 11, 1714–1727. [Google Scholar] [PubMed]
- Taneja, V. Sex Hormones Determine Immune Response. Front. Immunol. 2018, 9, 1931. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J. Sex differences and cross-immunity in DBA/2 mice infected with L. mexicana and L. major. Parasitology 1988, 96, 297–302. [Google Scholar] [CrossRef]
- Mock, B.A.; Nacy, C.A. Hormonal modulation of sex differences in resistance to Leishmania major systemic infections. Infect. Immun. 1988, 56, 3316–3319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, M.; Alexander, J.; Blackwell, J.M. Genetic analysis of Leishmania mexicana Infection in mice: Single gene (Scl-2) controlled predisposition to cutaneous lesion development. J. Immunol. 1990, 17, 89–100. [Google Scholar] [CrossRef]
- Satoskar, A.; Alexander, J. Sex-determined susceptibility and differential IFN-gamma and TNF-alpha mRNA expression in DBA/2 mice infected with Leishmania mexicana. Immunology 1995, 84, 1–4. [Google Scholar] [PubMed]
- Satoskar, A.; Al-Quassi, H.H.; Alexander, J. Sex-determined resistance against Leishmania mexicana is associated with the preferential induction of a Th1-like response and IFN-gamma production by female but not male DBA/2 mice. Immunol. Cell Biol. 1988, 76, 159–166. [Google Scholar] [CrossRef] [PubMed]
- D’agostino, P.; Milano, S.; Barbera, C.; Di Bella, G.; La Rosa, M.; Ferlazzo, V.; Farruggio, R.; Miceli, D.; Miele, M.; Castagnetta, L.; et al. Sex hormones modulate inflammatory mediators produced by macrophages. Ann. N. Y. Acad. Sci. 1999, 876, 426–429. [Google Scholar] [CrossRef]
- Piccinni, M.P.; Giudizi, M.G.; Biagiotti, R.; Beloni, L.; Giannarini, L.; Sampognaro, S.; Parronchi, P.; Manetti, R.; Annunziato, F.; Livi, C. Progesterone favors the development of human T helper cells producing Th2-type cytokines and promotes both IL-4 production and membrane expression in established Th1 cell clones. J. Immunol. 1995, 155, 128–133. [Google Scholar]
- Isaac-Márquez, A.; Lezama-Dávila, C. Cytokine regulation of female-macrophage resistance to L. mexicana parasites. Parasite Immunol. 2019, 42, e12685. [Google Scholar]
- Liu, L.; Wang, L.; Zhao, Y.; Wang, Y.; Wang, Z.; Qiao, Z. Testosterone attenuates p38 MAPK pathway during Leishmania donovani infection of macrophages. Parasitol. Res. 2006, 99, 189–193. [Google Scholar] [CrossRef]
- Zhang, H.; Zhao, J.; Wang, P.; Qiao, Z. Effect of testosterone on Leishmania donovani infection of macrophages. Parasitol. Res. 2001, 87, 674–676. [Google Scholar] [CrossRef] [PubMed]
- Yin, G.; Guo, Z.; Yin, K.; Zhao, J. Effect of testosterone on Leishmania donovani infection levels of murine bone marrow-derived macrophages. Zhongguo Ji Sheng Chong Xue Yu Ji Sheng Chong Bing Za Zhi 1988, 16, 251–255. [Google Scholar]
- Sánchez-García, L.; Wilkins-Rodriguez, A.; Salaiza-Suazo, N.; Morales-Montor, J.; Becker, I. Dihydrotestosterone enhances growth and infectivity of Leishmania mexicana. Parasite Immunol. 2018, 40, e12512. [Google Scholar] [CrossRef] [PubMed]
- von Stebut, E. Immunology of cutaneous leishmaniasis: The role of mast cells, phagocytes and dendritic cells for protective immunity. Eur. J. Dermatol. 2007, 17, 115–122. [Google Scholar] [PubMed]
- Maurer, M.; Lopez Kostka, S.; Siebenhaar, F.; Moelle, K.; Metz, M.; Knop, J.; von Stebut, E. Skin mast cells control T cell-dependent host defense in Leishmania major infections. FASEB J. 2006, 20, 2460–2467. [Google Scholar] [CrossRef] [PubMed]
- von Stebut, E.; Metz, M.; Milon, M.; Knop, J.; Maurer, M. Early macrophage influx to sites of cutaneous granuloma formation is dependent on MIP-1α/β released from neutrophils recruited by mast cell-derived TNFα. FASEB Blood 2006, 101, 210–215. [Google Scholar] [CrossRef]
- Bidri, M.; Vouldoukis, I.M. Evidence for direct interaction between mast cells and Leishmania parasites. Parasite Immunol. 1997, 19, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Dantas, C.; Carvalho de Souza, C.; França, C.; Quercia, V.; Crocco, L.; Esteves, A. Histopathology of Leishmania major infection: Revisiting L. major histopathology in the ear dermis infection model. Mem. Inst. Oswal. Cruz 2009, 104, 918. [Google Scholar]
- Sánchez-García, L. Estudio de la Regulación que Ejercen la Testosterona y Extractos Salivales de Lutzomyia Olmeca Sobre Las Células Cebadas y su Asociación con la Leishmaniasis. Ph.D. Thesis, Universidad Nacional Autónoma de Mexico, Ciudad de Mexico, Mexico, 2018. [Google Scholar]
- Lockard, R.D.; Wilson, M.E.; Rodríguez, N.E. Sex-Related Differences in Immune Response and Symptomatic Manifestations to Infection with Leishmania Species. J. Immunol. Res. 2019, 2019, 4103819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vom Steeg, L.G.; Klein, S.L. Sex steroids mediate bidirectional interactions between hosts and microbes. Hormon. Behav. 2017, 88, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Hurrel, P.B.; Beumann, M.; Heyde, S.; Regli, I.B.; Müller, J.A.; Tachinni-Cottier, F. Frontline Science: Leishmania mexicana amastigotes can replicate within neutrophils. J. Leukoc. Biol. 2017, 102, 1187–1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro-Gomes, F.L.; Sacks, D. The influence of early neutrophil-Leishmania interactions on the host immune response to infection. Front. Cell. Infect. Microbiol. 2012, 59, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Hurrell, B.P.; Schuster, S.; Grün, E.; Coutaz, M.; Williams, R.A.; Held, W.; Bernard Malissen, B.; Malissen, M.; Yousefi, S.; Simon, H.U.; et al. Rapid Sequestration of Leishmania mexicana by Neutrophils Contributes to the Development of Chronic Lesion. PLoS Pathog. 2015, 11, e1004929. [Google Scholar] [CrossRef] [PubMed]
- Soong, L. Modulation of Dendritic Cell Function by Leishmania Parasites. J. Immunol. 2008, 180, 4355–4360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, C.M.; Creer, H.M.; McHowat, J. Potential role for mast cell tryptase in recruitment of inflammatory cells to endothelium. Amer. J. Physiol. Cell. Physiol. 2005, 289, C1485–C1491. [Google Scholar] [CrossRef]
- Pós, Z.; Müller, K.; Novák, I.; Buzás, E.; Solbach, W.; Falus, A.; Laskay, T. Different patterns of the L-histidine decarboxylase (HDC) gene expression in mice resistant and susceptible to experimental cutaneous leishmaniasis. Inflamm. Res. 2004, 53, 38–43. [Google Scholar] [CrossRef]
- Qi, H.; Popov, V.; Soong, L. Leishmania amazonensis-dendritic cell interactions in vitro and the priming of parasite-specific CD4(+) T cells in vivo. J. Immunol. 2001, 5, 4534–4542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzoni, A.; Young, H.; Spitzer, J.; Visintin, A.; Segal, D. Histamine regulates cytokine production in maturing dendritic cells, resulting in altered T cell polarization. J. Clin. Invest. 2001, 108, 1865–1873. [Google Scholar] [CrossRef] [PubMed]
- Caron, G.; Delneste, Y.; Roelants, E.; Duez, C.; Bonnefoy, J.; Pestel, J.; Jeannin, P. Histamine polarizes human dendritic cells into Th2 cell promoting effector dendritic cells. J. Immunol. 2001, 167, 3682–3686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernard, G. Within-Host Parasite Dynamics, Emerging Trade-Off, and Evolution of Virulence with Immune System. Evolution 2003, 57, 489–1497. [Google Scholar]
- Muñoz-Cruz, S.; Mendoza-Rodriguez, Y.; Nava-Castro, K.; Yepez-Mulia, L.; Morales-Montor, J. Gender-related Effect of Se Steroides on Histamine Released and FceRI Expression in Rat Peritoneal Mast Cells. J. Immunol. Res. 2015, 351829, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Shirama, K.; Kohda, M.; Hokano, M. Effects of endocrine glands and hormone replacement on the mast cell count of the Harderian gland of mice. Acta Anat. 1988, 131, 327–331. [Google Scholar] [CrossRef] [PubMed]
- Haywood, M.; Spaliviero, J.; Jimemez, M.; King, N.J.; Handelsman, D.J.; Allan, C.M. Sertoli and germ cell development in hypogonadal (hpg) mice expressing transgenic follicle-stimulating hormone alone or in combination with testosterone. Endocrinology 2003, 144, 509–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.; Chen, Y.T.; Yeh, S.D.; Xu, Q.; Wang, R.S.; Guillou, F.; Lardy, H.; Yeh, S. Infertility with defective spermatogenesis and hypotestosteronemia in male mice lacking the androgen receptor in Sertoli cells. Proc. Natl. Acad. Sci. USA 2004, 101, 6876–6881. [Google Scholar] [CrossRef] [Green Version]
- Sridharan, G.; Shanka, A.A. Toluidine blue: A review of its chemistry and clinical utility. J. Oral Maxillo. Pathol. 2012, 16, 251–255. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Unpaired Test (t-Test) | |||||
|---|---|---|---|---|---|
| Treatments | p Value | p Value Summary | Significantly Different (p < 0.05) | ||
| 30 min | vs. | 30Or | 0.0001 | *** | Yes |
| 8SH | vs. | 8Or | 0.0001 | *** | Yes |
| 24SH | vs. | 24Or | 0.0004 | *** | Yes |
| 48SH | vs. | 48Or | 0.0169 | * | Yes |
| 72SH | vs. | 72Or | 0.1369 | ns | No |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-García, L.; Pérez-Torres, A.; Muñoz-Cruz, S.; Salaiza-Suazo, N.; Morales-Montor, J.; Becker, I. Mast-Cell Response to Leishmania mexicana and Sand-Fly Salivary Proteins Is Modulated by Orchiectomy. Pathogens 2022, 11, 398. https://doi.org/10.3390/pathogens11040398
Sánchez-García L, Pérez-Torres A, Muñoz-Cruz S, Salaiza-Suazo N, Morales-Montor J, Becker I. Mast-Cell Response to Leishmania mexicana and Sand-Fly Salivary Proteins Is Modulated by Orchiectomy. Pathogens. 2022; 11(4):398. https://doi.org/10.3390/pathogens11040398
Chicago/Turabian StyleSánchez-García, Laura, Armando Pérez-Torres, Samira Muñoz-Cruz, Norma Salaiza-Suazo, Jorge Morales-Montor, and Ingeborg Becker. 2022. "Mast-Cell Response to Leishmania mexicana and Sand-Fly Salivary Proteins Is Modulated by Orchiectomy" Pathogens 11, no. 4: 398. https://doi.org/10.3390/pathogens11040398
APA StyleSánchez-García, L., Pérez-Torres, A., Muñoz-Cruz, S., Salaiza-Suazo, N., Morales-Montor, J., & Becker, I. (2022). Mast-Cell Response to Leishmania mexicana and Sand-Fly Salivary Proteins Is Modulated by Orchiectomy. Pathogens, 11(4), 398. https://doi.org/10.3390/pathogens11040398