

Inhibitory Effect of Select Nitrocompounds and Chlorate against Yersinia ruckeri and Yersinia aleksiciae In Vitro

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Tests with Pure Cultures
2.2. Tests with Mixed Microbial Populations
2.3. Statistical Analysis
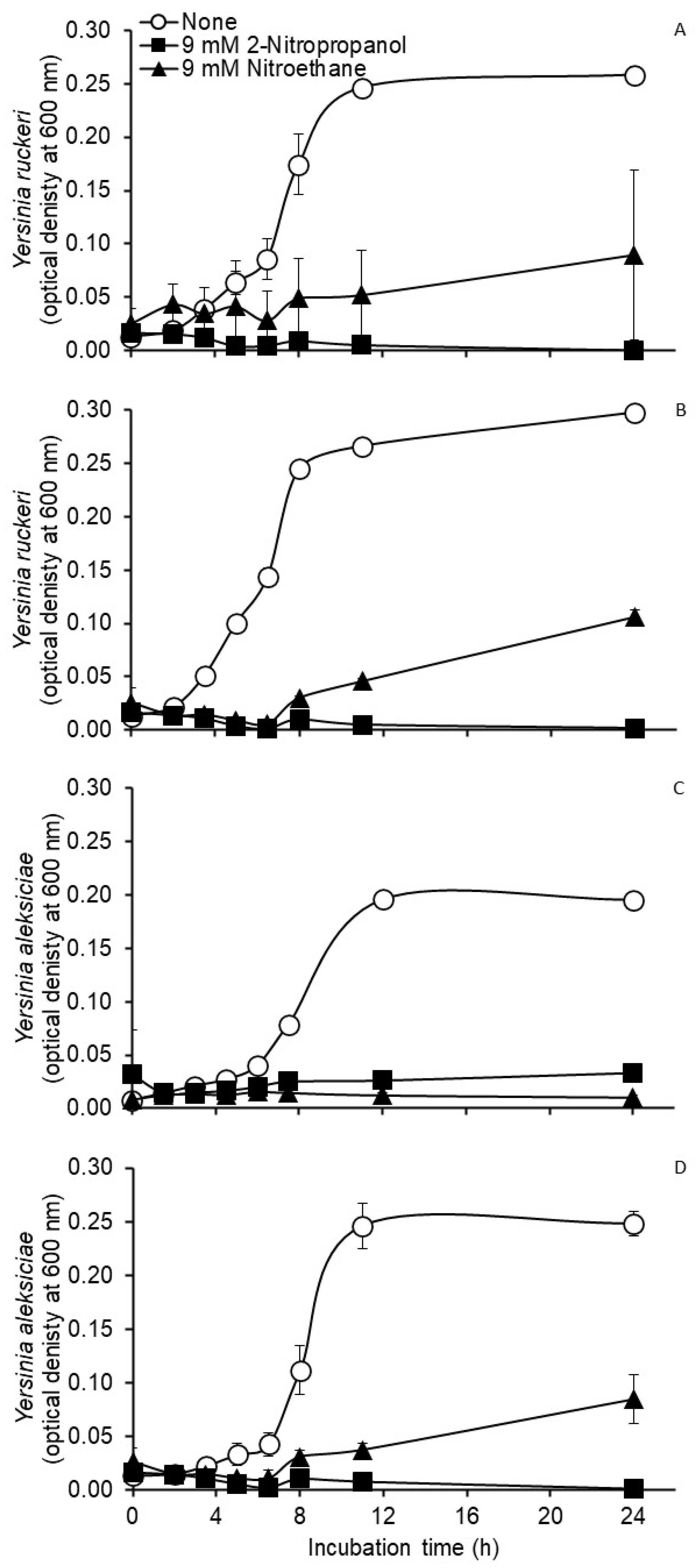
3. Results
3.1. Tests with Pure Cultures
3.2. Tests with Mixed Cultures
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pajdak-Czaus, J.; Platt-Samoraj, A.; Szweda, W.; Siwicki, A.K.; Terech-Majewska, E. Yersinia ruckeri—A threat not only to rainbow trout. Aquac. Res. 2019, 50, 3083–3096. [Google Scholar] [CrossRef]
- Kumar, G.; Menanteau-Ledouble, S.; Saleh, M.; El-Matbouli, M. Yersinia ruckeri, the causative agent of enteric redmouth disease in fish. Vet. Res. 2015, 46, 103. [Google Scholar] [CrossRef]
- Guijarro, J.A.; García-Torrico, A.I.; Cascales, D.; Méndez, J. The infection process of Yersinia ruckeri: Reviewing the pieces of the jigsaw puzzle. Front. Cell. Infect. Microbiol. 2018, 8, 218. [Google Scholar] [CrossRef]
- Thorsen, B.K.; Enger, O.; Norland, S.; Hoff, K.A. Long-term starvation survival of Yersinia ruckeri at different salinities studied by microscopical and flow cytometric methods. Appl. Environ. Microbiol. 1992, 58, 1624–1628. [Google Scholar] [CrossRef]
- Ryckaert, J.; Bossier, P.; D’Herde, K.; Diez-Fraile, A.; Sorgeloos, P.; Haesebrouck, F.; Pasmans, F. Persistence of Yersinia ruckeri in trout macrophages. Fish Shellfish Immunol. 2010, 29, 648–655. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, C.J. The usage of vaccination and antimicrobial agents for control of Yersinia ruckeri. J. Fish Dis. 1991, 14, 291–301. [Google Scholar] [CrossRef]
- Yang, H.; Zhujin, D.; Marana, M.H.; Dalsgaard, I.; Rzgar, J.; Heidi, M.; Asma, K.M.; Per, K.W.; Kurt, B. Immersion vaccines against Yersinia ruckeri infection in rainbow trout: Comparative effects of strain differences. J. Fish Dis. 2021, 44, 1937–1950. [Google Scholar] [CrossRef] [PubMed]
- Orozova, P.; Chikova, V.; Sirakov, I. Diagnostics and antibiotic resistance of Yersinia ruckeri strains isolated from trout fish farms in Bulgaria. Intern. J. Dev. Res. 2014, 4, 2727–2733. [Google Scholar]
- Huang, Y.; Jung, A.; Schäfer, W.-J.; Mock, D.; Michael, B.B.; Runge, M.; Schwarz, S.; Steinhagen, D. Analysis of Yersinia ruckeri strains isolated from trout farms in northwest Germany. Dis. Aquat. Org. 2015, 116, 243–249. [Google Scholar] [CrossRef]
- Wrobel, A.; Leo, J.C.; Linke, D. Overcoming fish defences: The virulence factors of Yersinia ruckeri. Genes 2019, 10, 700. [Google Scholar] [CrossRef]
- Rigos, G.; Stevenson, R. The effect of antibiotic treatment on the establishment of persistent infection with Yersinia ruckeri Serovar II in rainbow trout Oncorhynchus mykiss (Walbaum). Aquac. Int. 2001, 9, 247–253. [Google Scholar] [CrossRef]
- Tobback, E.; Decostere, A.; Hermans, K.; Haesebrouck, F.; Chiers, K. Yersinia ruckeri infections in salmonid fish. J. Fish Dis. 2007, 30, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.C.; Jung, Y.S.; Oliver, C.E.; Horrocks, S.M.; Genovese, K.J.; Harvey, R.B.; Callaway, T.R.; Edrington, T.S.; Nisbet, D.J. Effects of nitrate or nitro-supplementation, with or without added chlorate, on Salmonella enterica serovar Typhimurium and Escherichia coli in swine feces. J. Food Prot. 2007, 70, 308–315. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Božić, A.; Anderson, R.; Alzola-Alvarez, C.; Ruiz-Barrera, O.; Corral-Luna, A.; Castillo-Castillo, Y.; Arzola-Rubio, A.; Poole, T.; Harvey, R.; Hume, M.; et al. Inhibition of multidrug-resistant Staphylococci by sodium chlorate and select nitro- and medium chain fatty acid compounds. J. Appl. Microbiol. 2019, 126, 1508–1518. [Google Scholar] [CrossRef] [PubMed]
- Nisbet, D.J.; Callaway, T.R.; Edrington, T.S.; Anderson, R.C.; Poole, T.L. Effects of ionophores on Enterococcus faecalis and E. faecium growth in pure and mixed ruminal culture. Foodborne Path. Dis. 2008, 5, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Stewart, V. Nitrate respiration in relation to facultative metabolism in enterobacteria. Microbiol. Rev. 1988, 52, 190–232. [Google Scholar] [CrossRef]
- Moreno-Vivián, C.; Cabello, P.; Martínez-Luque, M.; Blasco, R.; Castillo, F. Prokaryotic nitrate reduction: Molecular properties and functional distinction among bacterial nitrate reductases. J. Bacteriol. 1999, 181, 6573–6584. [Google Scholar] [CrossRef]
- Wauters, G.; Janssens, M.; Steigerwalt, A.G.; Brenner, D.J. Yersinia mollaretii sp. nov. and Yersinia bercovieri sp. nov., formerly called Yersinia enterocolitica Biogroups 3A and 3B. Int. J. Syst. Bacteriol. 1988, 38, 424–429. [Google Scholar] [CrossRef]
- Reuter, S.; Connor, T.R.; Barquist, L.; Walker, D.; Feltwell, T.; Harris, S.R.; Fookes, M.; Hall, M.E.; Petty, N.K.; Fuchs, T.M.; et al. Parallel independent evolution of pathogenicity within the genus Yersinia. Proc. Nat. Acad. Sci. USA 2014, 111, 6768–6773. [Google Scholar] [CrossRef]
- Oltmann, L.F.; Claassen, V.P.; Kastelein, P.; Reijnders, W.N.M.; Stouthamer, A.H. Influence of tungstate on the formation and activities of four reductases in Proteus mirabilis: Identification of two new molybdo-enzymes: Chlorate reductase and tetrathionate reductase. FEBS Lett. 1979, 106, 43–46. [Google Scholar] [CrossRef]
- Chen, P.E.; Cook, C.; Stewart, A.C.; Nagarajan, N.; Sommer, D.D.; Pop, M.; Thomason, B.; Thomason, M.P.; Lentz, S.; Nolan, N.; et al. Genomic characterization of the Yersinia genus. Genome Biol. 2010, 11, R1. [Google Scholar] [CrossRef] [PubMed]
- Oltmann, L.F.; Reijnders, W.N.M.; Stouthamer, A.H. The correlation between the protein composition of cytoplasmic membranes and the formation of nitrate reductase A, chlorate reductase C and tetrathionate reductase in Proteus mirabilis wild type and some chlorate resistant mutants. Arch. Microbiol. 1976, 111, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Callaway, T.R.; Anderson, R.C.; Anderson, T.J.; Poole, T.L.; Bischoff, K.M.; Kubena, L.F.; Nisbet, D.J. Escherichia coli O157:H7 becomes resistant to sodium chlorate in pure culture, but not in mixed culture or in vivo. J. Appl. Microbiol. 2001, 91, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.S.; Anderson, R.C.; Callaway, T.R.; Edrington, T.S.; Genovese, K.J.; Harvey, R.B.; Poole, T.L.; Nisbet, D.J. Inhibitory activity of 2-nitropropanol against select foodborne pathogens in vitro. Lett. Appl. Microbiol. 2004, 39, 471–476. [Google Scholar] [CrossRef]
- Jung, Y.S.; Anderson, R.C.; Edrington, T.S.; Genovese, K.J.; Byrd, J.A.; Callaway, T.R.; Nisbet, D.J. Experimental use of 2-nitropropanol for reduction of Salmonella Typhimurium in the ceca of broiler chicks. J. Food Prot. 2004, 67, 1945–1947. [Google Scholar] [CrossRef] [PubMed]
- Dimitrijevic, M.; Anderson, R.C.; Callaway, T.R.; Jung, Y.S.; Harvey, R.B.; Ricke, S.C.; Nisbet, D.J. Inhibitory effect of select nitroalkanes on growth rate of Listeria monocytogenes in vitro. J. Food Prot. 2006, 69, 1061–1065. [Google Scholar] [CrossRef]
- Horrocks, S.M.; Jung, Y.S.; Huwe, J.K.; Harvey, R.B.; Ricke, S.C.; Carstens, G.E.; Callaway, T.R.; Anderson, R.C.; Ramlachan, N.; Nisbet, D.J. Effects of short-chain nitrocompounds against Campylobacter jejuni and Campylobacter coli in vitro. J. Food Sci. 2007, 72, M50-–M55. [Google Scholar] [CrossRef]
- Adhikari, P.; Cosby, D.; Cox, N.; Kim, W. Effect of dietary supplementation of nitrocompounds on Salmonella colonization and ileal immune gene expression, in laying hens challenged with Salmonella Enteritidis. Poult. Sci. 2017, 96, 4280–4286. [Google Scholar] [CrossRef]
- Anderson, R.C.; Rasmussen, M.A. Use of a novel nitrotoxin-metabolizing bacterium to reduce ruminal methane production. Bioresour. Technol. 1998, 64, 89–95. [Google Scholar] [CrossRef]
- Anderson, R.C.; Callaway, T.R.; Van Kessel, J.S.; Jung, Y.S.; Edrington, T.S.; Nisbet, D.J. Effect of select nitrocompounds on ruminal fermentation; an initial look at their potential to reduce economic and environmental costs associated with ruminal methanogenesis. Bioresour. Technol. 2003, 90, 59–63. [Google Scholar] [CrossRef]
- Anderson, R.C.; Krueger, N.A.; Stanton, T.B.; Callaway, T.R.; Edrington, T.S.; Harvey, R.B.; Jung, Y.S.; Nisbet, D.J. Effects of select nitrocompounds on in vitro ruminal fermentation during conditions of limiting or excess added reductant. Bioresour. Technol. 2008, 99, 8655–8661. [Google Scholar] [CrossRef]
- Gutierrez-Bañuelos, H.; Anderson, R.C.; Carstens, G.E.; Tedeschi, L.O.; Pinchak, W.E.; Cabrera-Diaz, E.; Krueger, N.A.; Callaway, T.R.; Nisbet, D.J. Effects of nitroethane and monensin on ruminal fluid fermentation characteristics and nitrocompound-metabolizing bacterial populations. J. Agric. Food Chem. 2008, 56, 4650–4658. [Google Scholar] [CrossRef] [PubMed]
- Božic, A.K.; Anderson, R.C.; Carstens, G.E.; Ricke, S.C.; Callaway, T.R.; Yokoyama, M.T.; Wang, J.K.; Nisbet, D.J. Effects of the methane-inhibitors nitrate, nitroethane, lauric acid, Lauricidin® and the Hawaiian marine algae, Chaetoceros, on ruminal fermentation in vitro. Bioresour. Technol. 2009, 100, 4017–4025. [Google Scholar] [CrossRef]
- Kim, W.K.; Weeks, L.J.; Anderson, R.C.; Nisbet, D.J.; Dunkley, K.; Ricke, S.C. Effects of nitrocompounds on uric acid-utilizing microorganisms, nitrogen retention, and microbial community in laying hen manure. J. Environ. Sci. Health Part B 2009, 44, 403–406. [Google Scholar] [CrossRef]
- Ochoa-García, P.A.; Anderson, R.C.; Arévalos-Sánchez, M.M.; Rodríguez-Almeida, F.A.; Félix-Portillo, M.; Muro-Reyes, A.; Božić, A.K.; Arzola-Álvarez, C.; Corral-Luna, A. Astragallus mollissimus plant extract: A strategy to reduce ruminal methanogenesis. Trop. Anim. Health Prod. 2021, 53, 436. [Google Scholar] [CrossRef]
- Teng, P.-Y.; Kim, W.K. Roles of nitrocompounds in inhibition of foodborne bacteria, parasites, and methane production in economic animals. Animals 2021, 11, 923. [Google Scholar] [CrossRef]
- Zhang, Z.W.; Wang, Y.L.; Wang, W.K.; Li, Y.H.; Cao, Z.J.; Li, L.; Yang, H.J. The inhibitory action mode of nitrocompounds on in vitro rumen methanogenesis: A comparison of nitroethane, 2-nitroethanol and 2-nitro-1-propanol. J. Agric. Sci. 2019, 157, 471–479. [Google Scholar] [CrossRef]
- Anderson, R.C.; Carstens, G.E.; Miller, R.K.; Callaway, T.R.; Schultz, C.L.; Edrington, T.S.; Harvey, R.B.; Nisbet, D.J. Effect of oral nitroethane and 2-nitropropanol administration on methane-producing activity and volatile fatty acid production in the ovine rumen. Bioresour. Technol. 2006, 97, 2421–2426. [Google Scholar] [CrossRef]
- Gutierrez-Bañuelos, H.; Anderson, R.C.; Carstens, G.E.; Slay, L.J.; Ramlachan, N.; Horrocks, S.M.; Callaway, T.R.; Edrington, T.S.; Nisbet, D.J. Zoonotic bacterial populations, gut fermentation characteristics and methane production in feedlot steers during oral nitroethane treatment and after the feeding of an experimental chlorate product. Anaerobe 2007, 13, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.G.; Anderson, R.C.; Carstens, G.E.; Gutierrez-Bañuelos, H.; McReynolds, J.L.; Slay, L.J.; Callaway, T.R.; Nisbet, D.J. Effects of oral nitroethane on enteric methane emissions and ruminal fermentation in cattle. Anim. Feed Sci. Technol. 2011, 166–167, 275–281. [Google Scholar] [CrossRef]
- Zhang, Z.W.; Wang, Y.L.; Chen, Y.Y.; Wang, W.K.; Zhang, L.T.; Luo, H.L.; Yang, H.J. Nitroethanol in comparison with monensin exhibits greater feed efficiency through inhibiting rumen methanogenesis more efficiently and persistently in feedlotting lambs. Animals 2019, 9, 784. [Google Scholar] [CrossRef]
- Zhang, Z.W.; Wang, Y.L.; Chen, Y.Y.; Zhang, L.T.; Zhang, Y.J.; Liu, Y.Q.; Guo, Y.X.; Yang, H.J. The dietary supplemental effect of nitroethanol in comparison with monensin on methane emission, growth performance and carcass characteristics in female lambs. Animals 2021, 11, 327. [Google Scholar] [CrossRef] [PubMed]
- Viedma, P.M.; Abriouel, H.; Omar, N.B.; López, R.L.; Gálvez, A. Antistaphylococcal effect of enterocin AS-48 in bakery ingredients of vegetable origin, alone and in combination with selected antimicrobials. J. Food Sci. 2009, 74, M384–M389. [Google Scholar] [CrossRef]
- Angermaier, L.; Simon, H. On the reduction of aliphatic and aromatic nitro compounds by Clostridia, the role of ferredoxin and its stabilization. Hoppe-Seylers Z. Physiol. Chem. 1983, 364, 961–975. [Google Scholar] [CrossRef]
- Ruiz-Barrera, O.; Anderson, R.C.; Hume, M.E.; Corrales-Millan, J.; Castillo-Castillo, Y.; Corral-Luna, A.; Guevara-Valdez, J.L.; Salinas-Chavira, J.; Rodriguez-Muela, C.; Arzola-Alvarez, C. Short chain nitrocompounds as a treatment of layer hen manure and litter; effects on in vitro survivability of Salmonella, generic E. coli and nitrogen metabolism. J. Environ. Sci. Health B 2017, 52, 23–29. [Google Scholar] [CrossRef]
- Ruiz-Barrera, O.; Ontiveros-Magadan, M.; Anderson, R.C.; Byrd, J.A.; Hume, M.E.; Latham, E.A.; Nisbet, D.J.; Arzola-Alvarez, C.; Salinas-Chavira, J.; Castillo-Castillo, Y. Nitro-treatment of composted poultry litter; effects on Salmonella, E. coli and nitrogen metabolism. Bioresour. Technol. 2020, 310, 123459. [Google Scholar] [CrossRef]
- Barz, M.; Beimgraben, C.; Staller, T.; Germer, F.; Opitz, F.; Marquardt, C.; Schwarz, C.; Gutekunst, K.; Vanselow, K.H.; Schmitz, R.; et al. Distribution analysis of hydrogenases in surface waters of marine and freshwater environments. PLoS ONE 2010, 5, e13846. [Google Scholar] [CrossRef] [PubMed]
- Jordaan, K.; Lappan, R.; Dong, X.; Aitkenhead, I.J.; Bay, S.K.; Chiri, E.; Wieler, N.; Meredith, L.K.; Cowan, D.A.; Chown, S.L.; et al. Hydrogen-oxidizing bacteria are abundant in desert soils and strongly stimulated by hydration. mSystems 2020, 5, e01131-20. [Google Scholar] [CrossRef] [PubMed]
- Greening, C.; Islam, Z.F.; Bay, S.K. Hydrogen is a major lifeline for aerobic bacteria. Trends Microbiol. 2022, 30, 330–337. [Google Scholar] [CrossRef]
- Müller, M.; Hopfner, K.-P.; Witte, G. C-di-AMP recognition by Staphylococcus aureus PstA. Febs Lett. 2015, 589, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Hofbauer, S.; Hagmüller, A.; Schaffner, I.; Mlynek, G.; Krutzler, M.; Stadlmayr, G.; Pirker, K.F.; Obinger, C.; Daims, H.; Djinović-Carugo, K.; et al. Structure and heme-binfing properties of HemQ (chlorite dismutase-like protein) from Listeria monocytogenes. Arch. Biochem. Biophys. 2015, 574, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Rutala, W.A.; Weber, D.J. Guideline for Disinfection and Sterilization in Healthcare Facilities, 2008 Update: May 2019. Centers for Disease Control and Prevention, National Center for Emerging and Zoonotic Infectious Diseases (NCEZID), Division of Healthcare Quality Promotion (DHQP). 2019. Available online: https://www.cdc.gov/infectioncontrol/guidelines/disinfection/ (accessed on 26 April 2022).
- Smith, D.J.; Anderson, R.C. Toxicity and metabolism of nitroalkanes and substituted nitroalkanes. J. Agric. Food Chem. 2013, 61, 763–779. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Mean Specific Growth Rate (h−1) | Maximum Optical Density (OD 600 nm) | |||
|---|---|---|---|---|
| Treatment | Yersinia ruckeri | Yersinia aleksiciae | Yersinia ruckeri | Yersinia aleksiciae |
| None | 0.179 a | 0.463 a | 0.200 b | 0.290 a |
| 9 mM Nitrate | 0.160 a | 0.459 a | 0.215 ab | 0.281 ab |
| 9 mM Chlorate | 0.173 a | 0.369 b | 0.229 a | 0.217 c |
| 9 mM 2-Nitropropanol | <0.001 b | 0.014 c | 0.045 c | 0.050 d |
| 9 mM Nitrate plus 9 mM chlorate | 0.170 a | 0.427 a | 0.228 a | 0.278 b |
| 9 mM 2-Nitropropanol plus 9 mM chlorate | <0.001 b | <0.001 c | 0.047 c | 0.050 d |
| p value | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Standard error of the mean | 0.0157 | 0.0107 | 0.0060 | 0.0036 |
| Y. aleksiciae Protein a | Percent Amino Acid Identity | ||
|---|---|---|---|
| Yersinia mollaretii b | Escherichia coli str. K-12 substr. MG1655 b | Yersinia ruckeri b | |
| Respiratory nitrate reductase alpha chain | 98.16 | 85.98 | No significant similarity found |
| Respiratory nitrate reductase beta chain | 97.86 | 87.97 | No significant similarity found |
| Respiratory nitrate reductase delta chain | 92.09 | 64.90 | No significant similarity found |
| Respiratory nitrate reductase gamma chain | 98.68 | 84.98 | No significant similarity found |
| Treatment | Mean Specific Growth Rate (h−1) | Maximum Optical Density (OD 600 nm) | ||
|---|---|---|---|---|
| Yersinia ruckeri | Yersinia Aleksiciae | Yersinia ruckeri | Yersinia aleksiciae | |
| None | 0.424 a | 0.364 ab | 0.233 a | 0.176 a |
| 9 mM 2-Nitropropanol | <0.001 d | <0.001 d | 0.005 e | <0.001 d |
| 9 mM 3-Nitropropanol | <0.001 d | <0.001 d | 0.004 e | <0.000 d |
| 9 mM 2-Nitroethanol | <0.001 d | <0.001 d | 0.005 e | 0.002 d |
| 9 mM Nitroethane | 0.082 c | 0.141 c | 0.047 b | 0.007 d |
| 9 mM Ethyl nitroacetate | 0.165 b | 0.146 bc | 0.076 d | 0.081 c |
| 9 mM Ethyl-2-nitropropionate | 0.237 b | 0.144 c | 0.115 b | 0.074 c |
| 9 mM Ethanol | 0.401 a | 0.387 a | 0.213 a | 0.172 a |
| 9 mM Methanol | 0.456 a | 0.344 abc | 0.226 a | 0.186 a |
| 9 mM Isopropanol | 0.460 a | 0.327 abc | 0.222 a | 0.132 b |
| p value | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Standard error of the mean | 0.0262 | 0.0904 | 0.0083 | 0.0094 |
| Treatment | Mean Specific Growth Rate (h−1) | Maximum Optical Density (OD 600 nm) | ||
|---|---|---|---|---|
| Yersinia ruckeri | Yersinia aleksiciae | Yersinia ruckeri | Yersinia aleksiciae | |
| None | 0.230 a | 0.327 a | 0.255 a | 0.187 a |
| 2.25 mM 2-Nitropropanol | 0.012 b | <0.001 b | 0.022 c | 0.027 b |
| 4.5 mM 2-Nitropropanol | 0.008 b | <0.001 b | 0.020 c | 0.013 b |
| 9 mM 2-Nitropropanol | <0.001 b | <0.001 b | 0.022 c | 0.014 b |
| p value | <0.0001 | 0.0077 | <0.0001 | <0.0001 |
| Standard error of the mean | 0.0182 | 0.0775 | 0.0038 | 0.0032 |
| Treatment | Net Change in log10 Colony Forming Units/mL Incubation Mixture | |
|---|---|---|
| Yersinia ruckeri Cultured Aerobically at 28 °C in Freshly Collected Tilapia Fish Water | Yersinia aleksiciae Cultured Anaerobically at 37 °C in Half-Strength BHI Broth Inoculated with 0.2% wt/vol Freshly Collected Pig Feces | |
| None | 0.94 a | −0.33 a |
| 9 mM 2-Nitropropanol | −4.63 e | −4.83 d |
| 9 mM 2-Nitroethanol | −4.63 e | −4.83 d |
| 9 mM Nitroethane | −0.14 d | −1.18 c |
| 9 mM Ethyl nitroacetate | 0.81 b | −0.82 b |
| 9 mM Ethyl-2-nitropropionate | 0.40 c | −0.83 b |
| 9 mM Sodium chlorate | 1.38 a | −1.25 c |
| p value | <0.0001 | <0.0001 |
| Standard error of the mean | 0.1031 | 0.6400 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Latham, E.A.; Anderson, R.C.; Wottlin, L.R.; Poole, T.L.; Crippen, T.L.; Schlosser, W.D.; Harvey, R.B.; Hume, M.E. Inhibitory Effect of Select Nitrocompounds and Chlorate against Yersinia ruckeri and Yersinia aleksiciae In Vitro. Pathogens 2022, 11, 1381. https://doi.org/10.3390/pathogens11111381
Latham EA, Anderson RC, Wottlin LR, Poole TL, Crippen TL, Schlosser WD, Harvey RB, Hume ME. Inhibitory Effect of Select Nitrocompounds and Chlorate against Yersinia ruckeri and Yersinia aleksiciae In Vitro. Pathogens. 2022; 11(11):1381. https://doi.org/10.3390/pathogens11111381
Chicago/Turabian StyleLatham, Elizabeth A., Robin C. Anderson, Lauren R. Wottlin, Toni L. Poole, Tawni L. Crippen, Wayne D. Schlosser, Roger B. Harvey, and Michael E. Hume. 2022. "Inhibitory Effect of Select Nitrocompounds and Chlorate against Yersinia ruckeri and Yersinia aleksiciae In Vitro" Pathogens 11, no. 11: 1381. https://doi.org/10.3390/pathogens11111381
APA StyleLatham, E. A., Anderson, R. C., Wottlin, L. R., Poole, T. L., Crippen, T. L., Schlosser, W. D., Harvey, R. B., & Hume, M. E. (2022). Inhibitory Effect of Select Nitrocompounds and Chlorate against Yersinia ruckeri and Yersinia aleksiciae In Vitro. Pathogens, 11(11), 1381. https://doi.org/10.3390/pathogens11111381