
Balantioides coli Fecal Excretion in Hunted Wild Cervids (Cervus elaphus and Dama dama) from Portugal
 ,
,  , ,
, , 
 and
and
Abstract
:1. Introduction
2. Results
3. Discussion
4. Materials and Methods
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Areán, V.M.; Koppisch, E. Balantidiasis: A review and report of cases. Am. J. Pathol. 1956, 32, 1089–1115. [Google Scholar] [PubMed]
- Ponce-Gordo, F.; García-Rodríguez, J. Balantioides coli . Res. Vet. Sci. 2021, 135, 424–431. [Google Scholar] [CrossRef]
- Bouwknegt, M.; Devleesschauwer, B.; Graham, H.; Robertson, L.J.; van der Giessen, J.W.; the Euro-Fbp Workshop Participants. Prioritisation of food-borne parasites in Europe, 2016. Eurosurveillance 2018, 23, 17–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giarratana, F.; Nalbone, L.; Napoli, E.; Lanzo, V.; Panebianco, A. Prevalence of Balantidium coli (Malmsten, 1857) infection in swine reared in South Italy: A widespread neglected zoonosis. Vet. World 2021, 14, 1044–1049. [Google Scholar] [CrossRef]
- Cho, H.S.; Shin, S.S.; Park, N.Y. Balantidiasis in the gastric lymph nodes of Barbary sheep (Ammotragus lervia): An incidental finding. J. Vet. Sci. 2006, 7, 207–209. [Google Scholar] [CrossRef] [Green Version]
- Headley, S.A.; Kummala, E.; Sukura, A. Balantidium coli-infection in a Finnish horse. Vet. Parasitol. 2008, 158, 129–132. [Google Scholar] [CrossRef] [PubMed]
- Schuster, F.L.; Ramirez-Avila, L. Current world status of Balantidium coli. Clin. Microbiol. Rev. 2008, 21, 626–638. [Google Scholar] [CrossRef] [Green Version]
- Vasilakopoulou, A.; Dimarongona, K.; Samakovli, A.; Papadimitris, K.; Avlami, A. Balantidium coli pneumonia in an immunocompromised patient. Scand. J. Infect. Dis. 2003, 35, 144–146. [Google Scholar] [CrossRef]
- García-Rodríguez, J.J.; Köster, P.C.; Ponce-Gordo, F. Cyst detection and viability assessment of Balantioides coli in environmental samples: Current status and future needs. Food Waterborne Parasitol. 2022, 26, e00143. [Google Scholar] [CrossRef]
- Li, J.; Wang, Z.; Karim, M.R.; Zhang, L. Detection of human intestinal protozoan parasites in vegetables and fruits: A review. Parasit. Vectors 2020, 13, 380. [Google Scholar] [CrossRef]
- Byun, J.W.; Park, J.H.; Moon, B.Y.; Lee, K.; Lee, W.K.; Kwak, D.; Lee, S.H. Identification of Zoonotic Balantioides coli in Pigs by Polymerase Chain Reaction-Restriction Fragment Length Polymorphism (PCR-RFLP) and Its Distribution in Korea. Animal 2021, 11, 2659. [Google Scholar] [CrossRef] [PubMed]
- Plutzer, J.; Karanis, P. Neglected waterborne parasitic protozoa and their detection in water. Water Res. 2016, 101, 318–332. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Hinojosa, P.Ú.; Espinoza-Ríos, J.; Carlin-Ronquillo, A.; Pinto-Valdivia, J.L.; Salas-Dueñas, Y.; Zare-Morales, W. Colonic balantidiasis: Report of a fatal case and review of the literature. Rev. Gastroenterol. Peru 2019, 39, 284–287. [Google Scholar]
- Bianchi, M.V.; Mello, L.S.; Wentz, M.F.; Panziera, W.; Soares, J.F.; Sonne, L.; Driemeier, D.; Pavarini, S.P. Fatal parasite-induced enteritis and typhlocolitis in horses in Southern Brazil. Rev. Bras. Parasitol. Vet. 2019, 28, 443–450. [Google Scholar] [CrossRef] [Green Version]
- Lankester, F.; Mätz-Rensing, K.; Kiyang, J.; Jensen, S.A.; Weiss, S.; Leendertz, F.H. Fatal ulcerative colitis in a western lowland gorilla (Gorilla gorilla gorilla). J. Med. Primatol. 2008, 37, 297–302. [Google Scholar] [CrossRef] [Green Version]
- Yazar, S.; Altuntas, F.; Sahin, I.; Atambay, M. Dysentery caused by Balantidium coli in a patient with non-Hodgkin’s lymphoma from Turkey. World J. Gastroenterol. 2004, 10, 458–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hechenbleikner, E.M.; McQuade, J.A. Parasitic colitis. Clin. Colon. Rectal. Surg. 2015, 28, 79–86. [Google Scholar] [CrossRef] [Green Version]
- Barmon, B.C.; Begum, N.; Labony, S.S.; Kundu, U.K.; Dey, A.R.; Dey, T.R. Study of gastrointestinal parasites of deer at Char Kukri Mukri in Bhola District. Bangl. J. Vet. Med. 2014, 12, 27–33. [Google Scholar] [CrossRef] [Green Version]
- Khatun, M.M.; Begum, N.; Mamun, M.A.; Mondal, M.M.; Azam, M.S. Coprological study of gastrointestinal parasites of captive animals at Rangpur Recreational Garden and Zoo in Bangladesh. J. Threat. Taxa 2014, 6, 6142–6147. [Google Scholar] [CrossRef] [Green Version]
- Rahman, S.; Dey, A.; Kundu, U.; Begum, N. Investigation of gastrointestinal parasites of herbivores at Dhaka National Zoological Garden of Bangladesh. J. Bangladesh Agril. Univ. 2014, 12, 79–85. [Google Scholar] [CrossRef] [Green Version]
- Valente, A.M.; Acevedo, P.; Figueiredo, A.M.; Fonseca, C.; Torres, R.T. Overabundant wild ungulate populations in Europe: Management with consideration of socio-ecological consequences. Mammal Rev. 2020, 50, 353–366. [Google Scholar] [CrossRef]
- Torres, R.T.; Cunha, M.V.; Araujo, D.; Ferreira, H.; Fonseca, C.; Palmeira, J.D. A walk on the wild side: Wild ungulates as potential reservoirs of multi-drug resistant bacteria and genes, including Escherichia coli harbouring CTX-M beta-lactamases. Environ. Pollut. 2022, 306, 119367. [Google Scholar] [CrossRef]
- Torres, R.T.; Cunha, M.V.; Ferreira, H.; Fonseca, C.; Palmeira, J.D. A high-risk carbapenem-resistant Pseudomonas aeruginosa clone detected in red deer (Cervus elaphus) from Portugal. Sci. Total Environ. 2022, 829, 154699. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Feng, Y.; Santin, M. Host Specificity of Enterocytozoon bieneusi and Public Health Implications. Trends Parasitol. 2019, 35, 436–451. [Google Scholar] [CrossRef] [PubMed]
- Griffin, J.F.; Mackintosh, C.G. Tuberculosis in deer: Perceptions, problems and progress. Vet. J. 2000, 160, 202–219. [Google Scholar] [CrossRef]
- Figueiredo, A.M.; Valente, A.M.; Fonseca, C.; de Carvalho, L.M.; Torres, R.T. Endoparasite diversity of the main wild ungulates in Portugal. Wildlife Biol. 2020, 1, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Dashti, A.; Köster, P.C.; Bailo, B.; Sánchez de las Matas, A.; Habela, M.A.; Rivero-Juarez, A.; Vicente, J.; Serrano, E.; Arnal, M.C.; Fernández de Luco, D.; et al. Low occurrence and limited zoonotic potential of Cryptosporidium spp., Giardia duodenalis, and Balantioides coli infections in free-ranging and farmed wild ungulates in Spain. Transbound. Emerg. Dis. 2022. under review. [Google Scholar]
- Ponce-Gordo, F.; Jimenez-Ruiz, E.; Martínez-Díaz, R.A. Tentative identification of the species of Balantidium from ostriches (Struthio camelus) as Balantidium coli-like by analysis of polymorphic DNA. Vet. Parasitol. 2008, 157, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Ponce-Gordo, F.; Fonseca-Salamanca, F.; Martínez-Díaz, R.A. Genetic heterogeneity in internal transcribed spacer genes of Balantidium coli (Litostomatea, Ciliophora). Protist 2011, 162, 774–794. [Google Scholar] [CrossRef]
- Pomajbíková, K.; Oborník, M.; Horák, A.; Petrželková, K.J.; Grim, J.N.; Levecke, B.; Todd, A.; Mulama, M.; Kiyang, J.; Modrý, D. Novel insights into the genetic diversity of Balantidium and Balantidium-like cyst-forming ciliates. PLoS Negl. Trop. Dis. 2013, 7, e2140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hindsbo, O.; Nielsen, C.V.; Andreassen, J.; Willingham, A.L.; Bendixen, M.; Nielsen, M.A.; Nielsen, N.O. Age-dependent occurrence of the intestinal ciliate Balantidium coli in pigs at a Danish research farm. Acta Vet. Scand. 2000, 41, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Damriyasa, I.M.; Bauer, C. Prevalence and age-dependent occurrence of intestinal protozoan infections in suckling piglets. Berl. Munch. Tierarztl. Wochenschr. 2006, 119, 287–290. [Google Scholar]
- Ferry, T.; Bouhour, D.; De Monbrison, F.; Laurent, F.; Dumouchel-Champagne, H.; Picot, S.; Piens, M.A.; Granier, P. Severe peritonitis due to Balantidium coli acquired in France. Eur. J. Clin. Microbiol. Infect. Dis. 2004, 23, 393–395. [Google Scholar] [CrossRef] [PubMed]
- Walzer, P.D.; Judson, F.N.; Murphy, K.B.; Healy, G.R.; English, D.K.; Schultz, M.G. Balantidiasis outbreak in Truk. Am. J. Trop. Med. Hyg. 1973, 22, 33–41. [Google Scholar] [CrossRef]
- Owen, I.L. Parasitic zoonoses in Papua New Guinea. J. Helminthol. 2005, 79, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, J.; Torres, R.T.; Acevedo, P.; Santos, J.P.V.; Barros, T.; Serrano, E.; Fonseca, C. Propagule pressure and land cover changes as main drivers of red and roe deer expansion in mainland Portugal. Divers. Distrib. 2018, 24, 551–564. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K. Estimation of the number of nucleotide substitutions when there are strong transition-transversion and G+C-content biases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Variable | Total (n) | B. coli Positive (n) | Frequency (%) | p-Value |
|---|---|---|---|---|
| Host species | 0.717 | |||
| C. elaphus | 95 | 4 | 4.21 | |
| D. dama | 35 | 2 | 5.71 | |
| Sex | 0.938 | |||
| Male | 67 | 3 | 4.48 | |
| Female | 63 | 3 | 4.76 | |
| Age | 0.763 | |||
| Juvenile | 14 | 1 | 7.14 | |
| Semi-adult | 7 | 0 | 0 | |
| Adult | 109 | 5 | 4.59 | |
| Fecal consistency | --- | |||
| Formed | 130 | 6 | 4.62 | |
| Diarrhea | 0 | 0 | 0 | |
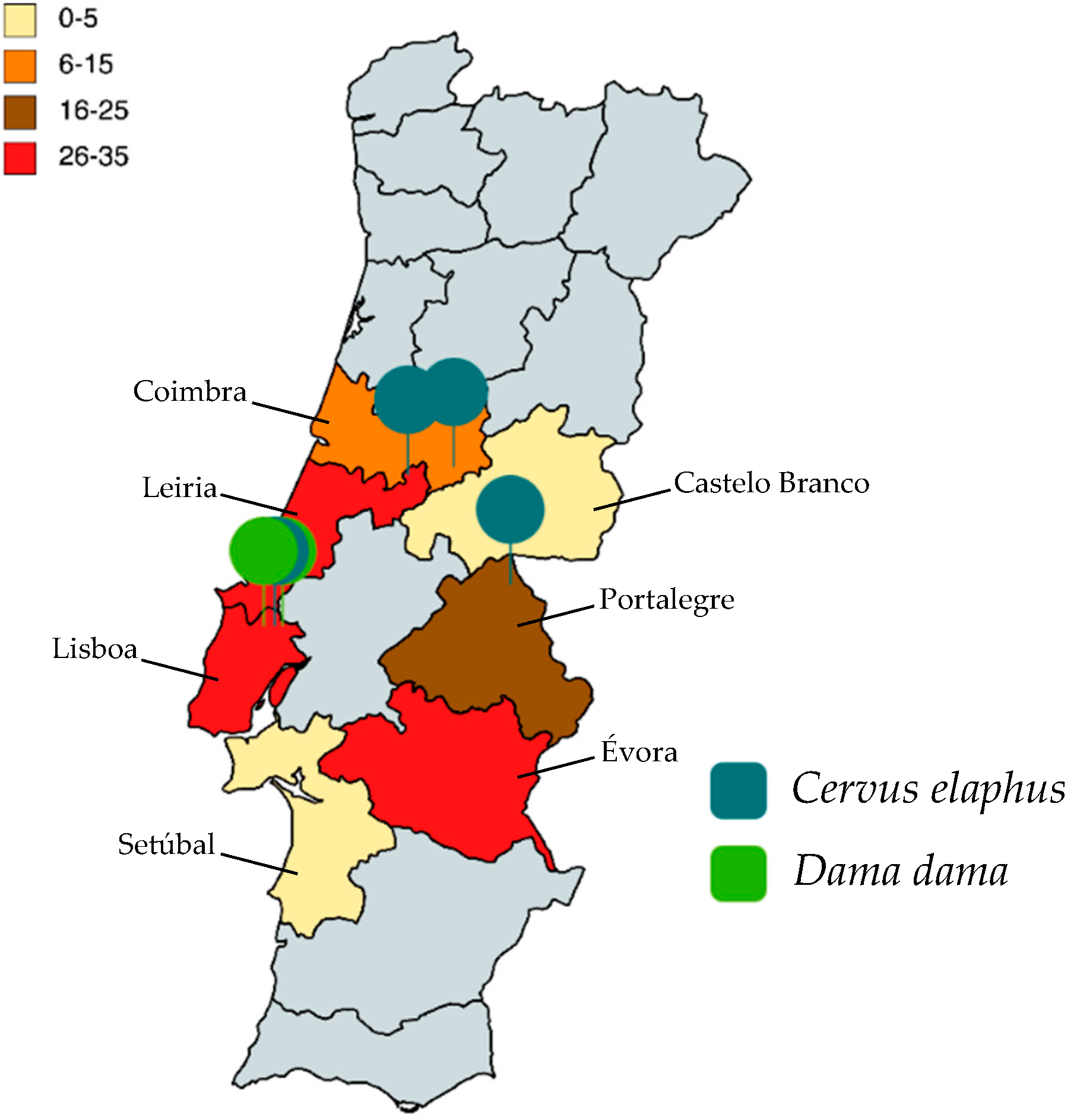
| Region | 0.718 | |||
| Castelo Branco | 1 | 0 | 0 | |
| Setúbal | 1 | 0 | 0 | |
| Coimbra | 10 | 1 | 10 | |
| Portalegre | 23 | 1 | 4.35 | |
| Évora | 30 | 0 | 0 | |
| Lisboa | 30 | 3 | 10 | |
| Leiria | 35 | 1 | 2.86 |
| Sample ID | Host Species | Age | Sex | Location |
|---|---|---|---|---|
| C24 | Cervus elaphus | Juvenile | Female | Coimbra |
| C39 | Cervus elaphus | Adult | Male | Leiria |
| C68 | Cervus elaphus | Adult | Female | Portalegre |
| C120 | Dama dama | Adult | Male | Lisbon |
| C140 | Dama dama | Adult | Female | Lisbon |
| C143 | Cervus elaphus | Adult | Male | Lisbon |
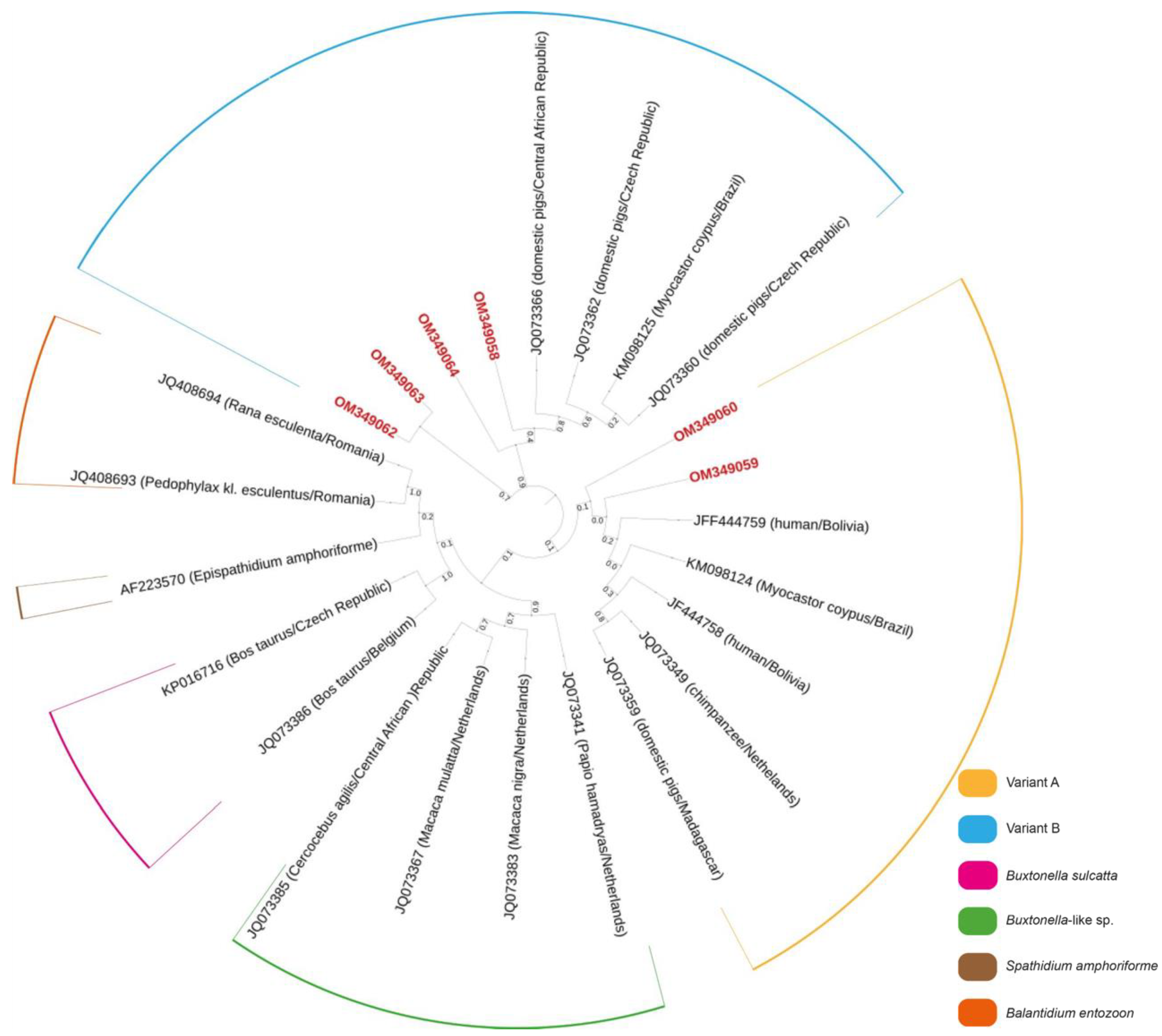
| Sequence | Host Species | Variant | GenBank ID | Identity (%) | Reference Sequence |
|---|---|---|---|---|---|
| C24 | Cervus elaphus | B | OM349058 | 99.5 | MT258438 |
| C39 | Cervus elaphus | A | OM349059 | 86.7 | MZ676835 |
| C68 | Cervus elaphus | A | OM349060 | 88.2 | JQ073346 |
| C120 | Dama dama | B | OM349062 | 83.0 | MT252079 |
| C140 | Dama dama | B | OM349063 | 99.7 | MT258433 |
| C143 | Cervus elaphus | B | OM349064 | 98.9 | MT258438 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mega, J.; Santos-Silva, S.; Loureiro, A.; Palmeira, J.D.; Torres, R.T.; Rivero-Juarez, A.; Carmena, D.; Mesquita, J. Balantioides coli Fecal Excretion in Hunted Wild Cervids (Cervus elaphus and Dama dama) from Portugal. Pathogens 2022, 11, 1242. https://doi.org/10.3390/pathogens11111242
Mega J, Santos-Silva S, Loureiro A, Palmeira JD, Torres RT, Rivero-Juarez A, Carmena D, Mesquita J. Balantioides coli Fecal Excretion in Hunted Wild Cervids (Cervus elaphus and Dama dama) from Portugal. Pathogens. 2022; 11(11):1242. https://doi.org/10.3390/pathogens11111242
Chicago/Turabian StyleMega, João, Sérgio Santos-Silva, Ana Loureiro, Josman D. Palmeira, Rita T. Torres, Antonio Rivero-Juarez, David Carmena, and João Mesquita. 2022. "Balantioides coli Fecal Excretion in Hunted Wild Cervids (Cervus elaphus and Dama dama) from Portugal" Pathogens 11, no. 11: 1242. https://doi.org/10.3390/pathogens11111242
APA StyleMega, J., Santos-Silva, S., Loureiro, A., Palmeira, J. D., Torres, R. T., Rivero-Juarez, A., Carmena, D., & Mesquita, J. (2022). Balantioides coli Fecal Excretion in Hunted Wild Cervids (Cervus elaphus and Dama dama) from Portugal. Pathogens, 11(11), 1242. https://doi.org/10.3390/pathogens11111242