
First Record of Colletotrichum anthrisci Causing Anthracnose on Avocado Fruits in Chile

 , , , and
, , , and
Abstract
1. Introduction
2. Results
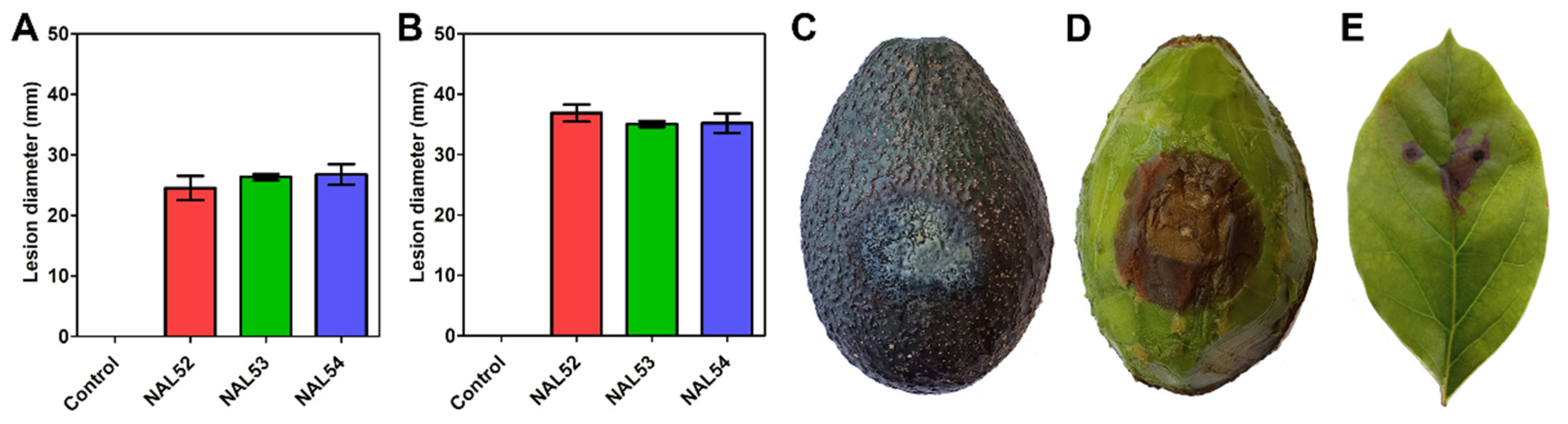
2.1. Symptomatology
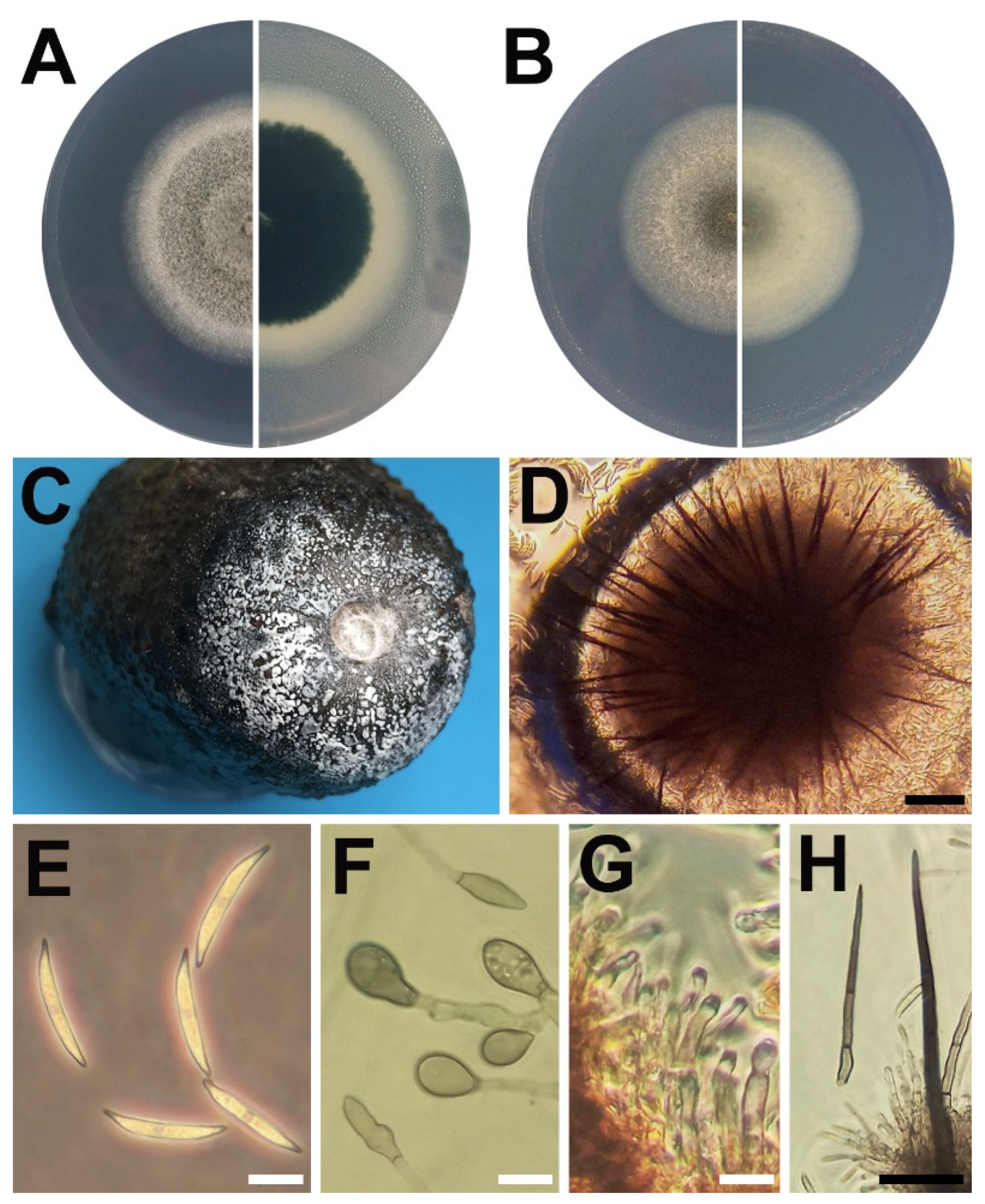
2.2. Morphological Analysis
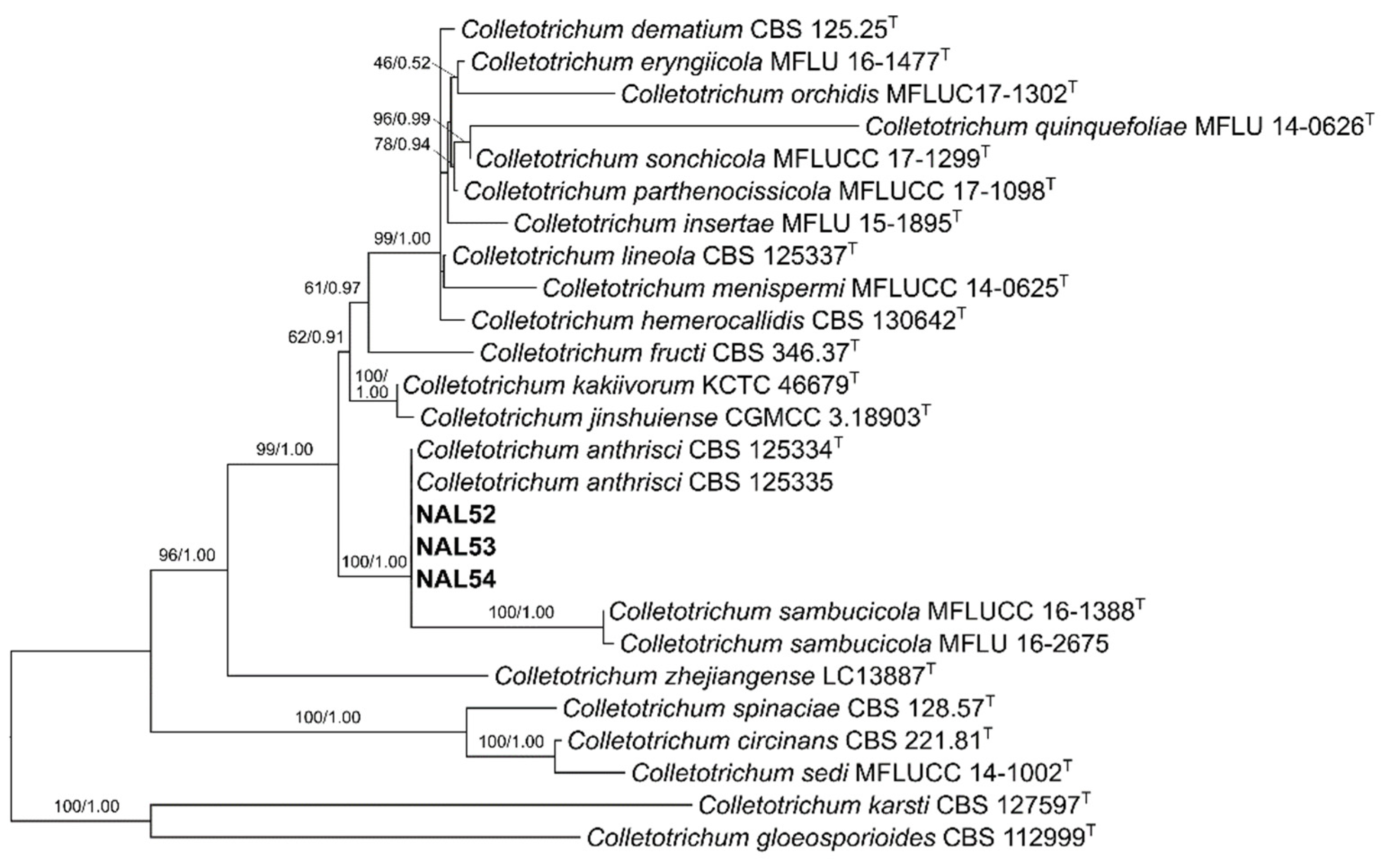
2.3. Phylogenetic Analysis
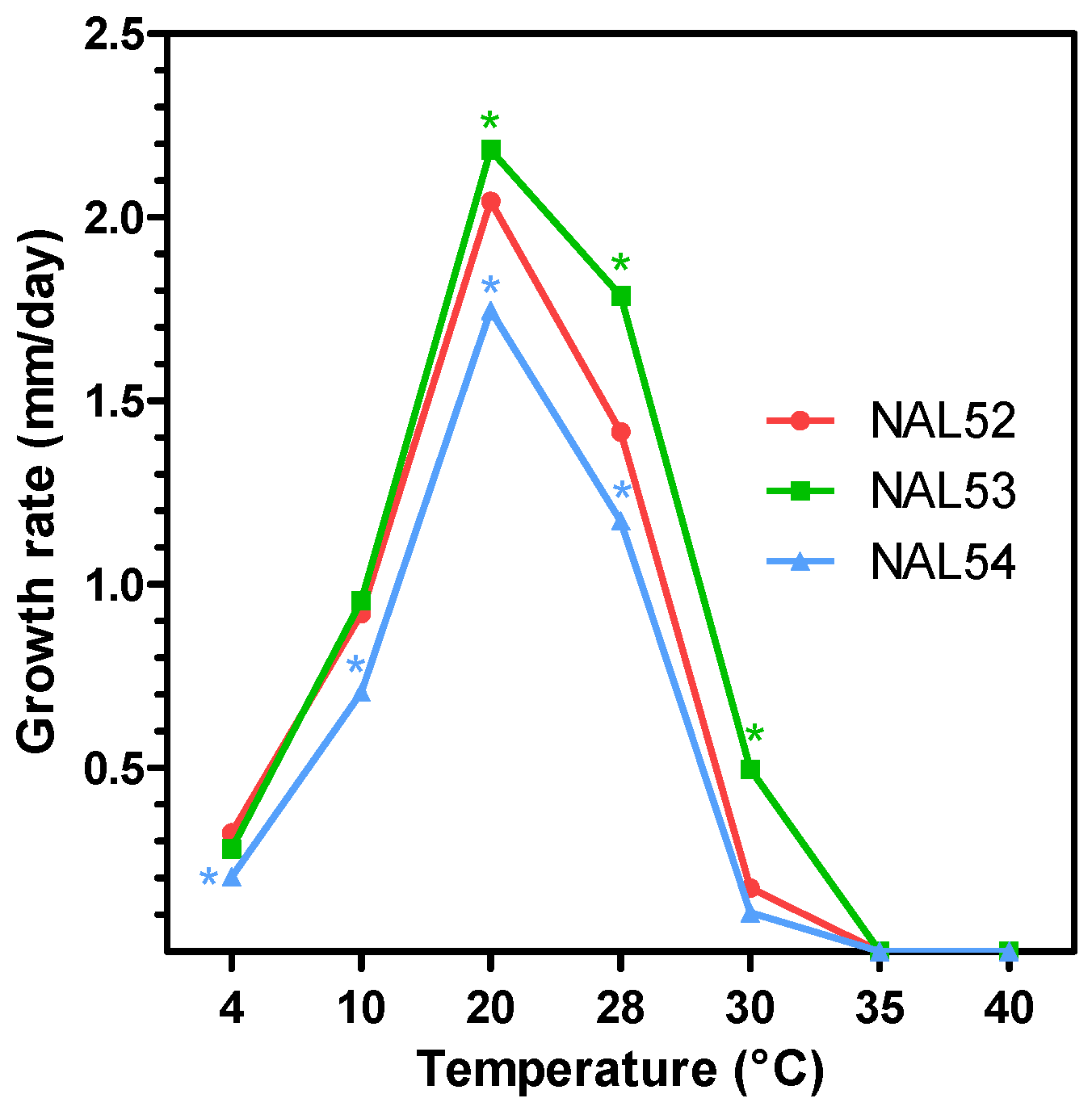
2.4. Effect of Temperature on Mycelial Growth
2.5. Pathogenicity Test
3. Discussion
4. Materials and Methods
4.1. Fruit Sampling
4.2. Fungal Isolation
4.3. Morphological Analysis
4.4. DNA Extraction, Amplification of DNA Barcodes, and Sequencing
4.5. Phylogenetic Analysis
4.6. Effect of Temperature on Mycelial Growth
4.7. Pathogenicity Tests
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, C.E., Jr. Archaeological evidence for selection in avocado. Econ. Bot. 1966, 20, 169–175. [Google Scholar] [CrossRef]
- Yahia, E.M. Avocado. In Crop Post-Harvest: Science and Technology-Perishables, 1st ed.; Rees, D., Farrell, G., Orchard, J., Eds.; Wiley-Blackwell: Chichester, UK, 2012; Volume 3, pp. 159–186. [Google Scholar] [CrossRef]
- Dreher, M.L.; Davenport, A.J. Hass avocado composition and potential health effects. Crit. Rev. Food Sci. Nutr. 2013, 53, 738–750. [Google Scholar] [CrossRef] [PubMed]
- Araújo, R.G.; Rodriguez-Jasso, R.M.; Ruiz, H.A.; Pintado, M.M.E.; Aguilar, C.N. Avocado by-products: Nutritional and functional properties. Trends Food Sci. Technol. 2018, 80, 51–60. [Google Scholar] [CrossRef]
- ODEPA. Boletín de la Fruta, Mayo 2022. Available online: https://apps.odepa.gob.cl/powerBI/boletin_fruta_mayo_2022.html (accessed on 9 August 2022).
- ODEPA-CIREN. Catastro Frutícola. Available online: https://www.odepa.gob.cl/estadisticas-del-sector/catastros-fruticolas/catastro-fruticola-ciren-odepa (accessed on 9 August 2022).
- Lemus, G.; Ferreyra, R.; Gil, P.; Sepúlveda, P.; Maldonado, P.; Toledo, C.; Barrera, C.; Celedón de Andraca, J.M. El Cultivo Del Palto, 3rd ed.; Instituto de Investigaciones Agropecuarias: Santiago, Chile, 2010; p. 39. [Google Scholar]
- Valencia, A.L.; Gil, P.M.; Latorre, B.A.; Rosales, I.M. Characterization and pathogenicity of Botryosphaeriaceae species obtained from avocado trees with branch canker and dieback and from avocado fruit with stem end rot in Chile. Plant Dis. 2019, 103, 996–1005. [Google Scholar] [CrossRef] [PubMed]
- Prusky, D.; Plumbley, R.A. Quiescent infections of Colletotrichum in tropical and subtropical fruits. In Colletotrichum: Biology, Pathology and Control, 1st ed.; Bailey, J.A., Jeger, M.J., Eds.; CABI: Wallingford, UK, 1992; pp. 289–307. [Google Scholar]
- Nelson, S.C. Anthracnose of avocado. Plant Dis. 2008, 58, 1–6. [Google Scholar]
- Cannon, P.F.; Damm, U.; Johnston, P.R.; Weir, B.S. Colletotrichum-current status and future directions. Stud. Mycol. 2012, 73, 181–213. [Google Scholar] [CrossRef]
- Hunupolagama, D.M.; Wijesundera, R.L.C.; Chandrasekharan, N.V.; Wijesundera, W.S.S.; Kathriarachchi, H.S.; Fernando, T.H.P.S. Characterization of Colletotrichum isolates causing avocado anthracnose and first report of C. gigasporum infecting avocado in Sri Lanka. Plant Pathol. Quar. 2015, 5, 132–143. [Google Scholar] [CrossRef]
- Sharma, G.; Maymon, M.; Freeman, S. Epidemiology, pathology and identification of Colletotrichum including a novel species associated with avocado (Persea americana) anthracnose in Israel. Sci. Rep. 2017, 7, 15839. [Google Scholar] [CrossRef]
- Giblin, F.R.; Tan, Y.P.; Mitchell, R.; Coates, L.M.; Irwin, J.A.G.; Shivas, R.G. Colletotrichum species associated with pre-and postharvest diseases of avocado and mango in eastern Australia. Australas. Plant Pathol. 2018, 47, 269–276. [Google Scholar] [CrossRef]
- Fuentes-Aragón, D.; Silva-Rojas, H.V.; Guarnaccia, V.; Mora-Aguilera, J.A.; Aranda-Ocampo, S.; Bautista-Martínez, N.; Téliz-Ortíz, D. Colletotrichum species causing anthracnose on avocado fruit in Mexico: Current status. Plant Pathol. 2020, 69, 1513–1528. [Google Scholar] [CrossRef]
- Soares, M.G.O.; Alves, E.; Silveira, A.L.; Pereira, F.D.; Guimarães, S.S.C. Colletotrichum siamense is the main aetiological agent of anthracnose of avocado in south-eastern Brazil. Plant Pathol. 2020, 70, 154–166. [Google Scholar] [CrossRef]
- Hofer, K.M.; Braithwaite, M.; Braithwaite, L.J.; Sorensen, S.; Siebert, B.; Pather, V.; Goudie, L.; Williamson, L.; Alexander, B.J.R.; Toome-Heller, M. First report of Colletotrichum fructicola, C. perseae, and C. siamense causing anthracnose disease of avocado (Persea americana) in New Zealand. Plant Dis. 2021, 105, 1564. [Google Scholar] [CrossRef] [PubMed]
- Morales, A.; Berger, H.; Luza, J. Identificación de hongos causantes de pudriciones en almacenaje refrigerado de paltas (Persea americana Mill.) Fuerte y Negra La Cruz. Inv. Agríc. 1979, 5, 1–4. [Google Scholar]
- Damm, U.; Woudenberg, J.H.C.; Cannon, P.F.; Crous, P.W. Colletotrichum species with curved conidia from herbaceous hosts. Fungal Divers. 2009, 39, 45–87. [Google Scholar]
- Tibpromma, S.; Hyde, K.D.; Jeewon, R.; Maharachchikumbura, S.S.N.; Liu, J.K.; Bhat, D.J.; Gareth Jones, E.B.; McKenzie, E.H.C.; Camporesi, E.; Bulgakov, T.S.; et al. Fungal diversity notes 491–602: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2017, 83, 1–261. [Google Scholar] [CrossRef]
- Buyck, B.; Duhem, B.; Das, K.; Jayawardena, R.S.; Niveiro, N.; Pereira, O.L.; Prasher, I.B.; Adhikari, S.; Albertó, E.O.; Bulgakov, T.S.; et al. Fungal biodiversity profiles 21-30. Cryptogam. Mycol. 2017, 38, 101–146. [Google Scholar] [CrossRef]
- Jayawardena, R.S.; Hyde, K.D.; Damm, U.; Cai, L.; Liu, M.; Li, X.H.; Zhang, W.; Zhao, W.S.; Yan, J.Y. Notes on currently accepted species of Colletotrichum. Mycosphere 2016, 7, 1192–1260. [Google Scholar] [CrossRef]
- Jayawardena, R.S.; Camporesi, E.; Elgorban, A.M.; Bahkali, A.H.; Yan, J.; Hyde, K.D. A new species of Colletotrichum from Sonchus sp. in Italy. Phytotaxa 2017, 314, 55–63. [Google Scholar] [CrossRef]
- Lee, S.Y.; Jung, H.Y. Colletotrichum kakivorum sp. nov., a new leaf spot pathogen of persimmon in Korea. Mycol. Prog. 2018, 17, 1113–1121. [Google Scholar] [CrossRef]
- Fu, M.; Crous, P.W.; Bai, Q.; Zhang, P.F.; Xiang, J.; Guo, Y.S.; Zhao, F.F.; Yang, M.M.; Hong, N.; Xu, W.X.; et al. Colletotrichum species associated with anthracnose of Pyrus spp. in China. Persoonia 2019, 42, 1–35. [Google Scholar] [CrossRef]
- Hyde, K.D.; Norphanphoun, C.; Maharachchikumbura, S.S.N.; Bhat, D.J.; Jones, E.B.G.; Bundhun, D.; Chen, Y.J.; Bao, D.F.; Boonmee, S.; Calabon, M.S.; et al. Refined families of Sordariomycetes. Mycosphere 2020, 11, 305–1059. [Google Scholar] [CrossRef]
- Jayawardena, R.S.; Hyde, K.D.; Chen, Y.J.; Papp, V.; Palla, B.; Papp, D.; Bhunjun, C.S.; Hurdeal, V.G.; Senwanna, C.; Manawasinghe, I.S.; et al. One stop shop IV: Taxonomic update with molecular phylogeny for important phytopathogenic genera: 76–100 (2020). Fungal Divers. 2020, 103, 87–218. [Google Scholar] [CrossRef]
- Liu, F.; Ma, Z.Y.; Hou, L.W.; Diao, Y.Z.; Wu, W.P.; Damm, U.; Song, S.; Cai, L. Updating species diversity of Colletotrichum, with a phylogenomic overview. Stud. Mycol. 2022, 101, 1–86. [Google Scholar] [CrossRef] [PubMed]
- Konno, M.; Iwamoto, S.; Seiwa, K. Specialization of a fungal pathogen on host tree species in a cross-inoculation experiment. J. Ecol. 2011, 99, 1394–1401. [Google Scholar] [CrossRef]
- Jayawardena, R.S.; Hyde, K.D.; Jeewon, R.; Li, X.H.; Liu, M.; Yan, J.Y. Mycosphere Essay 6: Why is it important to correctly name Colletotrichum species? Mycosphere 2016, 7, 1076–1092. [Google Scholar] [CrossRef]
- Shivaprakash, M.R.; Baghela, A.; Chakrabarti, A. Colletotrichum. In Molecular Detection of Human Fungal Pathogens, 1st ed.; Liu, D., Ed.; CRC Press: New York, NY, USA, 2011; pp. 401–409. [Google Scholar]
- Natarajan, S.V.; Rekha, N.S.; Sharda, R.D.; Mahalingam, N. Colletotrichum keratitis: A rare but definite clinical entity. J. Clin. Diagn. Res. 2013, 7, 1430–1433. [Google Scholar] [CrossRef] [PubMed]
- Navalpotro Rodríguez, D.; Martinez-Macias, O.; Domínguez-Márquez, V.; Burgos Teruel, A. Severe corneal infection by Colletotrichum gloeosporioides in a farmer. Med. Clin. 2014, 142, 138–139. [Google Scholar] [CrossRef]
- Lin, L.-Y.; Yang, C.-C.; Wan, J.-Y.; Chang, T.S.; Lee, J.Y.-Y. Cutaneous infection caused by plant pathogen Colletotrichum gloeosporioides. JAMA Dermatol. 2015, 151, 1383–1384. [Google Scholar] [CrossRef]
- Cai, L.; Hyde, K.D.; Taylor, P.W.J.; Weir, B.; Waller, J.M.; Abang, M.M.; Zang, J.C.; Yang, Y.L.; Phoulivong, S.; Prihastuti, Z.Y.; et al. A polyphasic approach for studying Colletotrichum. Fungal Divers. 2009, 39, 183–204. [Google Scholar]
- Han, Y.C.; Zeng, X.G.; Xiang, F.Y.; Ren, L.; Chen, F.Y.; Gu, Y.C. Distribution and characteristics of Colletotrichum spp. associated with anthracnose of strawberry in Hubei, China. Plant Dis. 2016, 100, 996–1006. [Google Scholar] [CrossRef]
- Thomas, G.J.; Sweetingham, M.W.; Yang, H.A.; Speijers, J. Effect of temperature on growth of Colletotrichum lupini and on anthracnose infection and resistance in lupines. Australas. Plant Pathol. 2008, 37, 35–39. [Google Scholar] [CrossRef]
- Lu, Q.; Wang, Y.; Li, N.; Yang, Y.; Wang, X. Differences in the characteristics and pathogenicity of Colletotrichum camelliae and C. fructicola isolated from the tea plant (Camellia (L.) O. Kuntze). Front. Microbiol. 2018, 9, 3060. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Liu, Z.; Zhang, W. First report of anthracnose disease on avocado (Persea americana) caused by Colletotrichum fructicola in China. Plant Dis. 2022, 106, 2529. [Google Scholar] [CrossRef] [PubMed]
- Henríquez-Sáez, J.L. (University of Chile, Santiago, Chile). Personal communication, 2022.
- Nirenberg, H.I. Untersuchungen über die morphologische und biologische Differenzierung in der Fusarium-Sektion Liseola. Mitt. Biol. Bundesanst. Land- Forstwirtsch. 1976, 169, 1–117. [Google Scholar]
- Loper, J.E.; Henkels, M.D.; Roberts, R.G.; Grove, G.G.; Willett, M.J.; Smith, T.J. Evaluation of streptomycin, oxytetracycline, and copper resistance of Erwinia amylovora isolated from pear orchards in Washington State. Plant Dis. 1991, 75, 287–290. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Shinsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Guerber, J.C.; Liu, B.; Correll, J.C.; Johnston, P.R. Characterization of diversity in Colletotrichum acutatum sensu lato by sequence analysis of two gene introns, mtDNA and intron RFLPs, and mating compatibility. Mycologia 2003, 95, 872–895. [Google Scholar] [CrossRef]
- Crous, P.W.; Groenewald, J.Z.; Risède, J.M.; Simoneau, P.; Hywel-Jones, N.L. Calonectria species and their Cylindrocladium anamorphs: Species with sphaeropedunculate vesicles. Stud. Mycol. 2004, 50, 415–430. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef]
- Huang, X.; Madan, A. CAP3: A DNA sequence assembly program. Genome Res. 1999, 9, 868–877. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Bourret, T.B.; Choudhury, R.A.; Mehl, H.K.; Blomquist, C.L.; McRoberts, N.; Rizzo, D.M. Multiple origins of downy mildews and mitonuclear discordance within the paraphyletic genus Phytophthora. PLoS ONE 2018, 13, e0192502. [Google Scholar] [CrossRef]
- Stöver, B.C.; Müller, K.F. TreeGraph 2: Combining and visualizing evidence from different phylogenetic analyses. BMC Bioinform. 2010, 11, 7. [Google Scholar] [CrossRef]
- Damm, U.; Cannon, P.F.; Woudenberg, J.H.C.; Johnston, P.R.; Weir, B.S.; Tan, Y.P.; Shivas, R.G.; Crous, P.W. The Colletotrichum boninense species complex. Stud. Mycol. 2012, 73, 1–36. [Google Scholar] [CrossRef]
- Li, G.J.; Hyde, K.D.; Zhao, R.L.; Hongsanan, S.; Abdel-Aziz, F.A.; Abdel-Wahab, M.A.; Alvarado, P.; Alves-Silva, G.; Ammirati, J.F.; Ariyawansa, H.A.; et al. Fungal diversity notes 253–366: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2016, 78, 1–237. [Google Scholar] [CrossRef]
- Liu, J.K.; Hyde, K.D.; Jones, E.B.G.; Ariyawansa, H.A.; Bhat, D.J.; Boonmee, S.; Maharachchikumbura, S.S.N.; McKenzie, E.H.C.; Phookamsak, R.; Phukhamsakda, C.; et al. Fungal diversity notes 1–110: Taxonomic and phylogenetic contributions to fungal species. Fungal Divers. 2015, 72, 1–197. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, Z.; Cai, L.; Hyde, K.D. New species and notes of Colletotrichum on daylilies (Hemerocallis spp.). Trop. Plant Pathol. 2012, 37, 165–174. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Character | NAL Isolates 1 | C. anthrisci 2 | C. sambucicola 3 |
|---|---|---|---|
| Conidia length (μm) | 25.8 ± 1.4 | 25 ± 1.5 | 17.7 |
| Conidia width (μm) | 4.3 ± 0.6 | 3.5 ± 0.2 | 3.8 |
| Conidia L/W ratio | 6 | 7.3 | 4.7 |
| Setae septum (n) | 2–5 | 2–4 | 1–6 |
| Setae length (μm) | 112.5–480 | 90–350 | 110–145 |
| Setae width (μm) | 7.5–14.5 | 6–18 | 6.1–8.5 |
| Setae L/W ratio | 26.9 | 18.3 | 17.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bustamante, M.I.; Osorio-Navarro, C.; Fernández, Y.; Bourret, T.B.; Zamorano, A.; Henríquez-Sáez, J.L. First Record of Colletotrichum anthrisci Causing Anthracnose on Avocado Fruits in Chile. Pathogens 2022, 11, 1204. https://doi.org/10.3390/pathogens11101204
Bustamante MI, Osorio-Navarro C, Fernández Y, Bourret TB, Zamorano A, Henríquez-Sáez JL. First Record of Colletotrichum anthrisci Causing Anthracnose on Avocado Fruits in Chile. Pathogens. 2022; 11(10):1204. https://doi.org/10.3390/pathogens11101204
Chicago/Turabian StyleBustamante, Marcelo I., Claudio Osorio-Navarro, Ysadora Fernández, Tyler B. Bourret, Alan Zamorano, and José Luis Henríquez-Sáez. 2022. "First Record of Colletotrichum anthrisci Causing Anthracnose on Avocado Fruits in Chile" Pathogens 11, no. 10: 1204. https://doi.org/10.3390/pathogens11101204
APA StyleBustamante, M. I., Osorio-Navarro, C., Fernández, Y., Bourret, T. B., Zamorano, A., & Henríquez-Sáez, J. L. (2022). First Record of Colletotrichum anthrisci Causing Anthracnose on Avocado Fruits in Chile. Pathogens, 11(10), 1204. https://doi.org/10.3390/pathogens11101204