
Neofusicoccum mediterraneum Is Involved in a Twig and Branch Dieback of Olive Trees Observed in Salento (Apulia, Italy)


Abstract
:1. Introduction
2. Results
2.1. Natural Symptoms
2.2. Assessment of Fungi and Bacteria in Twigs and Branches Symptomatic Samples
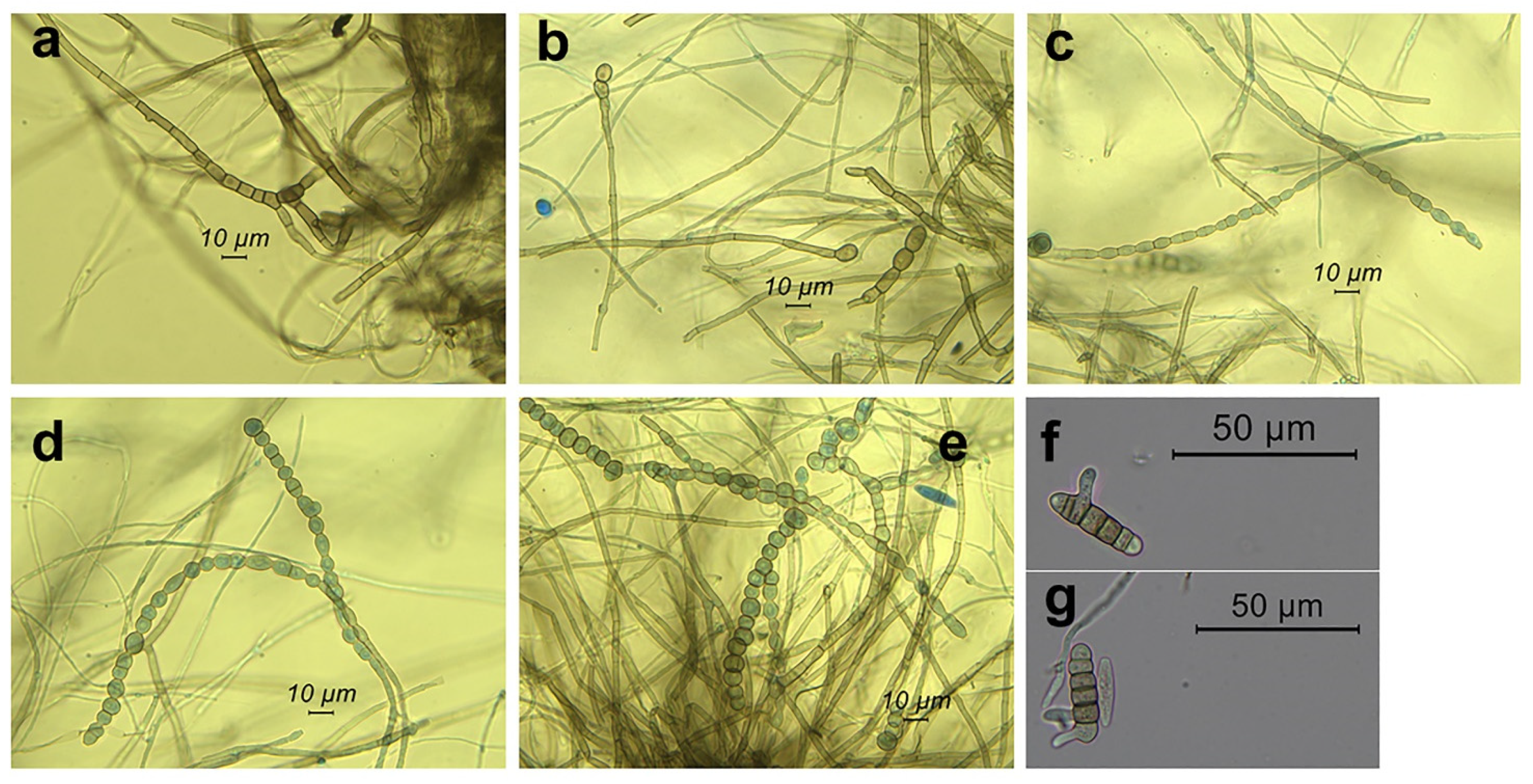
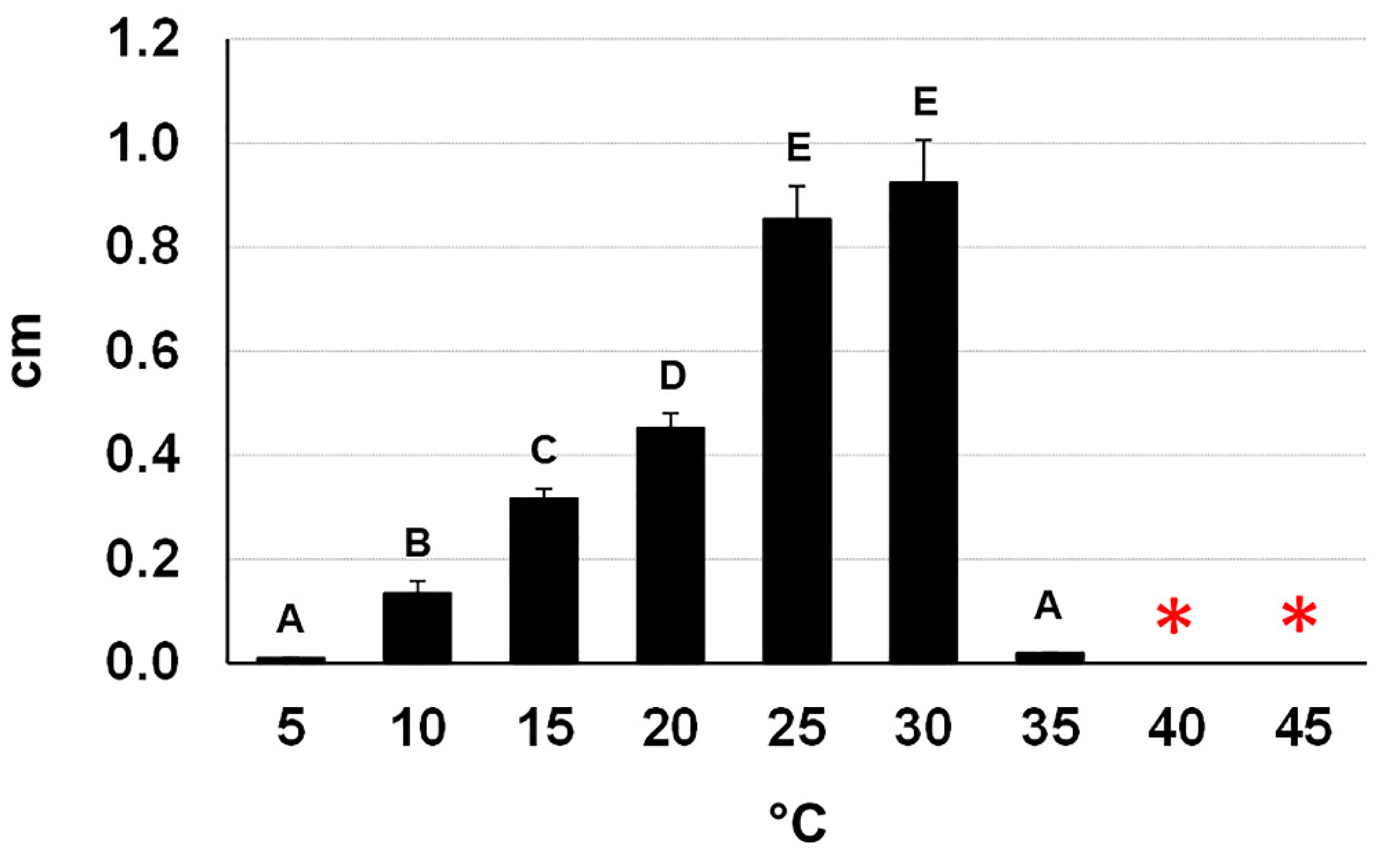
2.3. Morpho-Cultural Characterization of the Botryosphaeriaceous Isolate CREA-DC TPR OL 427
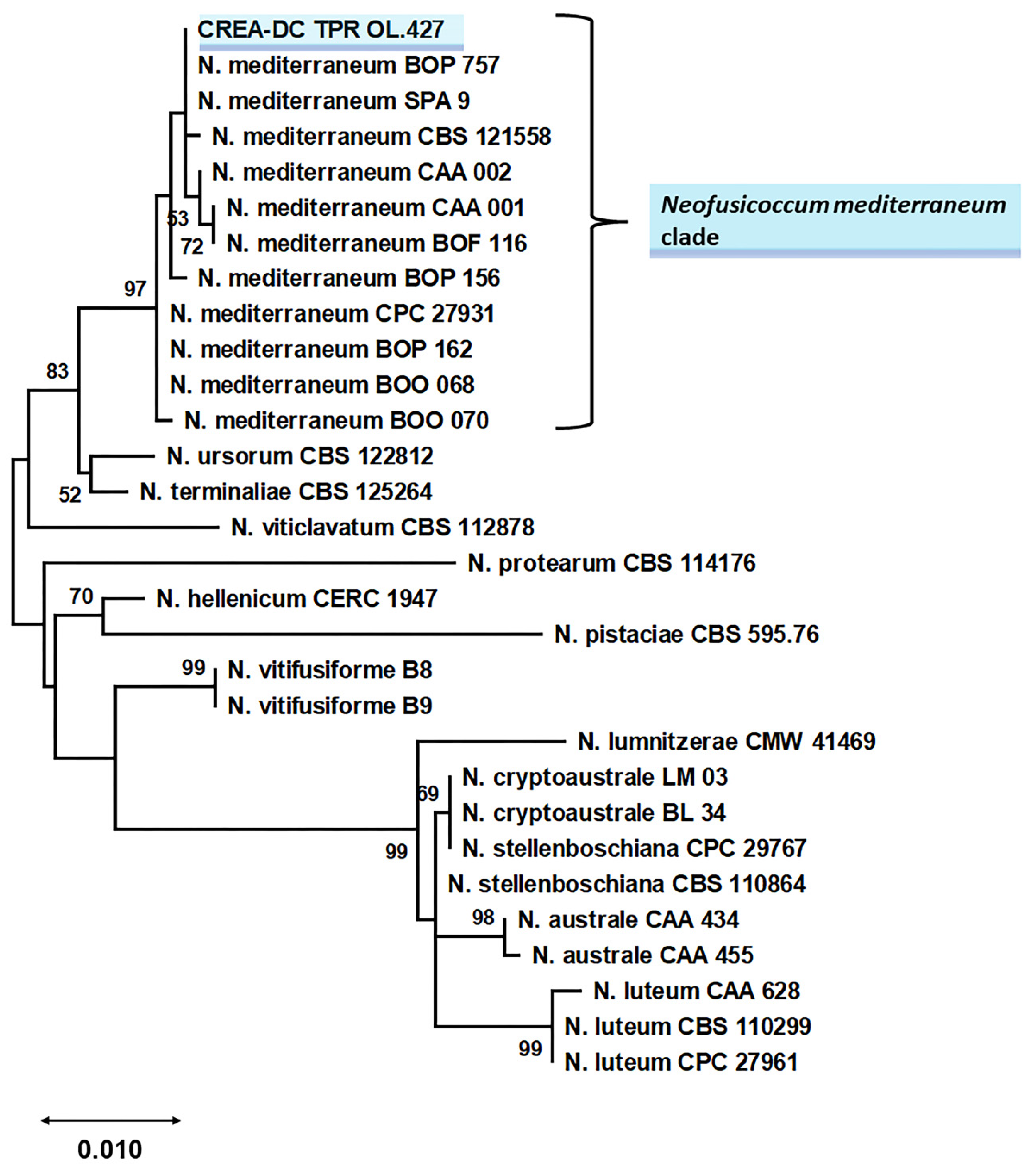
2.4. Sequencing and Phylogenetic Analysis for Species Identification of the Selected Isolate
2.5. Pathogenicity Tests
3. Discussion
4. Materials and Methods
4.1. Symptom Survey, Sample Collection, Isolations, and Detections
4.2. Morphological Features of the Selected Fungal Isolate and Its Cultural Characteristics
4.3. Sequencing and Phylogenetic Analysis for Species Identification
4.4. Pathogenicity Tests
4.5. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Besnard, G.; Khadari, B.; Navascués, M.; Fernández-Mazuecos, M.; El Bakkali, A.; Arrigo, N.; Baali-Cherif, D.; Brunini-Bronzini de Caraffa, V.; Santoni, S.; Vargas, P.; et al. The complex history of the olive tree: From late quaternary diversification of Mediterranean lineages to primary domestication in the northern Levant. Proc. Royal Soc. B 2013, 280, 1–7. [Google Scholar] [CrossRef] [Green Version]
- European Commission. Market Situation in the Olive Oil and Table Olives Sectors—Production, Committee for the Common Organisation of the Agricultural Markets—Arable Crops and Olive Oil—30 September 2021—Consumption and Ending Stocks. Available online: https://ec.europa.eu/info/sites/default/files/food-farming-fisheries/plants_and_plant_products/documents/market-situation-olive-oil-table-olives_en.pdf (accessed on 1 November 2021).
- Rigacci, S.; Stefani, M. Nutraceutical Properties of Olive Oil Polyphenols. An Itinerary from Cultured Cells through Animal Models to Humans. Int. J. Mol. Sci. 2016, 17, 843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Özcan, M.M.; Matthäus, B. A review: Benefit and bioactive properties of olive (Olea europaea L.) leaves. Eur. Food Res. Technol. 2017, 243, 89–99. [Google Scholar] [CrossRef]
- Lima-Cueto, F.J.; Blanco-Sepúlveda, R.; Gómez-Moreno, M.L. Soil erosion and environmental regulations in the european agrarian policy for olive groves (Olea europaea) of southern Spain. Agrociencia 2018, 52, 293–308. [Google Scholar]
- López-Escudero, F.J.; Mercado-Blanco, J. Verticillium wilt of olive: A case study to implement an integrated strategy to control a soil-borne pathogen. Plant. Soil 2011, 344, 1–50. [Google Scholar] [CrossRef] [Green Version]
- Jimenez-Diaz, R.M.; Cirulli, M.; Bubici, G.; del Mar Jimenez-Gasco, M.; Antoniou, P.P.; Tjamos, E.C. Verticillium wilt, a major threat to olive production: Current status and future prospects for its management. Plant Dis. 2012, 96, 304–329. [Google Scholar] [CrossRef] [Green Version]
- Hopkins, D.L. Xylella fastidiosa: Xylem-limited bacterial pathogens of plants. Annu. Rev. Phytopathol. 1989, 27, 271–290. [Google Scholar] [CrossRef]
- Frisullo, S.; Camele, I.; Agosteo, G.E.; Boscia, D.; Martelli, G.P. Brief historical account of olive leaf scorch (“brusca”) in the Salento peninsula of Italy and state-of-the-art of the olive quick decline syndrome. J. Plant Pathol. 2014, 96, 441–449. [Google Scholar] [CrossRef]
- Scortichini, M. Predisposing Factors for “Olive Quick Decline Syndrome” in Salento (Apulia, Italy). Agronomy 2020, 10, 1445. [Google Scholar] [CrossRef]
- Moral, J.; Muñoz-Díez, C.; González, N.; Trapero, A.; Michailides, T.J. Characterization and pathogenicity of Botryosphaeriaceae species collected from olive and other hosts in Spain and California. Phytopathology 2010, 100, 1340–1351. [Google Scholar] [CrossRef] [Green Version]
- Carlucci, A.; Raimondo, M.L.; Cibelli, F.; Phillips, A.J.; Lops, F. Pleurostomophora richardsiae, Neofusicoccum parvum and Phaeoacremonium aleophilum associated with a decline of olives in southern Italy. Phytopathol. Mediterr. 2013, 517–527. [Google Scholar] [CrossRef]
- Úrbez-Torres, J.R.; Peduto, F.; Vossen, P.M.; Krueger, W.H.; Gubler, W.D. Olive twig and branch dieback: Etiology, incidence, and distribution in California. Plant Dis. 2013, 97, 231–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlucci, A.; Lops, F.; Cibelli, F.; Raimondo, M.L. Phaeoacremonium species associated with olive wilt and decline in southern Italy. Eur. J. Plant Pathol. 2015, 141, 717–729. [Google Scholar] [CrossRef] [Green Version]
- Moral, J.; Agustí-Brisach, C.; Pérez-Rodríguez, M.; Xaviér, C.; Raya, M.C.; Rhouma, A.; Trapero, A. Identification of fungal species associated with branch dieback of olive and resistance of table cultivars to Neofusicoccum mediterraneum and Botryosphaeria dothidea. Plant Dis. 2017, 101, 306–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, A.J.L.; Alves, A.; Abdollahzadeh, J.; Slippers, B.; Wingfield, M.J.; Groenewald, J.Z.; Crous, P.W. The Botryosphaeriaceae: Genera and species known from culture. Stud. Mycol. 2013, 76, 51–167. [Google Scholar] [CrossRef] [Green Version]
- Lopes, A.; Phillips, A.J.L.; Alves, A. Mating type genes in the genus Neofusicoccum: Mating strategies and usefulness in species delimitation. Fungal Biol. 2017, 121, 394–404. [Google Scholar] [CrossRef]
- Bezerra, J.D.P.; Crous, P.W.; Aiello, D.; Gullino, M.L.; Polizzi, G.; Guarnaccia, V. Genetic Diversity and Pathogenicity of Botryosphaeriaceae Species Associated with Symptomatic Citrus Plants in Europe. Plants 2021, 10, 492. [Google Scholar] [CrossRef] [PubMed]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press Inc.: Oxford, UK, 2000; p. 333. [Google Scholar]
- Slippers, B.; Wingfield, M.J. Botryosphaeriaceae as endophytes and latent pathogens of woody plants: Diversity, ecology and impact. Fungal Biol. Rev. 2007, 21, 90–106. [Google Scholar] [CrossRef]
- Kaliterna, J.; Milicevic, T.; Ivic, D.; Bencic, D.; Mesic, A. First report of Diplodia seriata as causal agent of olive dieback in Croatia. Plant Dis. 2012, 96, 290. [Google Scholar] [CrossRef] [PubMed]
- Manca, D.; Bregant, C.; Maddau, L.; Pinna, C.; Montecchio, L.; Linaldeddu, B.T. First report of canker and dieback caused by Neofusicoccum parvum and Diplodia olivarum on oleaster in Italy. Ital. J. Mycol. 2020, 49, 85–91. [Google Scholar] [CrossRef]
- Romero, M.A.; Sánchez, M.E.; Trapero, A. First Report of Botryosphaeria ribis as a Branch Dieback Pathogen of Olive Trees in Spain. Plant Dis. 2007, 89, 208. [Google Scholar] [CrossRef] [PubMed]
- Lazzizera, C.; Frisullo, S.; Alves, A.; Phillips, A.J.L. Morphology, phylogeny and pathogenicity of Botryosphaeria and Neofusicoccum species associated with drupe rot of olives in southern Italy. Plant Path. 2008, 57, 948–956. [Google Scholar] [CrossRef]
- Zlatković, M.; Keča, N.; Wingfield, M.J.; Jami, F.; Slippers, B. Botryosphaeriaceae associated with the die-back of ornamental trees in the Western Balkans. Anton. Leeuw. Int. J. G. 2016, 109, 543–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deidda, A.; Buffa, F.; Linaldeddu, B.D.; Pinna, C.; Scanu, B.; Deiana, V.; Satta, A.; Franceschini, A.; Floris, I. Emerging pests and diseases threaten Eucalyptus camaldulensis plantations in Sardinia, Italy. iForest Biogeosci. For. 2016, 9, 883–891. [Google Scholar] [CrossRef]
- Billones-Baaijens, R.; Savocchia, S. A review of Botryosphaeriaceae species associated with grapevine trunk diseases in Australia and New Zealand. Australas. Plant Pathol. 2019, 48, 3–18. [Google Scholar] [CrossRef]
- Harper, S.J.; Ward, L.I.; Clover, G.R.G. Development of LAMP and real-time PCR methods for the rapid detection of Xylella fastidiosa for quarantine and field applications. Phytopathol. 2010, 100, 1282–1288. [Google Scholar] [CrossRef] [PubMed]
- EPPO PM 7/24 (4). Xylella fastidiosa. Bull. OEPP 2019, 49, 175–227. [CrossRef] [Green Version]
- Bilodeau, G.J.; Koike, S.T.; Uribe, P.; Martin, F.N. Development of an Assay for Rapid Detection and Quantification of Verticillium dahliae in Soil. Phytopathology 2012, 102, 331–342. [Google Scholar] [CrossRef] [Green Version]
- Scortichini, M.; Chen, J.; De Caroli, M.; Dalessandro, G.; Pucci, N.; Modesti, V.; L’Aurora, A.; Petriccione, M.; Zampella, L.; Mastrobuoni, F.; et al. A zinc, copper and citric acid biocomplex shows promise for control of Xylella fastidiosa subsp. pauca in olive trees in Apulia region (southern Italy). Phytopathol. Mediterr. 2018, 57, 48–72. [Google Scholar] [CrossRef]
- Crous, P.; Slippers, B.; Wingfield, M.; Rheeder, J.; Marasas, W.; Philips, A.; Alves, A.; Burgess, T.; Barber, P.; Groenewald, J. Phylogenetic lineages in the Botryosphaeriaceae. Stud. Mycol. 2006, 55, 235–253. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press Inc.: London, UK, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Alves, A.; Crous, P.W.; Correia, A.; Phillips, A.J.L. Morphological and molecular data reveal cryptic speciation in Lasiodiplodia theobromae. Fungal Divers. 2008, 28, 1–13. [Google Scholar]
- Glass, N.L.; Donaldson, G. Development of primer sets designed for use with PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Twig Trials | Replicates | Ø 1 (cm) | Bark Canker cm (SD) | Wood Discoloration cm (SD) | Wilting 2 |
|---|---|---|---|---|---|
| 17 May 2021 | 20 | 0.35–0.5 | 7.1 (1.8) | 16.9 (8.1) | + |
| 16 June 2021 | 20 | 0.45–0.7 | 7.0 (2.6) | 9.1 (2.4) | + |
| Stem trial | |||||
| 17 June 2021 | 10 | 1.35–1.55 | 12.2 (2.7) | 20.3 (8.7) | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brunetti, A.; Matere, A.; Lumia, V.; Pasciuta, V.; Fusco, V.; Sansone, D.; Marangi, P.; Cristella, N.; Faggioli, F.; Scortichini, M.; et al. Neofusicoccum mediterraneum Is Involved in a Twig and Branch Dieback of Olive Trees Observed in Salento (Apulia, Italy). Pathogens 2022, 11, 53. https://doi.org/10.3390/pathogens11010053
Brunetti A, Matere A, Lumia V, Pasciuta V, Fusco V, Sansone D, Marangi P, Cristella N, Faggioli F, Scortichini M, et al. Neofusicoccum mediterraneum Is Involved in a Twig and Branch Dieback of Olive Trees Observed in Salento (Apulia, Italy). Pathogens. 2022; 11(1):53. https://doi.org/10.3390/pathogens11010053
Chicago/Turabian StyleBrunetti, Angela, Antonio Matere, Valentina Lumia, Vittorio Pasciuta, Valeria Fusco, Domenico Sansone, Paolo Marangi, Nicola Cristella, Francesco Faggioli, Marco Scortichini, and et al. 2022. "Neofusicoccum mediterraneum Is Involved in a Twig and Branch Dieback of Olive Trees Observed in Salento (Apulia, Italy)" Pathogens 11, no. 1: 53. https://doi.org/10.3390/pathogens11010053
APA StyleBrunetti, A., Matere, A., Lumia, V., Pasciuta, V., Fusco, V., Sansone, D., Marangi, P., Cristella, N., Faggioli, F., Scortichini, M., & Pilotti, M. (2022). Neofusicoccum mediterraneum Is Involved in a Twig and Branch Dieback of Olive Trees Observed in Salento (Apulia, Italy). Pathogens, 11(1), 53. https://doi.org/10.3390/pathogens11010053