
Bluetongue and Epizootic Hemorrhagic Disease in the United States of America at the Wildlife–Livestock Interface

 ,
,  , , , , and
, , , , and
Abstract
1. Introduction

2. Animal Health Impact of BT and EHD
2.1. Etiology
2.2. Pathology
2.2.1. Bluetongue Disease (BT)
2.2.2. Epizootic Hemorrhagic Disease (EHD)
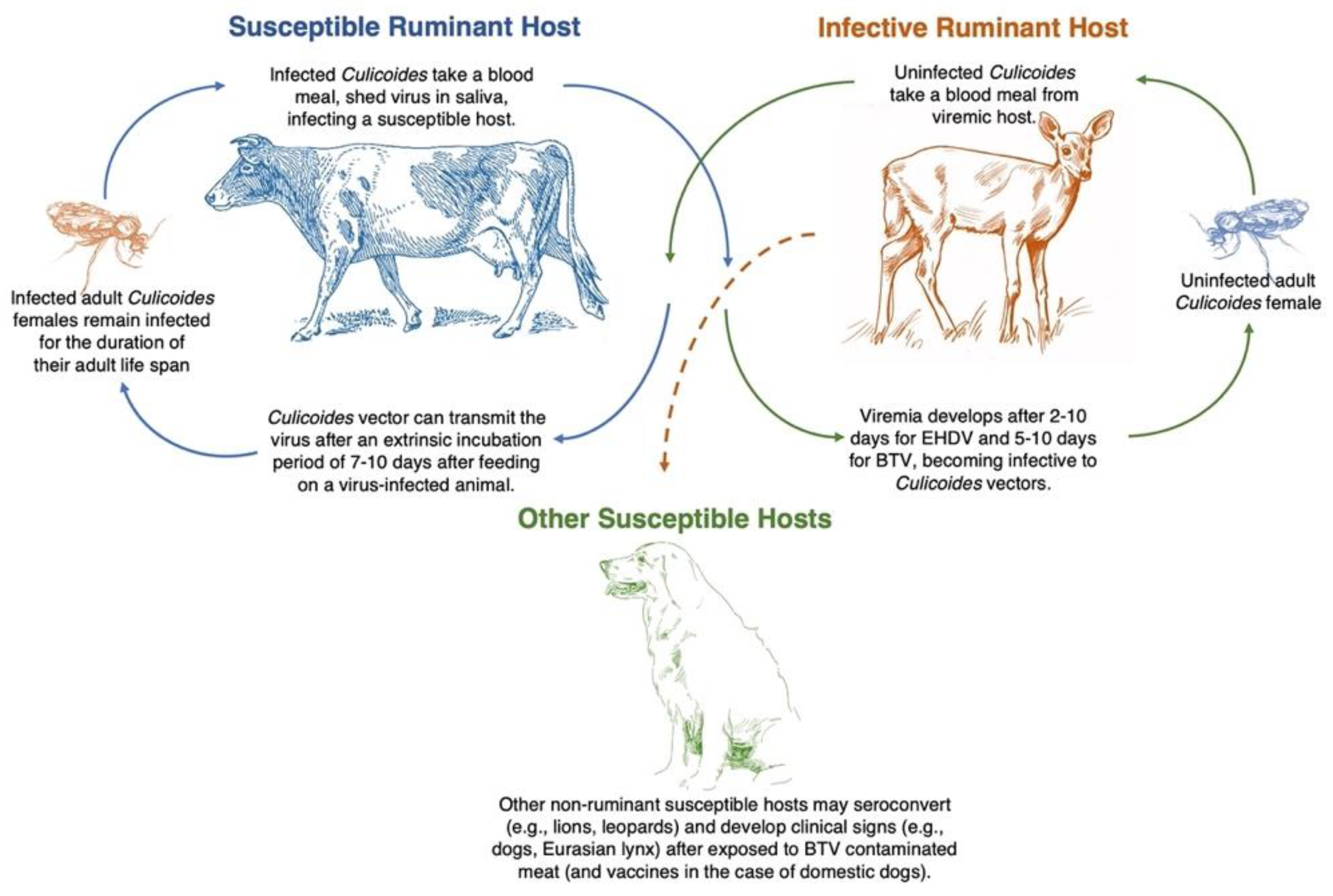
2.3. Biological Vector: Culicoides Midges
2.4. Structural and Molecular Biology
2.5. Impact
3. Prevalence and Distribution of BT and EHD in the USA
4. The Importance of Surveillance for Ruminant Populations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thomas, F.C.; Willis, N.; Ruckerbaker, G. Identification of the viruses involved in the 1971 outbreak of hemorrhagic disease in southeastern United States white- tailed deer. J. Wildl. Dis. 1974, 10, 187–189. [Google Scholar] [CrossRef]
- Gibbs, E.P.J.; Greiner, E.C. Bluetongue and epizootic hemorrhagic disease. In The Arboviruses: Epidemiology and Ecology; Monath, T.P., Ed.; CRC: Boca Raton, FL, USA, 1989; Volume 2, pp. 39–70. [Google Scholar]
- Annetti, K.L.; Rivera, N.A.; Andrews, J.E.; Mateus-Pinilla, N. Survey of haemosporidian parasites in resident and migrant game birds of Illinois. J. Fish Wildl. Manag. 2017, 8, 661–668. [Google Scholar] [CrossRef]
- Sick, F.; Beer, M.; Kampen, H.; Wernike, K. Culicoides biting midges—Underestimated vectors for arboviruses of public health and veterinary importance. Viruses 2019, 11, 376. [Google Scholar] [CrossRef]
- Maclachlan, N.J.; Zientara, S.; Wilson, W.C.; Richt, J.A.; Savini, G. Bluetongue and epizootic hemorrhagic disease viruses: Recent developments with these globally re-emerging arboviral infections of ruminants. Curr. Opin. Virol. 2019, 34, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Alexander, K.A.; Holekamp, K.; MacLachlan, N.J.; Frank, L.G.; Sawyer, M.; Osburn, B.I.; Mills, M.G.L.; Lerche, N.W.; Laurenson, M.K.; McNutt, J.W.; et al. Evidence of natural bluetongue virus infection among African carnivores. Am. J. Top Med. Hyg. 1994, 51, 568–576. [Google Scholar] [CrossRef]
- Alkhamis, M.A.; Aguilar-Vega, C.; Fountain-Jones, N.M.; Lin, K.; Perez, A.M.; Sánchez-Vizcaíno, J.M. Global emergence and evolutionary dynamics of bluetongue virus. Sci. Rep. 2020, 10, 21677. [Google Scholar] [CrossRef]
- Maclachlan, N.J.; Zientara, S.; Savini, G.; Daniels, P. Bluetongue. Rev. Sci. Tech. Off. Int. Epiz. 2015, 34, 329–340. [Google Scholar] [CrossRef] [PubMed]
- Savini, G.; Afonso, A.; Mellor, P.; Aradaib, I.A.O.; Yadin, H.; Sanaa, M.; Wilson, W.; Monaco, F.; Domingo, M. Epizootic hemorrhagic disease. Res. Vet. Sci. 2011, 91, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Howerth, E.W.; Stallknecht, D.E.; Kirkland, P.D. Bluetongue, epizootic haemorrhagic disease, and other orbivirus-related diseases. In Infectious Diseases of Wild Mammals; Williams, E.S., Barker, I.K., Eds.; Iowa State University Press: Ames, IA, USA, 2001; pp. 77–97. [Google Scholar]
- Spreull, J. Malarial catarrhal fever (bluetongue) of sheep in South Africa. J. Comp. Pathol. Ther. 1905, 18, 321–337. [Google Scholar] [CrossRef]
- Verwoerd, D.W. History of bluetongue research at Onderstepoort. J. Vet. Res. 2009, 76, 99–102. [Google Scholar]
- Hardy, W.T.; Price, D.A. Soremuzzle of sheep. J. Am. Vet. Med. Assoc. 1952, 120, 23–25. [Google Scholar]
- McKercher, D.G.; McGowan, B.L.A.I.N.E.; Howarth, J.A.; SAITO, J.K. A preliminary report on the isolation and identification of the bluetongue virus from sheep in California. J. Am. Vet. Med. Assoc. 1953, 122, 300–301. [Google Scholar]
- McVey, D.S.; Drolet, B.S.; Ruder, M.G.; Wilson, W.C.; Nayduch, D.; Pfannenstiel, R.; Cohnstaedt, L.W.; MacLachlan, N.J.; Gay, C.G. Orbiviruses: A north American perspective. Vector Borne Zoonotic Dis. 2015, 15, 335–338. [Google Scholar] [CrossRef]
- Shope, R.E.; MacNamara, L.G.; Mangold, R. A virus-induced epizootic hemorrhagic disease of the Virginia white-tailed deer (Odocoileus virginianus). J. Exp. Med. 1960, 111, 155–170. [Google Scholar] [CrossRef]
- MacLachlan, N.J.; Dubovi, E.J. Reoviridae. In Fenner’s Veterinary Virology, 5th ed.; Academic Press: Cambridge, MA, USA, 2017; pp. 299–317. [Google Scholar] [CrossRef]
- Mullen, G.R.; Murphree, C.S. Biting Midges (Ceratopogonidae). In Medical and Veterinary Entomology; Academic Press: Cambridge, MA, USA, 2019; pp. 213–236. [Google Scholar]
- Mertens, P.P.C.; Attoui, H.; Mellor, P.S. Orbiviruses. In Encyclopedia of Virology; Academic Press: Cambridge, MA, USA, 2008; pp. 454–465. [Google Scholar]
- Ruder, M.G.; Lysyk, T.J.; Stallknecht, D.E.; Foil, L.D.; Johnson, D.J.; Chase, C.C.; Dargatz, D.A.; Gibbs, E.P.J. Transmission and epidemiology of bluetongue and epizootic hemorrhagic disease in North America: Current perspectives, research gaps, and future directions. Vector Borne Zoonotic Dis. 2015, 15, 348–363. [Google Scholar] [CrossRef] [PubMed]
- Ruder, M.G.; Johnson, D.; Ostlund, E.; Allison, A.B.; Kienzle, C.; Phillips, J.E.; Poulson, R.L.; Stallknecht, D.E. The first 10 years (2006–2015) of epizootic hemorrhagic disease virus serotype 6 in the USA. J. Wildl. Dis. 2017, 53, 901–905. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Acevedo, A.M.; Hinojosa, Y.; Relova, D.; Perera, C.L. Bluetongue virus: A known virus, a current threat. Rev. Salud Ani. 2016, 38, 52–59. [Google Scholar]
- Tabachnick, W.; Chelsea, S.; Connelly, C. “Bluetongue”. EDIS. 2008. Available online: https://journals.flvc.org/edis/article/view/117291 (accessed on 20 July 2021).
- OIE. Bluetongue. Technical Disease Card. 2013. Available online: https://www.oie.int/fileadmin/Home/eng/Animal_Health_in_the_World/docs/pdf/Disease_cards/BLUETONGUE.pdf (accessed on 20 July 2021).
- OIE. Epizootic Hemorrhagic Disease. Technical Disease Card. 2019. Available online: https://www.oie.int/fileadmin/Home/eng/Animal_Health_in_the_World/docs/pdf/Disease_cards/EPIZOOTIC_HEAMORRHAGIC_DISEASE.pdf (accessed on 20 July 2021).
- George, T.S.; Standfast, H.A.; Cybinski, D.H.; Dyce, A.L.; Muller, M.J.; Doherty, R.L.; Carley, J.G.; FRAZIER, C.L. The isolation of a bluetongue virus from Culicoides collected in the Northern Territory of Australia. Aust. Vet. J. 1978, 54, 153–154. [Google Scholar] [CrossRef] [PubMed]
- Koltsov, A.; Tsybanov, S.; Gogin, A.; Kolbasov, D.; Koltsova, G. Identification and characterization of Bluetongue virus serotype 14 in Russia. Front. Vet. Sci. 2020, 7, 26. [Google Scholar] [CrossRef]
- Balasuriya, U.B.; Nadler, S.A.; Wilson, W.C.; Pritchard, L.I.; Smythe, A.B.; Savini, G.; Monaco, F.; De Santis, P.; Zhang, N.; Tabachnick, W.J.; et al. The NS3 proteins of global strains of bluetongue virus evolve into regional topotypes through negative (purifying) selection. Vet. Microbiol. 2008, 126, 91–100. [Google Scholar] [CrossRef]
- Nomikou, K.; Hughes, J.; Wash, R.; Kellam, P.; Breard, E.; Zientara, S.; Palmarini, M.; Biek, R.; Mertens, P.P.C. Widespread reassortment shapes the evolution and epidemiology of bluetongue virus following European invasion. PLoS Pathog. 2015, 11, e1005056. [Google Scholar] [CrossRef] [PubMed]
- Wilson, W.C.; Gaudreault, N.N.; Jasperson, D.C.; Johnson, D.J.; Ostlund, E.N.; Chase, C.L.; Ruder, M.G.; Stallknecht, D.E. Molecular evolution of American field strains of Bluetongue and Epizootic haemorrhagic disease viruses. Vet. Ital. 2015, 51, 269–273. [Google Scholar] [PubMed]
- Wilson, W.C.; Ruder, M.G.; Jasperson, D.; Smith, T.P.; Naraghi-Arani, P.; Lenhoff, R.; Stallknecht, D.E.; Valdiva-Granda, W.A.; Sheoran, D. Molecular evolution of epizootic hemorrhagic disease viruses in North America based on historical isolates using motif fingerprints. Virus Genes 2016, 52, 495–508. [Google Scholar] [CrossRef] [PubMed]
- Schulz, C.; Bréard, E.; Sailleau, C.; Jenckel, M.; Viarouge, C.; Vitour, D.; Palmarini, M.; Gallois, M.; Höper, D.; Hoffmann, B.; et al. Bluetongue virus serotype 27: Detection and characterization of two novel variants in Corsica, France. J. Gen. Virol. 2016, 97, 2073–2083. [Google Scholar] [CrossRef]
- Mayo, C.; McDermott, E.; Kopanke, J.; Stenglein, M.; Lee, J.; Mathiason, C.; Carpenter, M.; Reed, K.; Perkins, T.A. Ecological dynamics impacting bluetongue virus transmission in North America. Front. Vet. Sci. 2020, 7, 186. [Google Scholar] [CrossRef]
- Belbis, G.; Zientara, S.; Bre´ard, E.; Sailleau, C.; Caignard, G.; Vitour, D.; Attoui, H. Bluetongue virus: From BTV-1 to BTV-27. In Advances in Virus Research; Beer, M., Hoper, D., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 161–197. [Google Scholar]
- Bumbarov, V.; Golender, N.; Jenckel, M.; Wernike, K.; Beer, M.; Khinich, E.; Zalesky, O.; Erster, O. Characterization of bluetongue virus serotype 28. Transbound. Emerg. Dis. 2020, 67, 171–182. [Google Scholar] [CrossRef]
- Ries, C.; Vögtlin, A.; Hüssy, D.; Jandt, T.; Gobet, H.; Hilbe, M.; Burgener, C.; Schweizer, L.; Häfliger-Speiser, S.; Beer, M.; et al. Putative Novel Atypical BTV Serotype ‘36’Identified in Small Ruminants in Switzerland. Viruses 2021, 13, 721. [Google Scholar] [CrossRef]
- Stallknecht, D. Working Together: The 40th Anniversary of the National Hemorrhagic Disease Survey. In SCWDS BRIEFS; The University of Georgia: Athens, GA, USA, 2020; Volume 36, pp. 1–3. Available online: https://vet.uga.edu/wp-content/uploads/2020/11/2020-October-SCWDS-Briefs-Newsletter.pdf (accessed on 20 July 2021).
- Stallknecht, D.E.; Allison, A.B.; Park, A.W.; Phillips, J.E.; Goekjian, V.H.; Nettles, V.F.; Fischer, J.R. Apparent increase of reported hemorrhagic disease in the midwestern and northeastern USA. J. Wildl. Dis. 2015, 51, 348–361. [Google Scholar] [CrossRef] [PubMed]
- USDA. Reportable Diseases in the United States—2018 Annual Report. Available online: https://www.aphis.usda.gov/aphis/ourfocus/animalhealth/monitoring-and-surveillance/sa_nahss/status-reportable-disease-us (accessed on 20 July 2021).
- More, S.; Bicout, D.; Bøtner, A.; Butterworth, A.; Depner, K.; Edwards, S.; Garin-Bastuji, B.; Good, M.; Gortázar Schmidt, C.; Michel, V. Assessment of listing and categorisation of animal diseases within the framework of the Animal Health Law (Regulation (EU) No 2016/429): Bluetongue. EFSA J. 2017, 15, 8. [Google Scholar]
- Orange, J.P.; Dinh, E.T.N.; Goodfriend, O.; Citino, S.B.; Wisely, S.M.; Blackburn, J.K. Evidence of Epizootic Hemorrhagic Disease Virus and Bluetongue Virus Exposure in Nonnative Ruminant Species in Northern Florida. J. Zoo Wildl. Med. 2020, 51, 745–751. [Google Scholar] [CrossRef]
- Favero, C.M.; Matos, A.C.D.; Campos, F.S.; Cândido, M.V.; Costa, É.A.; Heinemann, M.B.; Barbosa-Stancioli, E.F.; Lobato, Z.I.P. Epizootic Hemorrhagic Disease in Brocket Deer, Brazil. Emerg. Infect. Dis. 2013, 19, 346–348. [Google Scholar] [CrossRef]
- Schwartz-Cornil, I.; Mertens, P.; Contreras, V.; Hemati, B.; Pascale, F.; Bréard, E.; Mellor, P.; Maclachlan, N.; Zientara, S. Bluetongue virus: Virology, pathogenesis and immunity. Vet. Res. 2008, 39, 16. [Google Scholar] [CrossRef]
- Mullens, B.A.; Dada, C.E. Spatial and seasonal distribution of potential vectors of hemorrhagic disease viruses to peninsular bighorn sheep in the Santa Rosa mountains of southern California. J. Wildl. Dis. 1992, 28, 192–205. [Google Scholar] [CrossRef] [PubMed]
- Borden, E.C.; Shoorijeh, S.J.; Murphy, F.A. Physiochemical and morphological relationships of some athropod-borne viruses to bluetongue virus—A new taxonomic group. Physiochemical and serological studies. J. Gen. Virol. 1971, 13, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Hendrix, C.M. Biting Midges. In Merck Veterinary Manual; Merck Sharp & Dohme Corp.: Kenilworth, NJ, USA, 2013; Available online: https://www.merckvetmanual.com/integumentary-system/flies/biting-midges?query=biting%20midges (accessed on 20 July 2021).
- Virus Taxonomy: 2020 Release. International Committee on Taxonomy of Viruses (ICTV). March 2021. Available online: https://talk.ictvonline.org/taxonomy (accessed on 20 July 2021).
- Drolet, B.S.; Van Rijn, P.; Howerth, E.W.; Beer, M.; Mertens, P.P. A review of knowledge gaps and tools for orbivirus research. Vector Borne Zoonotic Dis. 2015, 15, 339–347. [Google Scholar] [CrossRef]
- Roy, P. Bluetongue viruses. In Desk Encyclopedia Animal and Bacterial Virology; Academic Press: Cambridge, MA, USA, 2008; pp. 43–50. [Google Scholar]
- De Clercq, K.; Vandaele, L.; Vanbinst, T.; Riou, M.; Deblauwe, I.; Wesselingh, W.; Pinard, A.; Van Eetvelde, M.; Boulesteix, O.; Leemans, B.; et al. Transmission of Bluetongue Virus Serotype 8 by Artificial Insemination with Frozen—Thawed Semen from Naturally Infected Bulls. Viruses 2021, 13, 652. [Google Scholar] [CrossRef] [PubMed]
- Osborn, D.A. Physical Condition Evaluation of Axis, Fallow, Sika, and White-Tailed Deer in Central Texas. Ph.D. Thesis, Texas Tech University, Lubbock, TX, USA, 1990. [Google Scholar]
- Work, T.M.; Jessup, D.A.; Sawyer, M.M. Experimental bluetongue and epizootic hemorrhagic disease virus infection in California black-tailed deer. J. Wildl. Dis. 1992, 28, 623–628. [Google Scholar] [CrossRef]
- Murray, J.O.; Trainer, D.O. Bluetongue virus in North American elk. J. Wildl. Dis. 1970, 6, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Corn, J.L.; Cartwright, M.E.; Alexy, K.J.; Cornish, T.E.; Manning, E.J.B.; Cartoceti, A.N.; Fischer, J.R. Surveys for disease agents in introduced elk in Arkansas and Kentucky. J. Wildl. Dis. 2010, 46, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Cover, M.A.; Hygnstrom, S.E.; Groepper, S.R.; Oates, D.W.; Hams, K.M.; VerCauteren, K.C. Surveillance of selected diseases in free-ranging elk (Cervus elaphus nelsoni) in Nebraska, 1995—2009. Gt. Plains Res. 2011, 21, 145–151. [Google Scholar]
- Thomas, F.C.; Prestwood, A.K. Plaque Neutralization Test Reactors to Bluetongue and EHD Viruses in the Southeastern USA. In Wildlife Diseases; Springer: Boston, MA, USA, 1976; pp. 401–411. [Google Scholar]
- Van Campen, H.; Davis, C.; Flinchum, J.D.; Bishop, J.V.; Schiebel, A.; Duncan, C.; Spraker, T. Epizootic hemorrhagic disease in yaks (Bos grunniens). J. Vet. Diagn. Investig. 2013, 25, 443–446. [Google Scholar] [CrossRef]
- Zarnke, R.L. Serologic survey for selected microbial pathogens in Alaskan wildlife. J. Wildl. Dis. 1983, 19, 324–329. [Google Scholar] [CrossRef]
- Stevens, G.; McCluskey, B.; King, A.; O’Hearn, E.; Mayr, G. Review of the 2012 epizootic hemorrhagic disease outbreak in domestic ruminants in the United States. PLoS ONE 2015, 10, e0133359. [Google Scholar] [CrossRef]
- Tessaro, S.V.; Clavijo, A. Duration of bluetongue viremia in experimentally infected American bison. J. Wildl. Dis. 2001, 37, 722–729. [Google Scholar] [CrossRef] [PubMed]
- Ostlund, E.N.; Moser, K.M.; Johnson, D.J.; Pearson, J.E.; Schmitt, B.J. Distribution of bluetongue in the United States of America, 1991–2002. Vet. Ital. 2004, 40, 83–88. [Google Scholar] [PubMed]
- Thorne, E.T.; Williams, E.S.; Spraker, T.R.; Helms, W.; Segerstrom, T. Bluetongue in free-ranging pronghorn antelope (Antilocapra americana) in Wyoming: 1976 and 1984. J. Wildl. Dis. 1988, 24, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Ortega, J.; Crossley, B.; Dechant, J.E.; Drew, C.P.; MacLachlan, N.J. Fatal bluetongue virus infection in an alpaca (Vicugna pacos) in California. J. Vet. Diagn. Investig. 2010, 22, 134–136. [Google Scholar] [CrossRef] [PubMed]
- Lear, A.S.; Callan, R.J. Overview of Bluetongue. In Merck Veterinary Manual; Merck Sharp & Dohme Corp.: Kenilworth, NJ, USA, 2014; Available online: https://www.merckvetmanual.com/generalized-conditions/bluetongue/overview-of-bluetongue (accessed on 20 July 2021).
- Coetzee, P.; Stokstad, M.; Myrmel, M.; Mutowembwa, P.; Loken, T.; Venter, E.H.; Van Vuuren, M. Transplacental infection in goats experimentally infected with a European strain of bluetongue virus serotype 8. Vet. J. 2013, 197, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Singer, R.S.; MacLachlan, N.J.; Carpenter, T.E. Maximal predicted duration of viremia in bluetongue virus—Infected cattle. J. Vet. Diagn. Investig. 2001, 13, 43–49. [Google Scholar] [CrossRef]
- Richards, R.; MacLachlan, N.; Heidner, H.; Fuller, F. Comparison of virologic and serologic responses of lambs and calves infected with bluetongue virus serotype 10. Vet. Microbiol. 1988, 18, 233–242. [Google Scholar] [CrossRef]
- Koumbati, M.; Mangana, O.; Nomikou, K.; Mellor, P.S.; Papadopoulos, O. Duration of bluetongue viraemia and serological responses in experimentally infected European breeds of sheep and goats. Vet. Microbiol. 1999, 64, 277–285. [Google Scholar] [CrossRef]
- Bonneau, K.R.; DeMaula, C.D.; Mullens, B.A.; MacLachlan, N.J. Duration of viraemia infectious to Culicoides sonorensis in bluetongue virus-infected cattle and sheep. Vet. Microbiol. 2002, 88, 115–125. [Google Scholar] [CrossRef]
- Maclachlan, N.J.; Gard, G.E.O.F.F. Clinical signs and pathology. In Bluetongue; Elsevier: Amsterdam, The Netherlands, 2009; pp. 285–293. [Google Scholar]
- Backx, A.; Heutink, R.; van Rooij, E.; van Rijn, P. Transplacental and oral transmission of wild-type bluetongue virus serotype 8 in cattle after experimental infection. Vet. Microbiol. 2009, 138, 235–243. [Google Scholar] [CrossRef]
- De Clercq, K.; Vandenbussche, F.; Vandemeulebroucke, E.; Vanbinst, T.; De Leeuw, I.; Verheyden, B.; Goris, N.; Mintiens, K.; Meroc, E.; Herr, C.; et al. Transplacental bluetongue infection in cattle. Vet. Rec. 2008, 162, 564. [Google Scholar] [CrossRef]
- Leudke, A.J.; Walton, T.E. Effect of natural breeding of heifers to a bluetongue virus carrier bull. Bov. Pract. 1981, 1981, 96–100. [Google Scholar]
- Zanella, G.; Durand, B.; Sellal, E.; Breard, E.; Sailleau, C.; Zientara, S.; Batten, C.A.; Mathevet, P.; Audeval, C. Bluetongue virus serotype 8: Abortion and transplacental transmission in cattle in the Burgundy region, France, 2008–2009. Theriogenology 2012, 77, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Mayo, C.E.; Crossley, B.M.; Hietala, S.K.; Gardner, I.A.; Breitmeyer, R.E.; MacLachlan, N.J. Colostral transmission of Bluetongue virus nucleic acid among newborn dairy calves in California. Transbound. Emerg. Dis. 2010, 57, 277–281. [Google Scholar] [CrossRef] [PubMed]
- Vögtlin, A.; Hofmann, M.A.; Nenniger, C.; Renzullo, S.; Steinrigl, A.; Loitsch, A.; Schwermer, H.; Kaufmann, C.; Thür, B. Long-term infection of goats with bluetongue virus serotype 25. Vet. Microbiol. 2013, 166, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Clarke, L.L.; Ruder, M.G.; Kienzle-Dean, C.; Carter, D.; Stallknecht, D.; Howerth, E.W. Experimental infection of white-tailed deer (odocoileus virginianus) with bluetongue virus serotype 3. J. Wildl. Dis. 2019, 55, 627–636. [Google Scholar] [CrossRef]
- Akita, G.Y.; Ianconescu, M.; MacLachlan, N.J.; Osburn, B.I.; Greene, R.T. Bluetongue disease in dogs associated with contaminated vaccine. Vet. Rec. 1994, 134, 283. [Google Scholar] [CrossRef]
- Brown, C.C.; Rhyan, J.C.; Grubman, M.J.; Wilbur, L.A. Distribution of bluetongue virus in tissues of experimentally infected pregnant dogs as determined by in situ hybridization. Vet. Pathol. 1996, 33, 337–340. [Google Scholar] [CrossRef] [PubMed]
- Jauniaux, T.P.; De Clercq, K.E.; Cassart, D.E.; Kennedy, S.; Vandenbussche, F.E.; Vandemeulebroucke, E.L.; Vanbinst, T.M.; Verheyden, B.I.; Goris, N.E.; Coignoul, F.L. Bluetongue in Eurasian lynx. Emerg. Infect. Dis. 2008, 14, 1496–1498. [Google Scholar] [CrossRef]
- Dunbar, M.R.; Cunningham, M.W.; Roof, J.C. Seroprevalence of selected disease agents from free-ranging black bears in Florida. J. Wildl. Dis. 1998, 34, 612–619. [Google Scholar] [CrossRef][Green Version]
- Scott, M. The Impact of Infection and Disease on Animal Populations: Implications for Conservation Biology. Conserv. Biol. 1988, 2, 40–56. [Google Scholar] [CrossRef]
- Ruder, M.G.; Stallknecht, D.E.; Allison, A.B.; Mead, D.G.; Carter, D.L.; Howerth, E.W. Host and potential vector susceptibility to an emerging orbivirus in the United States: Epizootic hemorrhagic disease virus serotype 6. Vet. Pathol. 2016, 53, 574–584. [Google Scholar] [CrossRef] [PubMed]
- Gaydos, J.K.; Davidson, W.R.; Elvinger, F.; Mead, D.G.; Howerth, E.W.; Stallknecht, D.E. Innate resistance to epizootic hemorrhagic disease in white-tailed deer. J. Wildl. Dis. 2002, 38, 713–719. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ditchfield, J.; Debbie, J.; Karstad, L. The virus of epizootic hemorrhagic disease of deer. In Transactions of the Twenty-Ninth North American Wildlife Natural Resources Conference; WMI Publications: Las Vegas, NV, USA, 1964; pp. 196–201. [Google Scholar]
- Gaydos, J.K.; Allison, A.B.; Hanson, B.A.; Yellin, A.S. Oral and fecal shedding of epizootic hemorrhagic disease virus, serotype 1 from experimentally infected white-tailed deer. J. Wildl. Dis. 2002, 38, 166–168. [Google Scholar] [CrossRef]
- Rivera, N.A.; Brandt, A.L.; Novakofski, J.E.; Mateus-Pinilla, N.E. Chronic wasting disease in cervids: Prevalence, impact and management strategies. Vet. Med. Res. Rep. 2019, 10, 123. [Google Scholar] [CrossRef]
- Vigil, S.L.; Ruder, M.G.; Shaw, D.; Wlodkowski, J.; Garrett, K.; Walter, M.; Corn, J.L. Apparent range expansion of Culicoides (Hoffmania) insignis (Diptera: Ceratopogonidae) in the southeastern United States. J. Med. Entomol. 2018, 55, 1043–1046. [Google Scholar] [CrossRef]
- Hill, C.A.; MacDonald, J.F. Biting Midges: Biology and Public Health Risk; Purdue Extension: West Lafayette, IN, USA, 2010; Available online: https://mdc.itap.purdue.edu/item.asp?itemID=18110 (accessed on 20 July 2021).
- USDA Bluetongue Standard Operating Procedures: Overview of Etiology and Ecology. 2016–2019. Available online: https://www.aphis.usda.gov/animal_health/emergency_management/downloads/sop/sop_btv_e-e.pdf (accessed on 20 July 2021).
- Mellor, P.S.; Carpenter, S.; Harrup, L.; Baylis, M.; Mertens, P.P.C. Bluetongue in Europe and the Mediterranean basin: History of occurrence prior to 2006. Prevent. Vet. Med. 2008, 87, 4–20. [Google Scholar] [CrossRef]
- McGregor, B.L.; Stenn, T.; Sayler, K.A.; Blosser, E.M.; Blackburn, J.K.; Wisely, S.M.; Burkett-Cadena, N.D. Host use patterns of Culicoides spp. biting midges at a big game preserve in Florida, U.S.A., and implications for the transmission of orbiviruses. Med. Vet. Entomol. 2019, 33, 110–120. [Google Scholar] [CrossRef]
- Maclachlan, N.J.; Guthrie, A.J. Re-emergence of bluetongue, African horse sickness, and other orbivirus diseases. Vet. Res. 2010, 41, 35. [Google Scholar] [CrossRef] [PubMed]
- Burns, D.A. Diseases caused by arthropods and other noxious animals. In Rook’s Textbook of Dermatology; Blackwell Publishing: Hoboken, NY, USA, 2004; pp. 1555–1618. [Google Scholar]
- Werner, D.; Groschupp, S.; Bauer, C.; Kampen, H. Breeding habitat preferences of major Culicoides species (Diptera: Ceratopogonidae) in Germany. Int. J. Environ. Res. Public Health 2020, 17, 5000. [Google Scholar] [CrossRef] [PubMed]
- Pullinger, G.D.; Busquets, M.G.; Nomikou, K.; Boyce, M.; Attoui, H.; Mertens, P.P. Identification of the Genome Segments of Bluetongue Virus Serotype 26 (Isolate KUW2010/02) that Restrict Replication in a Culicoides sonorensis Cell Line (KC Cells). PLoS ONE 2016, 11, e0149709. [Google Scholar] [CrossRef]
- Mertens, P.P.; Brown, F.; Sangar, D.V. Assignment of the genome segments of bluetongue virus type 1 to the proteins which they encode. Virology 1984, 135, 207–217. [Google Scholar] [CrossRef]
- Roy, P. Bluetongue virus genetics and genome structure. Virus Res. 1989, 13, 179–206. [Google Scholar] [CrossRef]
- Ratinier, M.; Caporale, M.; Golder, M.; Franzoni, G.; Allan, K.; Nunes, S.F.; Armezzani, A.; Bayoumy, A.; Rixon, F.; Shaw, A.; et al. Identification and characterization of a novel non-structural protein of bluetongue virus. PLoS Pathog. 2011, 7, e1002477. [Google Scholar] [CrossRef]
- Chaignat, V.; Worwa, G.; Scherrer, N.; Hilbe, M.; Ehrensperger, F.; Batten, C.; Cortyen, M.; Hofmann, M.; Thuer, B. Toggenburg Orbivirus, a new bluetongue virus: Initial detection, first observations in field and experimental infection of goats and sheep. Vet. Microbiol. 2009, 138, 11–19. [Google Scholar] [CrossRef]
- Howell, P.G.; Kumm, N.A.; Botha, M.J. The application of improved techniques to the identification of strains of bluetongue virus. Onderstepoort. J. Vet. Res. 1970, 37, 59–66. [Google Scholar]
- Maan, S.; Maan, N.S.; Nomikou, K.; Batten, C.; Antony, F.; Belaganahalli, M.N.; Samy, A.M.; Reda, A.A.; Al-Rashid, S.A.; El, B.M.; et al. Novel bluetongue virus serotype from Kuwait. Emerg. Infect. Dis. 2011, 17, 886–889. [Google Scholar] [CrossRef] [PubMed]
- Savini, G.; Puggioni, G.; Meloni, G.; Marcacci, M.; Di Domenico, M.; Rocchigiani, A.M.; Spedicato, M.; Oggiano, A.; Manunta, D.; Teodori, L.; et al. Novel putative Bluetongue virus in healthy goats from Sardinia, Italy. Infect. Genet. Evol. 2017, 51, 108–117. [Google Scholar] [CrossRef]
- Sun, E.C.; Huang, L.P.; Xu, Q.Y.; Wang, H.X.; Xue, X.M.; Lu, P.; Li, W.J.; Liu, W.; Bu, Z.G.; Wu, D.L. Emergence of a novel bluetongue virus serotype, China 2014. Transbound. Emerg. Dis. 2016, 63, 585–589. [Google Scholar] [CrossRef]
- Vijaykrishna, D.; Mukerji, R.; Smith, G.J. RNA virus reassortment: An evolutionary mechanism for host jumps and immune evasion. PLoS Pathog. 2015, 11, e1004902. [Google Scholar] [CrossRef]
- Planzer, J.; Kaufmann, C.; Worwa, G.; Gavier-Widen, D.; Hofmann, M.A.; Chaignat, V.; Thuer, B. In vivo and in vitro propagation and transmission of Toggenburg orbivirus. Res. Vet. Sci. 2011, 91, e163–e168. [Google Scholar] [CrossRef]
- Christensen, S.A.; Ruder, M.G.; Williams, D.M.; Porter, W.F.; Stallknecht, D.E. The role of drought as a determinant of hemorrhagic disease in the eastern United States. Glob. Chang. Biol. 2020, 26, 3799–3808. [Google Scholar] [CrossRef] [PubMed]
- Mellor, P.S.; Boorman, J.; Baylis, M. Culicoides biting midges: Their role as arbovirus vectors. Annu. Rev. Entomol. 2000, 45, 307–340. [Google Scholar] [CrossRef]
- Lysyk, T.J.; Dergousoff, S.J. Distribution of Culicoides sonorensis (Diptera: Ceratopogonidae) in Alberta, Canada. J. Med. Entomol. 2014, 51, 560–571. [Google Scholar] [CrossRef] [PubMed]
- Allison, A.B.; Goekjian, V.H.; Potgieter, A.C.; Wilson, W.C.; Johnson, D.J.; Mertens, P.P.C.; Stallknecht, D.E. Detection of a novel reassortant epizootic hemorrhagic disease virus (EHDV) in the USA containing RNA segments derived from both exotic (EHDV-6) and endemic (EHDV-2) serotypes. J. Gen. Virol. 2010, 91, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.E.; Patel, N.G.; Levy, M.A.; Storeygard, A.; Balk, D.; Gittleman, J.L.; Daszak, P. Global trends in emerging infectious diseases. Nature 2008, 451, 990–993. [Google Scholar] [CrossRef] [PubMed]
- Green, A.L.; Dargatz, D.A.; Schmidtmann, E.T.; Herrero, M.V.; Seitzinger, A.H.; Ostlund, E.N.; Wagner, B.A.; Moser, K.M.; Wineland, N.E.; Walton, T.E. Risk factors associated with herd-level exposure of cattle in Nebraska, North Dakota, and South Dakota to bluetongue virus. Am. J. Vet. Res. 2005, 66, 853–860. [Google Scholar] [CrossRef]
- Mecham, J.O.; Nunamaker, R.A. Complex interactions between vectors and pathogens: Culicoides variipennis sonorensis (Diptera: Ceratopogonidae) infection rates with bluetongue viruses. J. Med. Entomol. 1994, 31, 903–907. [Google Scholar] [CrossRef]
- Tabachnick, W.J. Genetic control of oral susceptibility to infection of Culicoides variipennis with bluetongue virus. Am. J. trop. Med. Hyg. 1991, 45, 666–671. [Google Scholar] [CrossRef]
- Wittmann, E.J.; Mellor, P.S.; Baylis, M. Effect of temperature on the transmission of orbiviruses by the biting midge, Culicoides sonorensis. Med. Vet. Entomol. 2002, 16, 147–156. [Google Scholar] [CrossRef]
- Brand, S.P.; Keeling, M.J. The impact of temperature changes on vector-borne disease transmission: Culicoides midges and bluetongue virus. J. R. Soc. Interface 2017, 14, 20160481. [Google Scholar] [CrossRef]
- NOAA National Climatic Data Center. State of the Climate: National Overview for Annual 2012. Published Online December 2012. Available online: http://www.ncdc.noaa.gov/sotc/national/2012/13 (accessed on 20 July 2021).
- Mullens, B.A. A Quantitative Survey of Culicoides variipennis (Diptera: Ceratopogonidae) in dairy waste water ponds in Southern California. J. Med. Entomol. 1989, 26, 559–565. [Google Scholar] [CrossRef]
- Casey, C.L.; Rathbun, S.L.; Stallknecht, D.E.; Ruder, M.G. Spatial Analysis of the 2017 Outbreak of Hemorrhagic Disease and Physiographic Region in the Eastern United States. Viruses 2021, 13, 550. [Google Scholar] [CrossRef]
- Carpenter, S.; Wilson, A.; Mellor, P.S. Culicoides and the emergence of bluetongue virus in northern Europe. Trends Microbiol. 2009, 17, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Purse, B.V.; Mellor, P.S.; Rogers, D.J.; Samuel, A.R.; Mertens, P.P.; Baylis, M. Climate change and the recent emergence of blue- tongue in Europe. Nat. Rev. Microbiol. 2005, 3, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Kedmi, M.; Herziger, Y.; Galon, N.; Cohen, R.M.; Perel, M.; Batten, C.; Braverman, Y.; Gottlieb, Y.; Shpigel, N.; Klement, E. The association of winds with the spread of EHDV in dairy cattle in Israel during an outbreak in 2006. Prev. Vet. Med. 2010, 96, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Guis, H.; Caminade, C.; Calvete, C.; Morse, A.P.; Tran, A.; Baylis, M. Modelling the effects of past and future climate on the risk of bluetongue emergence in Europe. J. R. Soc. Interface 2012, 9, 339–350. [Google Scholar] [CrossRef]
- Mayo, C.E.; Mullens, B.A.; Reisen, W.K.; Osborne, C.J.; Gibbs, E.P.J.; Gardner, I.A.; MacLachlan, N.J. Seasonal and interseasonal dynamics of bluetongue virus infection of dairy cattle and Culicoides sonorensis midges in northern California–implications for virus overwintering in temperate zones. PLoS ONE 2014, 9, e106975. [Google Scholar] [CrossRef] [PubMed]
- Köhler, W. Arboviruses and Human Disease. World Health Organization Technical Report Series No. 369. 84 S., 4 Tab. Genf 1967: World Health Organization. Sw. Fr. 4,-. J. Basic Microbiol. 1968, 8, 249. [Google Scholar]
- Schmidtmann, E.T.; Jones, C.J.; Gollands, B. Comparative host-seeking activity of Culicdides (Diptera: Ceratopogonidae) attracted to pastured livestock in central New York state, USA. J. Med. Entomol. 1980, 17, 221–231. [Google Scholar] [CrossRef]
- Batten, C.A.; Henstock, M.R.; Steedman, H.M.; Waddington, S.; Edwards, L.; Oura, C.A. Bluetongue virus serotype 26: Infection kinetics, pathogenesis and possible contact transmission in goats. Vet. Microbiol. 2013, 162, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Cauvin, A.; Dinh, E.T.; Orange, J.P.; Shuman, R.M.; Blackburn, J.K.; Wisely, S.M. Antibodies to epizootic hemorrhagic disease virus (EHDV) in farmed and wild Florida white-tailed deer (Odocoileus virginianus). J. Wildl. Dis. 2020, 56, 208–213. [Google Scholar] [CrossRef]
- McGregor, B.L.; Blackburn, J.K.; Wisely, S.M.; Burkett-Cadena, N.D. Culicoides (Diptera: Ceratopogonidae) Communities Differ Between a Game Preserve and Nearby Natural Areas in Northern Florida. J. Med. Entomol. 2021, 58, 450–457. [Google Scholar] [CrossRef]
- Purse, B.V.; Falconer, D.; Sullivan, M.J.; Carpenter, S.; Mellor, P.S.; Piertney, S.B.; Mordue, A.J.; Albon, S.; Gunn, G.J.; Blackwell, A. Impacts of climate, host and landscape factors on Culicoides species in Scotland. Med. Vet. Entomol. 2012, 26, 168–177. [Google Scholar] [CrossRef]
- Searle, K.R.; Blackwell, A.; Falconer, D.; Sullivan, M.; Butler, A.; Purse, B.V. Identifying environmental drivers of insect phenology across space and time: Culicoides in Scotland as a case study. Bull. Entomol. Res. 2013, 103, 155. [Google Scholar] [CrossRef]
- Becker, M.E.; Roberts, J.; Schroeder, M.E.; Gentry, G.; Foil, L.D. Prospective study of epizootic hemorrhagic disease virus and bluetongue virus transmission in captive ruminants. J. Med. Entomol. 2020, 57, 1277–1285. [Google Scholar] [CrossRef]
- Melville, L.; Weir, R.; Hunt, N.; Davis, S.; Walsh, S. Forty years continuous monitoring for bluetongue virus at an Australian site of high arbovirus activity. What have we achieved. Vet. Ital. 2015, 51, 263–267. [Google Scholar]
- Backx, A.; Heutink, C.G.; Van Rooij, E.M.; Van Rijn, P.A. Clinical signs of bluetongue virus serotype 8 infection in sheep and goats. Vet. Rec. 2007, 161, 591–592. [Google Scholar] [CrossRef] [PubMed]
- Darpel, K.E.; Batten, C.A.; Veronesi, E.; Shaw, A.E.; Anthony, S.; Bachanek- Bankowska, K.; Kgosana, L.; Bin-Tarif, A.; Carpenter, S.; Muller-Doblies, U.U.; et al. Clinical signs and pathology shown by British sheep and cattle infected with bluetongue virus serotype 8 derived from the 2006 outbreak in northern Europe. Vet. Rec. 2007, 161, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Leemans, J.; Raes, M.; Vanbinst, T.; De Clercq, K.; Saegerman, C.; Kirschvink, N. Viral RNA load in semen from bluetongue serotype 8-infected rams: Relationship with sperm quality. Vet. J. 2012, 192, 304–310. [Google Scholar] [CrossRef] [PubMed]
- Vanbinst, T.; Vandenbussche, F.; Dernelle, E.; De Clercq, K. A duplex real-time RT-PCR for the detection of bluetongue virus in bovine semen. J. Virol. Methods 2010, 169, 162–168. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Ruminant Species | Detected (✓ ) | References | |||
|---|---|---|---|---|---|
| Family | Common Name | Latin Name | BTV | EHDV | |
| Cervidae | White-tailed deer | Odocoileus virginianus | ✓ | ✓ | [9,10,20,41,51] |
| Mule deer | Odocoileus hemionus | ✓ | ✓ | [10] | |
| Black-tailed deer | Odocoileus hemionus columbianus | ✓ | ✓ | [20,52] | |
| Elk (wapiti) | Cervus canadensis | ✓ | ✓ | [10,41] | |
| Rocky Mountain Elk | Cervus elaphus nelsoni | ✓ | ✓ | [53,54,55] | |
| Axis deer | Axis axis | ✓ | ✓ | [10,51] | |
| Fallow deer | Dama dama | ✓ | ✓ | [10,41,51,56] | |
| Sika deer | Cervus nippon | ✓ | ✓ | [10] | |
| Yaks | Bos grunniens | ✓ | [57] | ||
| Père David’s deer | Elaphurus davidianus | ✓ | [41] | ||
| Moose | Alces alces | ✓ | [58] | ||
| Bovidae | Cattle | Bos taurus | ✓ | ✓ | [9,56,59] |
| Mountain goat | Oreamnos americanus | ✓ | [9,10] | ||
| Bison | Bison bison | ✓ | ✓ | [10,59,60] | |
| Blackbuck antelope | Antilope cervicapra | ✓ | ✓ | [10] | |
| Gerenuk | Litocranius walleri | ✓ | [61] | ||
| Bighorn sheep | Ovis canadensis | ✓ | ✓ | [9,10,59] | |
| Dall sheep | Ovis dalli | ✓ | [58] | ||
| Bongo antelope | Tragelaphus eurycerus | ✓ | [41] | ||
| Roan antelope | Hippotragus equinus | ✓ | [41] | ||
| Lesser kudu | Tragelaphus imberbis | ✓ | [41] | ||
| Dama gazelle | Nanger dama | ✓ | [41] | ||
| Antilocapridae | Pronghorn | Antilocapra americana | ✓ | ✓ | [10,62] |
| Camelidae | Alpaca | Vicugna pacos | ✓ | [63] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rivera, N.A.; Varga, C.; Ruder, M.G.; Dorak, S.J.; Roca, A.L.; Novakofski, J.E.; Mateus-Pinilla, N.E. Bluetongue and Epizootic Hemorrhagic Disease in the United States of America at the Wildlife–Livestock Interface. Pathogens 2021, 10, 915. https://doi.org/10.3390/pathogens10080915
Rivera NA, Varga C, Ruder MG, Dorak SJ, Roca AL, Novakofski JE, Mateus-Pinilla NE. Bluetongue and Epizootic Hemorrhagic Disease in the United States of America at the Wildlife–Livestock Interface. Pathogens. 2021; 10(8):915. https://doi.org/10.3390/pathogens10080915
Chicago/Turabian StyleRivera, Nelda A., Csaba Varga, Mark G. Ruder, Sheena J. Dorak, Alfred L. Roca, Jan E. Novakofski, and Nohra E. Mateus-Pinilla. 2021. "Bluetongue and Epizootic Hemorrhagic Disease in the United States of America at the Wildlife–Livestock Interface" Pathogens 10, no. 8: 915. https://doi.org/10.3390/pathogens10080915
APA StyleRivera, N. A., Varga, C., Ruder, M. G., Dorak, S. J., Roca, A. L., Novakofski, J. E., & Mateus-Pinilla, N. E. (2021). Bluetongue and Epizootic Hemorrhagic Disease in the United States of America at the Wildlife–Livestock Interface. Pathogens, 10(8), 915. https://doi.org/10.3390/pathogens10080915