
Immunogenic Peptides from Pap31 and SCS-α of Bartonella bacilliformis: One Step Closer to a Rapid Diagnostic Tool for Carrion’s Disease

 , ,
, ,
Abstract
1. Introduction
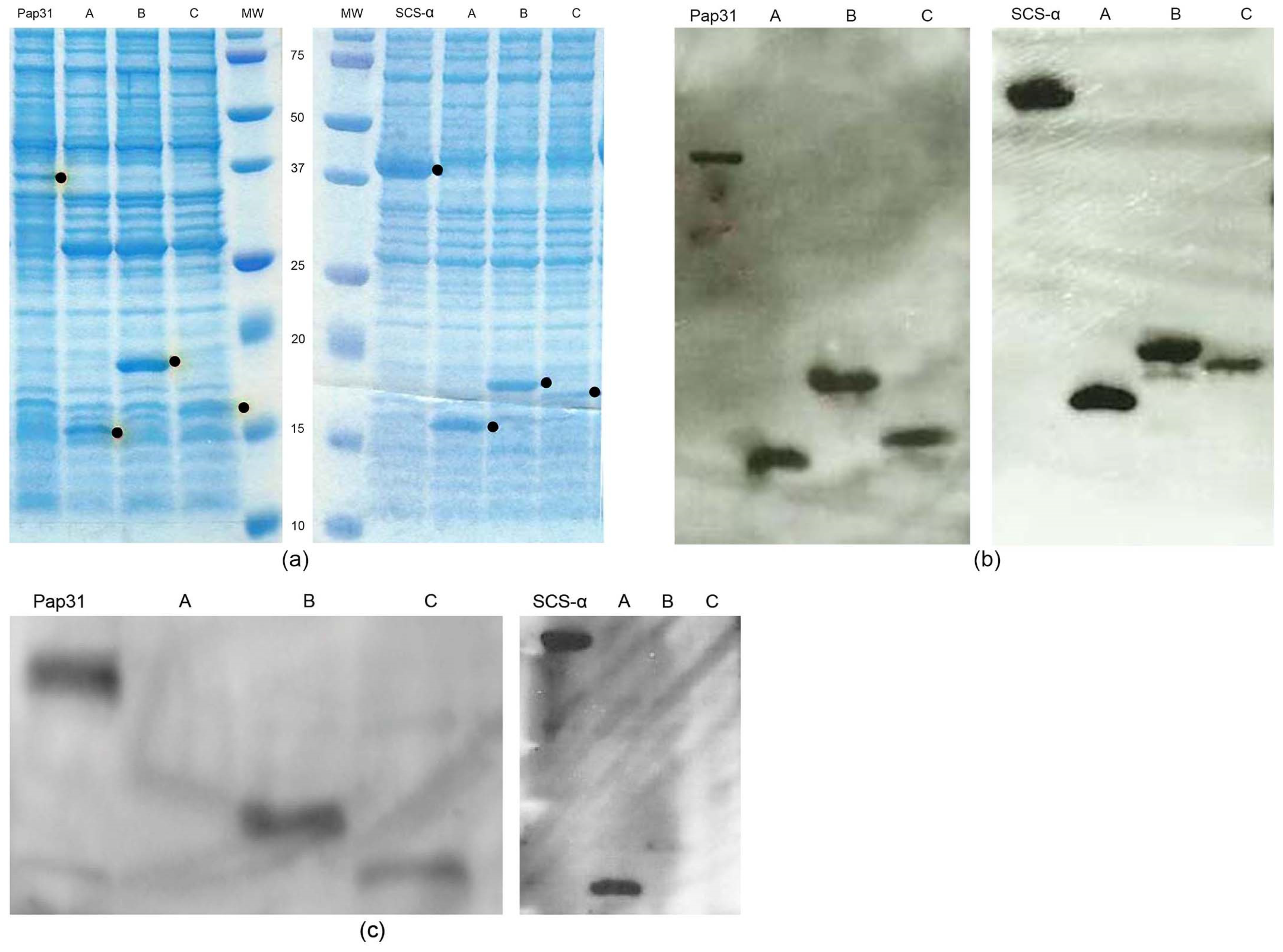
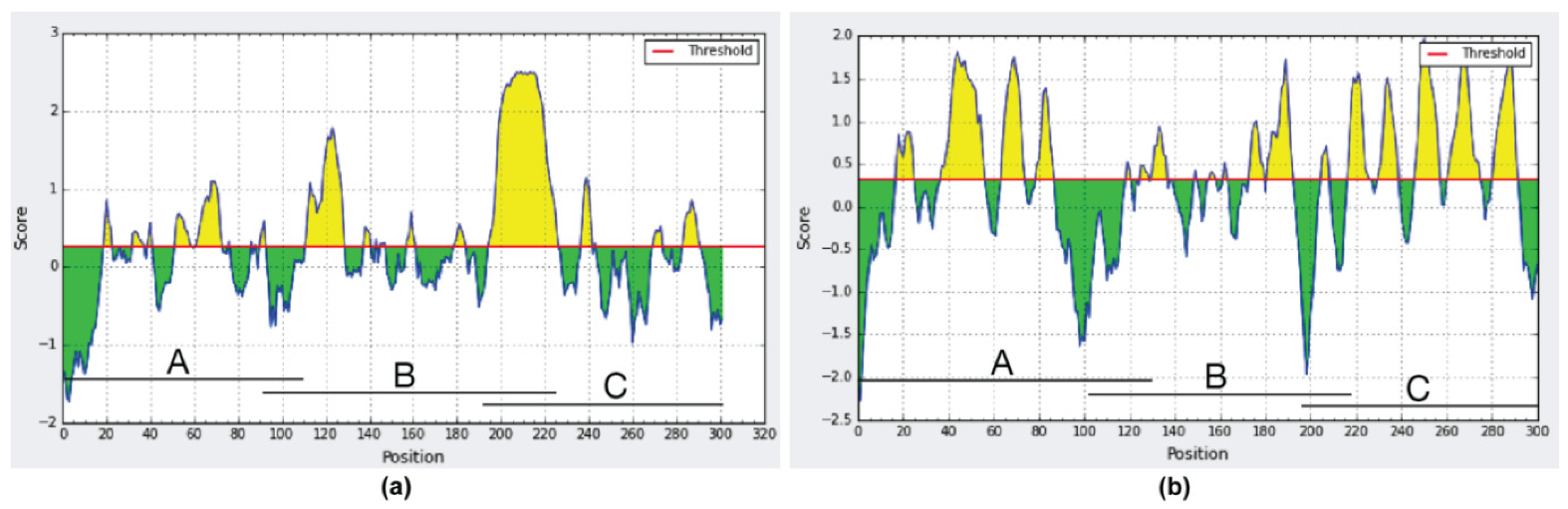
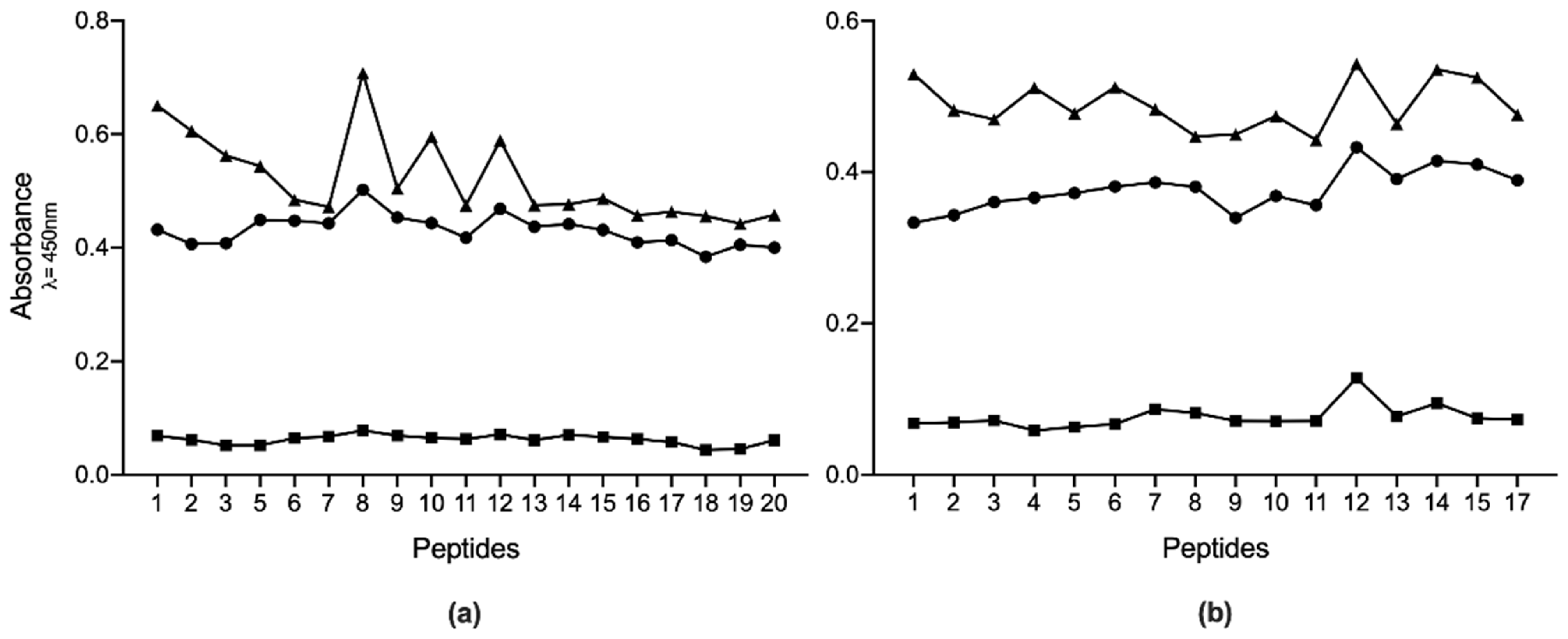
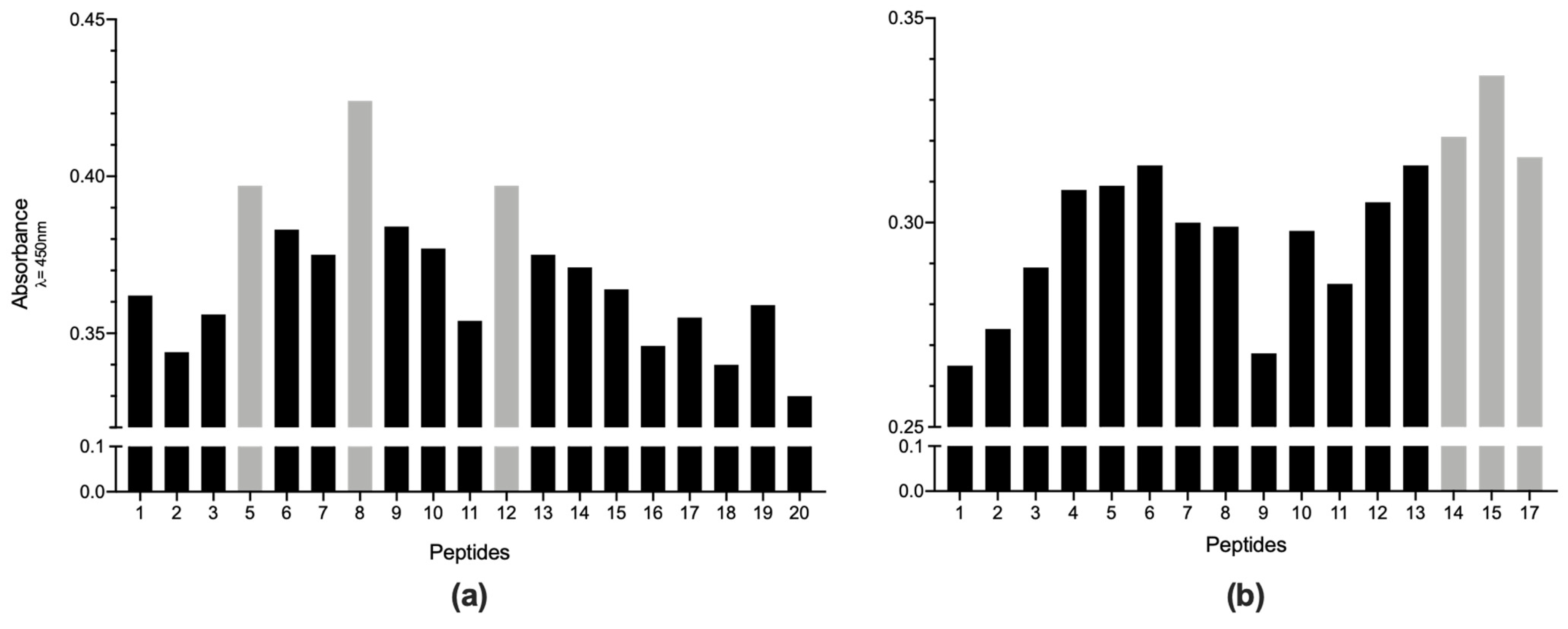
2. Results
3. Discussion
4. Materials and Methods
4.1. Serum Samples
4.2. Bacterial Strains
4.3. Amplification, Cloning, and Purification of Antigenic Proteins and Respective Truncated Constructs
4.4. Western Blotting
4.5. In-Silico Analysis
4.6. Design and Synthesis of Biotinylated Peptides
4.7. ELISA
4.8. Selection of the Best Immunogenic Candidates
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gomes, C.; Ruiz, J. Carrion’s disease. The sound of silence. Clin. Microb. Rev. 2018, 31, e00056-17. [Google Scholar] [CrossRef]
- Pons, M.J.; Gomes, C.; del Valle, J.; Ruiz, J. Carrion’s Disease, more than a sandfly-vectored illness. PLoS Path. 2016, 12, e1005863. [Google Scholar] [CrossRef]
- Gray, G.C.; Johnson, A.A.; Thornton, S.A.; Smith, W.A.; Knobloch, J.; Kelley, P.W.; Obregon Escudero, L.; Arones Huayda, M.; Wignall, F.S. An epidemic of Oroya fever in the Peruvian Andes. Am. J. Trop. Med. Hyg. 1990, 42, 215–221. [Google Scholar] [CrossRef]
- Chamberlin, J.; Laughlin, L.W.; Romero, S.; Solórzano, N.; Gordon, S.; Andre, R.G.; Pachas, P.; Friedman, H.; Ponce, C.; Watts, D. Epidemiology of endemic Bartonella bacilliformis: A prospective cohort study in a Peruvian mountain valley community. J. Infect. Dis. 2002, 186, 983–990. [Google Scholar] [CrossRef] [PubMed]
- Gomes, C.; Palma, N.; Pons, M.J.; Magallón-Tejada, A.; Sandoval, I.; Tinco-Valdez, C.; Gutarra, C.; del Valle-Mendoza, J.; Ruiz, J.; Matsuoka, M. Succinyl-CoA synthetase: New antigen candidate of Bartonella bacilliformis. PLoS Negl. Trop. Dis. 2016, 10, e0004989. [Google Scholar] [CrossRef] [PubMed]
- Chamberlin, J.; Laughlin, L.; Gordon, S.; Romero, S.; Solórzano, N.; Regnery, R.L. Serodiagnosis of Bartonella bacilliformis infection by indirect fluorescence antibody assay: Test development and application to a population in an area of bartonellosis endemicity. J. Clin. Microbiol. 2000, 38, 4269–4271. [Google Scholar] [CrossRef]
- Maguiña, C.; García, P.J.; Gotuzzo, E.; Cordero, L.; Spach, D.H. Bartonellosis (Carrión’s disease) in the modern era. Clin. Infect. Dis. 2001, 33, 772–779. [Google Scholar] [CrossRef] [PubMed]
- Cornejo, A.; Gomes, C.; Suarez, L.; Martinez-Puchol, S.; Bustamante, P.; Pons, M.J.; Ruiz, J.; del Valle, J. An unidentified cluster of infection in the Peruvian Amazon region. J. Infect. Dev. Ctries 2015, 9, 524–529. [Google Scholar] [CrossRef]
- del Valle, J.; Silva, W.; Tinco, C.; Pons, M.J.; del Valle, L.J.; Champin Michelena, D.; Bazán Mayra, J.; Zavaleta Gavidea, V.; Vargas, M.; Ruiz, J. Diagnosis of Carrion’s disease by direct blood PCR in thin blood smear negative samples. PLoS ONE 2014, 9, e92283. [Google Scholar] [CrossRef]
- Ellis, B.A.; Rotz, L.D.; Leake, J.A.; Samalvides, F.; Bernable, J.; Ventura, G.; Padilla, C.; Villaseca, P.; Beati, L.; Regnery, R.; et al. An outbreak of acute bartonellosis (Oroya fever) in the Urubamba region of Peru, 1998. Am. J. Trop. Med. Hyg. 1999, 61, 344–349. [Google Scholar] [CrossRef]
- Sanchez Clemente, N.; Ugarte-Gil, C.A.; Solórzano, N.; Maguiña, C.; Pachas, P.; Blazes, D.; Bailey, R.; Mabey, D.; Moore, D. Bartonella bacilliformis: A systematic review of the literature to guide the research agenda for elimination. PLoS Negl. Trop. Dis. 2012, 6, e1819. [Google Scholar] [CrossRef]
- Taye, A.; Chen, H.; Duncan, K.; Zhang, Z.; Hendrix, L.; Gonzalez, J.; Ching, W. Production of recombinant protein Pap31 and its application for the diagnosis of Bartonella bacilliformis infection. Ann. N. Y. Acad. Sci. 2005, 1063, 280–285. [Google Scholar] [CrossRef] [PubMed]
- Angkasekwinai, N.; Atkins, E.H.; Romero, S.; Grieco, J.; Chao, C.C.; Ching, W.M. An evaluation study of enzyme-linked immunosorbent assay (ELISA) using recombinant protein Pap31 for detection of antibody against Bartonella bacilliformis infection among the Peruvian population. Am. J. Trop. Med. Hyg. 2014, 90, 690–696. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Henriquez-Camacho, C.A.; Ventosilla, P.; Minnick, M.; Ruiz, J.; Maguiña, C. Proteins of Bartonella bacilliformis: Candidates for vaccine development. Int. J. Pept. 2015, 2015, 702784. [Google Scholar] [CrossRef] [PubMed]
- Teixeira-Gomes, A.P.; Cloeckaert, A.; Bézard, G.; Bowden, R.A.; Dubray, G.; Zygmunt, M.S. Identification and characterization of Brucella ovis immunogenic proteins using two-dimensional electrophoresis and immunoblotting. Electrophoresis 1997, 18, 1491–1497. [Google Scholar] [CrossRef]
- Teixeira-Gomes, A.P.; Cloeckaert, A.; Bézard, G.; Dubray, G.; Zygmunt, M.S. Mapping and identification of Brucella melitensis proteins by two-dimensional electrophoresis and microsequencing. Electrophoresis 1997, 18, 156–162. [Google Scholar] [CrossRef]
- Ruiz, J.; Pons, M.J. Revisiting Bartonella bacilliformis MLST. Infect. Genet. Evol. 2018, 63, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Loparev, V.N.; Dasch, G.A.; Knipe, K.M.; Rowe, L.A.; Lydy, S.L. Optical mapping and genome sequences of isolates of Bartonella bacilliformis. In Proceedings of the Program and Abstracts Book of International Conference on Emerging Infectious Diseases, Atlanta, GA, USA, 24–26 August 2015; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2015; p. 6. [Google Scholar]
- Paul, S.; Minnick, M.F.; Chattopadhyay, S. Mutation-driven divergence and convergence indicate adaptive evolution of the intracellular human-restricted pathogen, Bartonella bacilliformis. PLoS Negl. Trop. Dis. 2016, 10, e0004712. [Google Scholar] [CrossRef]
- Chan, W.S.; Au, C.H.; Leung, H.C.; Ho, D.N.; Li, D.; Chan, T.L.; Lam, T.W.; Ma, E.S.; Tang, B.S. Potential utility of metagenomic sequencing for improving etiologic diagnosis of infective endocarditis. Futur. Cardiol. 2019, 15, 411–424. [Google Scholar] [CrossRef] [PubMed]
- Fournier, P.E.; Couderc, C.; Buffet, S.; Flaudrops, C.; Raoult, D. Rapid and cost-effective identification of Bartonella species using mass spectrometry. J. Med. Microbiol. 2009, 58 Pt 9, 1154–1159. [Google Scholar] [CrossRef]
- Cretich, M.; Gori, A.; D’Annessa, I.; Chiari, M.; Colombo, G. Peptides for infectious diseases: From probe design to diagnostic microarrays. Antibodies 2019, 8, 23. [Google Scholar] [CrossRef]
- Feliciano, N.D.; Ribeiro, V.S.; Gonzaga, H.T.; Santos, F.A.; Fujimura, P.T.; Goulart, L.R.; Costa-Cruz, J.M. Short epitope-based synthetic peptides for serodiagnosis of human strongyloidiasis. Immunol. Lett. 2016, 172, 89–93. [Google Scholar] [CrossRef]
- Boonjakuakul, J.K.; Gerns, H.L.; Chen, Y.T.; Hicks, L.D.; Minnick, M.F.; Dixon, S.E.; Hall, S.C.; Koehler, J.E. Proteomic and immunoblot analyses of Bartonella quintana total membrane proteins identify antigens recognized by sera from infected patients. Infect. Immun. 2007, 75, 2548–2561. [Google Scholar] [CrossRef]
- Matsuoka, M.; Sasaki, T.; Seki, N.; Kobayashi, M.; Sawabe, K.; Sasaki, Y.; Shibayama, K.; Sasaki, T.; Arakawa, Y. Hemin-binding proteins as potent markers for serological diagnosis of infections with Bartonella quintana. Clin. Vaccine Immunol. 2013, 20, 620–626. [Google Scholar] [CrossRef] [PubMed]
- Saisongkorh, W.; Kowalczewska, M.; Azza, S.; Decloquement, P.; Rolain, J.M.; Raoult, D. Identification of candidate proteins for the diagnosis of Bartonella henselae infections using an immunoproteomic approach. FEMS Microbiol. Lett. 2010, 310, 158–167. [Google Scholar] [CrossRef]
- Carroll, J.A.; Coleman, S.A.; Smitherman, L.S.; Minnick, M.F. Hemin-binding surface protein from Bartonella quintana. Infect. Immun. 2000, 68, 6750–6757. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, J.; Gomes, C. In silico analysis of Pap31 from Bartonella bacilliformis and other Bartonella spp. Infect. Genet. Evol. 2020, 84, 104482. [Google Scholar] [CrossRef] [PubMed]
- Sakolvaree, Y.; Maneewatch, S.; Jiemsup, S.; Klaysing, B.; Tongtawe, P.; Srimanote, P.; Saengjaruk, P.; Banyen, S.; Tapchaisri, P.; Chonsa-nguan, M.; et al. Proteome and immunome of pathogenic Leptospira spp. revealed by 2DE and 2DE-immunoblotting with immune serum. Asian Pac. J. Allergy Immunol. 2007, 25, 53–73. [Google Scholar]
- Chang, C.C.; Chen, Y.J.; Tseng, C.S.; Lai, W.L.; Hsu, K.Y.; Chang, C.L.; Lu, C.C.; Hsu, Y.M. A comparative study of the interaction of Bartonella henselae strains with human endothelial cells. Vet. Microbiol. 2011, 149, 147–156. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | % 1 | Organism 2 | Protein 3 | GenBank 4 | Sequence |
|---|---|---|---|---|---|
| QAIGSAILKGTKDTGT | 100.00 | Bartonella bacilliformis5 | Pap31 | WP_005767899 | QAIGSAILKGTKDTGT |
| 93.75 | Bartonella bacilliformis6 | Pap31 | WP_041848867 | QAIGSAILKGVKDTGT | |
| 68.75 | Escherichia coli | MdtK | EEW1185817 | QAIGSGILRGYKDTRS | |
| 68.75 | Klebsiella pneumoniae7 | MdtK | WP_032418890 | QAIGSGILRGYKDTRS | |
| IFASVAEGKEKTGANA 8 | 100.00 | Bartonella bacilliformis9 | SCS-α | WP_035454724 | IFASVAEGKEKTGANA |
| 93.75 | Bartonella bacilliformis10 | SCS-α | WP_005765714 | IFASVTEGKEKTGANA | |
| 93.75 | Bartonella bacilliformis6 | SCS-α | WP_041849026 | IFASVAEGKEKTEANA | |
| 93.75 | Bartonella ancashesis10 | SCS-α | WP_053943861 | IFASVMEGKEKTGANA | |
| 93.75 | Bartonella alsatica | SCS-α | WP_005866529 | IFASVAEGKEKTGADA | |
| 93.75 | Bartonella clarridgeiae | SCS-α | WP_013545653 | IFASVVEGKEKTGANA | |
| 93.75 | Bartonella henselae | SCS-α | WP_005866529 | IFASVAEGKEKTGADA | |
| 93.75 | Candidatus Bartonella melophagi | SCS-α | WP_007476332 | IFASVAEGKEKTGADA | |
| 93.75 | Bartonella schoenbuchensis | SCS-α | WP_010704443 | IFASVAEGKEKTGADA | |
| 93.75 | Bartonella vinsonii | SCS-α | WP_010705938 | IFASVAEGKEKTGADA | |
| 87.50 | Bartonella elizabethae | SCS-α | WP_005774305 | IFASVAEAKEKTGADA | |
| 87.50 | Bartonella koehlearae | SCS-α | WP_034458042 | IFASVAEGKEKTDADA | |
| 87.50 | Bartonella quintana | SCS-α | WP_011179955 | IFASVAEGKERTGADA | |
| 87.50 | Bartonella rochalimae | SCS-α | WP_035006433 | IFASVVEGKEKTGADA | |
| 87.50 | Candidatus Bartonella tamiae | SCS-α | WP_008037783 | IFSSVAEGKEKTGADA | |
| 87.50 | Bartonella tribocorum | SCS-α | WP_100129872 | IFASVAEAKEKTGADA | |
| 87.50 | Candidatus Bartonella washoensis | SCS-α | WP_006925899 | IFSSVAEGKEKTGADA | |
| 87.50 | Brevundimonas spp. | SCS-α | WP_087141820 | IFASVAEGKERTGADA | |
| 81.25 | Bartonella grahamii | SCS-α | WP_015857104 | IFANVAEAKEKTGADA | |
| 81.25 | Bartonella doshiae | SCS-α | WP_004856238 | IFASVAEAKEKTDADA | |
| 81.25 | Brucella spp. | SCS-α | WP_004684457 | IFATVAEGKERTGADA | |
| 68.75 | Nocardia spp. | SCS-α | WP_067485390 | VFASVAEAMEKTGADT | |
| 75.00 | Human | SCS-α | SJM31584 | IFSTVAEGKATTGANA |
| Description | Source or Reference | |
|---|---|---|
| Strains | ||
| B. bacilliformis | CIP 57.20—NCTC12136 | Institute Pasteur |
| E. coli TOP10 | Host strain for cloning | Invitrogen |
| E. coli BL21Star (DE3) | Host strain for gene expression | Invitrogen |
| Plasmids | ||
| pCR4-TOPO | TA-cloning vector | Invitrogen |
| pET100D/TOPO | Expression vector | Invitrogen |
| pPap31 | pET100D/TOPO containing Pap31 | [5] |
| pPap31–A | pET100D/TOPO containing clone A of Pap31 | This study |
| pPap31–B | pET100D/TOPO containing clone B of Pap31 | This study |
| pPap31–C | pET100D/TOPO containing clone C of Pap31 | This study |
| pSCS-α | pET100D/TOPO containing SCS-α | [5] |
| pSCS-α–A | pET100D/TOPO containing clone A of SCS-α | This study |
| pSCS- α–B | pET100D/TOPO containing clone B of SCS-α | This study |
| pSCS- α–C | pET100D/TOPO containing clone C of SCS-α | This study |
| Sequence (5′→3′) | Fragment (bp) | Ref. | |
|---|---|---|---|
| Pap31 F | 1 CACCATGAATATAAAATGTTTAGTGACA | 1–903 | [5] |
| Pap31 R | TCAGAATTTGTAAGCAACACCAACGCG | ||
| Pap31–A F | 1 CACCATGAATATAAAATGTTTAGTGACA | 1–325 | This study |
| Pap31–A R | 2 TCACAACCCAAACAATATCTG | ||
| Pap31–B F | 1 CACCATCTCGGCAGTGGCCTT | 275–675 | This study |
| Pap31–B R | 2 TCATACCTGCTTTGCTAGCACTAC | ||
| Pap31–C F | 1 CACCTATCTTAAAAGGCACG | 576–903 | This study |
| Pap31–C R | TCAGAATTTGTAAGCAACACCAACGCG | ||
| SCS α F | 1 CACCATGTCAATTCTTATC | 1–903 | [5] |
| SCS α R | CTAACCCTTCAAGACTGAAACC | ||
| SCS α–A F | 1 CACCATGTCAATTCTTATC | 1–381 | This study |
| SCS α–A R | 2 TCAACCAATTAAGCGCGATTTCG | ||
| SCS α–B F | 1 CACCTGTATTACAGAAGGTATACC | 301–651 | This study |
| SCS α–B R | 2 TCAACCAATCTCACCGATC | ||
| SCS α–C F | 1 CACCGATGTGTTAGAAATGT | 586–903 | This study |
| SCS α–C R | CTAACCCTTCAAGACTGAAACC |
| Peptide | Peptide Sequence | Position (aa 1) | |
|---|---|---|---|
| Pap 31 | 1 | IGFGADRIMPYVSGGV | 166–181 |
| 2 | GADRIMPYVSGGVAYT | 169–184 | |
| 3 | RIMPYVSGGVAYTQVQ | 172–187 | |
| 4 2 | PYVSGGVAYTQVQAIG | 175–190 | |
| 5 | SGGVAYTQVQAIGSAI | 178–193 | |
| 6 | VAYTQVQAIGSAILKG | 181–196 | |
| 7 | TQVQAIGSAILKGTKD | 184–199 | |
| 8 | QAIGSAILKGTKDTGT | 187–202 | |
| 9 | GSAILKGTKDTGTEGG | 190–205 | |
| 10 | ILKGTKDTGTEGGGGT | 193–208 | |
| 11 | GTKDTGTEGGGGTEGG | 196–211 | |
| 12 | DTGTEGGGGTEGGGGT | 199–214 | |
| 13 | TEGGGGTEGGGGTEGG | 202–217 | |
| 14 | GGGTEGGGGTEGGGGS | 205–220 | |
| 15 | TEGGGGTEGGGGSASK | 208–223 | |
| 16 | GGGTEGGGGSASKAVR | 211–226 | |
| 17 | TEGGGGSASKAVRSEA | 214–229 | |
| 18 | GGGSASKAVRSEALDV | 217–232 | |
| 19 | SASKAVRSEALDVLAS | 220–235 | |
| 20 | KAVRSEALDVLASGTI | 223–238 | |
| SCS-α | 1 | EQALAYHGTQMVGGIN | 26–41 |
| 2 | LAYHGTQMVGGINPKK | 29–44 | |
| 3 | HGTQMVGGINPKKGGE | 32–47 | |
| 4 | QMVGGINPKKGGETWT | 35–50 | |
| 5 | GGINPKKGGETWTGAK | 38–53 | |
| 6 | NPKKGGETWTGAKGET | 41–56 | |
| 7 | KGGETWTGAKGETLPI | 44–59 | |
| 8 | ETWTGAKGETLPIFAS | 47–62 | |
| 9 | TGAKGETLPIFASVAE | 50–65 | |
| 10 | KGETLPIFASVAEGKE | 53–68 | |
| 11 | TLPIFASVAEGKEKTG | 56–71 | |
| 12 | IFASVAEGKEKTGANA | 59–74 | |
| 13 | SVAEGKEKTGANASVI | 62–77 | |
| 14 | EGKEKTGANASVIYVP | 65–80 | |
| 15 | EKTGANASVIYVPPAG | 68–83 | |
| 16 2 | GANASVIYVPPAGAAD | 71–86 | |
| 17 | ASVIYVPPAGAADAII | 74–89 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomes, C.; Pons, M.J.; del Valle-Mendoza, J.; Matsuoka, M.; Ruiz, J. Immunogenic Peptides from Pap31 and SCS-α of Bartonella bacilliformis: One Step Closer to a Rapid Diagnostic Tool for Carrion’s Disease. Pathogens 2021, 10, 917. https://doi.org/10.3390/pathogens10080917
Gomes C, Pons MJ, del Valle-Mendoza J, Matsuoka M, Ruiz J. Immunogenic Peptides from Pap31 and SCS-α of Bartonella bacilliformis: One Step Closer to a Rapid Diagnostic Tool for Carrion’s Disease. Pathogens. 2021; 10(8):917. https://doi.org/10.3390/pathogens10080917
Chicago/Turabian StyleGomes, Cláudia, Maria J. Pons, Juana del Valle-Mendoza, Mayumi Matsuoka, and Joaquim Ruiz. 2021. "Immunogenic Peptides from Pap31 and SCS-α of Bartonella bacilliformis: One Step Closer to a Rapid Diagnostic Tool for Carrion’s Disease" Pathogens 10, no. 8: 917. https://doi.org/10.3390/pathogens10080917
APA StyleGomes, C., Pons, M. J., del Valle-Mendoza, J., Matsuoka, M., & Ruiz, J. (2021). Immunogenic Peptides from Pap31 and SCS-α of Bartonella bacilliformis: One Step Closer to a Rapid Diagnostic Tool for Carrion’s Disease. Pathogens, 10(8), 917. https://doi.org/10.3390/pathogens10080917