Abstract
Coxiella burnetii is the causative agent of Q fever. It can infect animals, humans, and birds, as well as ticks, and it has a worldwide geographical distribution. To better understand the epidemiology of C. burnetii in Somalia, ticks infesting camels were collected from five different regions, including Bari, Nugaal, Mudug, Sool, and Sanaag, between January and March 2018. Collected ticks were tested for C. burnetii and Coxiella-like endosymbiont DNA by using IS1111, icd, and Com1-target PCR assays. Moreover, sequencing of the 16S-rRNA was conducted. Molecular characterization and typing were done by adaA-gene analysis and plasmid-type identification. Further typing was carried out by 14-marker Multi-Locus Variable-Number Tandem Repeats (MLVA/VNTR) analysis. The investigated ticks (n = 237) were identified as Hyalomma spp. (n = 227, 95.8%), Amblyomma spp. (n = 8, 3.4%), and Ripicephalus spp. (n = 2, 0.8%), and 59.1% (140/237) of them were positive for Coxiella spp. While Sanger sequencing and plasmid-type identification revealed a C. burnetii that harbours the QpRS-plasmid, MLVA/VNTR genotyping showed a new genotype which was initially named D21. In conclusion, this is the first report of C. burnetii in ticks in Somalia. The findings denote the possibility that C. burnetii is endemic in Somalia. Further epidemiological studies investigating samples from humans, animals, and ticks within the context of “One Health” are warranted.
1. Introduction
Coxiella burnetii is an obligate intracellular zoonotic Gram-negative bacterium that causes Q fever [1,2,3,4,5]. It is considered one of the important emerging pathogens worldwide [6,7]. C. burnetii infection is sub-clinical in animals and does not result in any clinically detectable signs or symptoms except in pregnant animals where the infection has been associated with several reproductive disorders, including abortion, premature delivery, stillbirth, and weak offspring [6,8]. C. burnetii has been found in birth products (placenta and offspring), as well as in milk, vaginal mucus, faeces, and urine [6,9]. Moreover, it has also been found in blood and organs, such as the lungs, liver, and spleen [6]. The main reservoirs of C. burnetii are ruminants [6]. Its transmission to humans is through inhalation of contaminated aerosols, ingestion of raw milk, or contact with infected animals [6]. C. burnetii infection in humans is often asymptomatic/subclinical in up to 50% of cases or manifests as an acute febrile illness with pneumonia and/or hepatitis, depending on the geographical origin of the infection [10]. In a small proportion of patients (i.e., 1–2% of acute cases), the disease may progress to a chronic form that is clinically associated with patients having an endocarditis like illness [11]. Moreover, recent studies have also demonstrated the importance of vascular infection affecting the intima of large arteries [12,13,14]. The risk of C. burnetii infection is higher for people living in rural regions, animal owners/herders, veterinarians, and slaughterhouse/abattoir workers [6].
In ticks, C. burnetii has mostly been detected in Amblyomma spp., Rhipicephalus spp., Ixodes spp., and Dermacentor spp. [2,5]. Nevertheless, C. burnetii can naturally infect more than 40 different tick species [2,15]. These ticks can transmit the pathogen vertically (i.e., transstadial or transovarial) and horizontally, via bites or in faeces, to wild mammals and birds [2,15]. Infected ticks play a major role in the natural cycle of C. burnetiid infection and thus represent a principal vector and reservoir [2].
In Somalia, the ecology and epidemiology of C. burnetii are not well understood due to the scarcity and under representation of research studies exposing an already vulnerable population to a high burden of zoonotic diseases [16,17,18]. We therefore aimed to identify and screen hard tick samples collected from camel populations in five states of Somalia for Coxiella spp.
2. Results
A total of 237 ticks were collected from camels in five states in Somalia, including Mudug, Sanaag, Bari, Sool, and Nugaal. Nearly all identified ticks were Hyalomma spp. (n = 227, 95.8%). Only eight (3.4%) were identified as Amblyomma spp. and two (0.8%) as Ripicephalus spp.
All samples were screened for C. burnetii and Coxiella-like endosymbionts DNA using IS1111-PCR, and 59.1% (140/237) samples were positive, of which 138 (98.6%) were Hyalomma ticks and 2 (1.4%) were Amblyomma ticks. Using icd- and com1-PCR, still 19.0% (45/237) of the samples were positive for Coxiella spp. However, due to the extreme low amount of specific DNA in most of the samples (i.e., 71.4% showed PCR cycle threshold/Ct-values higher than 35), only 4 (2.9%) samples with a higher DNA amount were selected for further genomic classification.
Confirmation of C. burnetii with 16S rRNA-PCR was successful, and further classification based on the acute disease antigen A (adaA)-PCR revealed an adaA-negative genotype (all samples were adaA-negative). In addition, the screening for C. burnetii specific plasmid types showed that all samples harbour the QpRS-plasmid.
Enhanced genomic typing using 14-marker Multi-Locus Variant Analysis (MLVA) could amplify successfully 11 of the 14 MLVA markers (ms/microsatellites marker). Uploading the repeat pattern data to the CoxBase online platform (https://coxbase.q-gaps.de, accessed on 26 April 2021) revealed a probably new incomplete (i.e., not all markers gave an amplification result) genotype which was preliminarily named D21 (Table 1).

Table 1.
Results of the CoxBase comparison of the newly identified MLVA-genotype pattern (first line of the table).
Comparison with the stored genotype information of other C. burnetii strains in the database showed a close relation between the newly identified genotype herein and a chronic strain genotyped from Saudi Arabia, with 9 out of 11 perfect matches, in addition to another chronic Q fever isolate from France, matching with 10 out of 11 microsatellites (ms) markers (Figure 1).
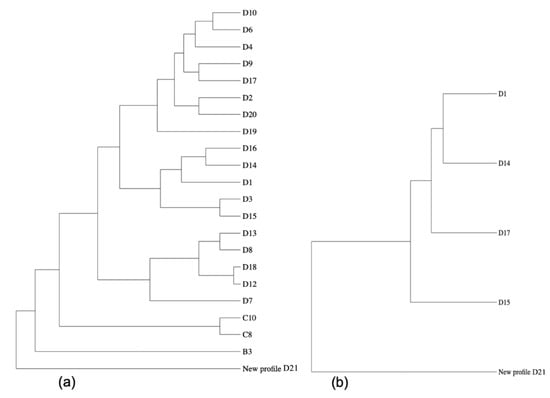
Figure 1.
Phylogenetic comparison of the MLVA-pattern of the C. burnetii identified in this study when analysed via the CoxBase-online platform (https://coxbase.q-gaps.de, accessed on 26 April 2021) using PhyD3 (https://phyd3.bits.vib.be/, accessed on 26 April 2021). (a) The new genotype (proposed name D21) is an outgroup due to missing microsatellite (ms)-markers, when maximal distance is not limited. (b) Nearest neighbours displayed when maximal distance value is limited to five.
3. Discussion
C. burnetii infection, known as Q fever, has been reported in domestic and wild animals, birds, and in a wide variety of tick species [11,19]. Moreover, C. burnetii infection is globally a major public health problem with infections being reported sporadically, as outbreaks or endemic disease [6,19,20,21]. However, little is known about C. burnetii in many resource-limited countries and this is also true for Africa, where limited diagnostic resources are resulting in a significant underestimation of the disease [22], although some studies have clearly demonstrated the importance of Q fever [23,24]. Therefore, this study investigated tick samples collected from camels in five states of Somalia for C. burnetii.
By screening for the C. burnetii multicopy target, the insertion sequence IS1111, more than half of the tested tick samples were Coxiella-positive (140/237, 59.1%). This impressive molecular prevalence result, when using a sensitive Taqman-probe-assay, could not—as expected—be confirmed with PCR’s using single copy gene targets, such as icd and com1. More than 92% (129 from 140) of the samples showed a Cycle threshold (Ct) higher than 33, representing the low amount of Coxiella DNA in the collected tick samples, resulting in dropouts of the single copy gene target PCRs. Although ticks are not the major source of infection, neither for animals nor humans [19], this finding is of high importance and is very remarkable. It clearly shows the high burden and probably of the endemicity of Coxiella spp. in the study area. It further highlights the need for additional epidemiological studies within the context of “One Health” (i.e., investigate samples from humans, domestic and wild animals, birds, and ticks). This will provide a better insight into the ecology and distribution of this important zoonotic pathogen. A few former studies conducted in Somalia reported the prevalence of C. burnetii, using serological methods, with a positivity ranging from 25% to 37% in humans and up to 12.1% in sheep and goats [4,16,17,18], whereas this study is the first of its kind reporting direct evidence of the existence of the C. burnetii pathogen. In other parts of the world, the prevalence of C. burnetii in ticks varied from 1.6% to 5.5% [19,25,26,27,28]. Nevertheless, in a study from Kenya, five out of 10 pools of Haemaphysalis leachi ticks collected from domestic dogs were found to be positive for C. burnetii [29]. Furthermore, a study in rural Senegal identified up to 37% IS1111-positive Amblyomma variegatum ticks [30].
Results of the genotyping using MLVA/VNTR analysis showed a new genotype that has never been reported before. This new genotype is closely related to a C. burnetii strain isolated in Saudi Arabia from a patient with chronic Q fever and heart valve infection [31]. The new genotype is also related to genotypes from France and Russia [32,33]. On one hand, this finding confirms that MLVA/VNTR typing is a powerful tool that can offer a good understanding of the molecular epidemiology of Coxiella spp. However, it ideally requires high quality and quantity DNA with a PCR cycle threshold/Ct of less than 33; in addition, not all markers behave equally according to their sensitivity in identification of microsatellite repeats. Furthermore, there is another challenge related to MLVA/VNTR typing, which is how to deal with missing microsatellite-marker results, as this influences the correlation of different typing patterns. Especially in samples with a low DNA-amount, very often no differentiation between true negative ms-marker positions and missing typing PCR-signals is possible. Therefore, the measuring of genetic distances between strains/isolates should primarily be based on identified tandem repeat marker patterns (see [33]). The MLVA-analysis method used there, and in the here presented study, fits best with the algorithm for highly polymorphic tandem repeat loci developed by Shriver et al. [34].
In this study, the observed C. burnetii-positive ticks were Hyalomma spp. (n = 138) and Amblyomma spp. (n = 2). This confirms the findings of Deavaux et al. [4] and Bellabidi et al. [35], who indicated that C. burnetii was mostly detected in Hyalomma spp. infesting dromedary camels. Nevertheless, C. burnetii has been detected in many other hard and soft ticks collected from humans, cattle, sheep, and goats, as well as from wild animals, such as deer, and from birds [5,26,27,28,29,30,36]. These ticks included, for instance, Ripicephalus spp., Amblyomma spp., Haemaphysalis spp., Ixodes spp., Dermacentor spp., and Ornithodoros spp.
In conclusion, this is the first report of the occurrence of C. burnetii in ticks collected from domestic camels in Somalia. The findings of the study show that this query agent might be endemic in the study area. All C. burnetii-positive ticks were Hyalomma spp., except for two Amblyomma spp. Future studies should investigate ticks, samples of blood, and milk from animals (e.g., cattle, sheep, goats, and camels), and samples from humans as well. Furthermore, characterization of C. burnetii isolates from Somalia are warranted to obtain more insights related to the pathogen specific genomic properties and molecular epidemiology.
4. Materials and Methods
4.1. Study Area
This study was conducted in five states of Somalia that include Mudug, Sanaag, Bari, Sool, and Nugaal (Figure 2). Bari and Nugaal are under the administration of Puntland State, which is bordered by Somaliland State to its west, the Gulf of Aden in the north, the Guardafui Channel in the northeast, the Indian Ocean in the southeast, the Galmudug region in the south, and the Somali region of Ethiopia in the southwest [37,38]. The study area is characterized by alternating wet and dry seasons with predictable but erratically distributed rainfall during the wet season [37,38]. The average rainfall is neither sufficient nor reliable enough to produce staple crops. Conversely, the interaction of climatic and ecological factors could only support livestock production, which is predominant throughout Somalia and contributes significantly to the national economy. Livestock in the investigated five states of Somalia are indigenous breeds of goats, sheep, camels, and cattle that are more adaptable to arid environment. The livestock production system is exclusively nomadic pastoralism, and rangeland is communal or free grazing throughout the country. Livestock numbers are subject to many factors that include vicious cycles of major droughts and disease epidemics [37,38]. The total number of animals in Puntland is estimated to be nearly 18,000,000 heads, including 10.7% camels, 2.3% cattle, 35.4% sheep, and 51.6% goats. However, no census has been carried out since 1988, and the above data is derived from an estimated growth rate of 0.07% in camels, 0.01% in cattle, 0.012% in goats, and 0.021% in sheep. With its strategic location at the Horn of Africa, Puntland remains one of the principal main hubs for livestock exportation in Somalia and connects the Gulf states, South and Central Somalia, Ethiopia, Kenya, and Yemen [37,38].
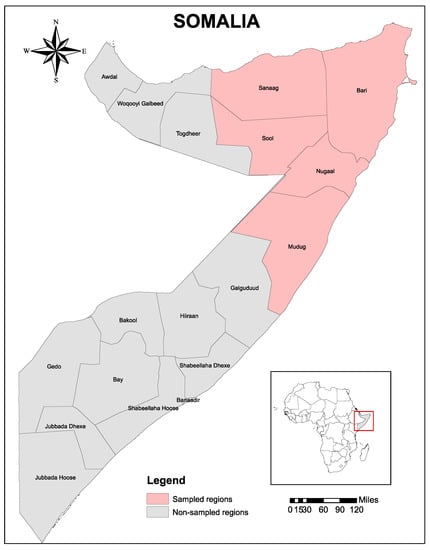
Figure 2.
Map of the study area. The sampled five regions, including Bari, Nugaal Mudug, Sool, and Sanaag, of Somalia are indicated in pink. The map was created using ArcGIS v. 10 (esri Inc., Redlands, CA, USA).
All investigated animals were domesticated camels, and generally, in Somalia, camels are raised for milk and meat. They also indicate social status and wealth and are used in events such as marriage to pay dowry.
4.2. Study Design and Tick Samples Collection
This cross-sectional study was conducted between January and March 2018 in five states of Somalia, namely Bari, Nugaal, Mudug, Sool, and Sanaag. Tick samples were collected from camels of different age groups during routine veterinary and animal health care activities in the field.
Attached ticks were collected from camels using a pair of blunt forceps. Prior to collection, each animal was cast and restrained. Collected ticks from the same animal were kept separately in a universal bottle containing 70% ethanol, labelled indicating location, sex, and age [39,40].
4.3. Laboratory Procedures
4.3.1. Tick Genera and Species Identification
Ticks were identified based on morphological characteristics according to Voltzit and Keirans [41], Apanaskevich and Horak [42,43,44], and Apanaskevich et al. [45].
Total nucleic acid was extracted using the MagNA Pure LC RNA/DNA Kit (Roche, Mannheim, Germany) in a MagNA Pure LC instrument (Roche), according to the manufacturer’s instructions. DNA was extracted from individual ticks or from pools containing 2 to 10 ticks per pool if ticks share the same developmental stage and species and had been collected from the same animal. The extracted total nucleic acid was stored at −80 °C until use.
When the morphology of a tick was not useful for identification, e.g., in the case of damage or if they were fully engorged ticks, the extracted DNA was used instead. The 16S rRNA gene was amplified using polymerase chain reaction (PCR) protocols and sequenced as described by Mangold et al. [46].
4.3.2. Detection of Coxiella burnetii
The extracted total nucleic acid was further used to detect the presence of C. burnetii in ticks. Using different genomic targets, the samples were screened for C. burnetii and Coxiella-like endosymbionts [47,48]. After screening with IS1111-PCR, further Coxiella specific icd and Com1-target real-time PCR assays were done [36]. Positive results were confirmed with a species-specific 16S-rRNA PCR with sequencing and differentiation by performing a blast-analysis in the NCBI database [36]. Further genomic differentiation and typing were done with adaA-gene and plasmid-type identification [49].
4.3.3. Molecular Characterization of Coxiella burnetii
Samples with enough DNA quality and quantity were genotyped with a 14-marker Multi-Locus Variable-Number of Tandem Repeats (MLVA/VNTR) analysis [26]. Identification and comparison of MLVA-genotype were done in the recent established online Coxiella-specific genotyping platform CoxBase (https://coxbase.q-gaps.de, accessed on 26 April 2021) [50]. This collection contains all of the known and published MLVA-genotypes of C. burnetii. The comparison is based on an algorithm for highly polymorphic tandem repeat loci developed by Shriver et al. [34]. Visualisation of phylogeny was done with PhyD3 (https://phyd3.bits.vib.be/, accessed on 26 April 2021), and to generate a breakdown of the strain relationship the maximal distance was limited to five. This means that only isolates of the CoxBase with up to five ms-loci having a different number of repeats to our MLVA-pattern were included in the analysis (excluding missing values).
Author Contributions
Conceptualization, D.F., A.S.S., L.C.-D. and Y.A.S.; methodology, D.F., L.C.-D. and Y.A.S.; validation, D.F. and L.C.-D.; formal analysis, D.F., A.Y.O., L.C.-D. and Y.A.S.; investigation and sampling, A.S.S.; resources, D.F., A.S.S., C.K., A.Y.O., L.C.-D. and Y.A.S.; data curation, D.F., L.C.-D. and Y.A.S.; writing—original draft preparation, D.F. and Y.A.S.; writing—review and editing, D.F., A.S.S., C.K., A.Y.O., L.C.-D. and Y.A.S.; visualization, D.F. and A.Y.O. All authors have read and agreed to the published version of the manuscript.
Funding
This study was partially funded by the German Ministry of Education and Research (Grant-No.: FKZ01KI1726E).
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the Ethics Review Committee (ERC) of the Department of Livestock Extension Services (DELES), reviewed by the Ministry of Livestock and Animal Husbandry (MoLAH) (Ref. MoLAH/DELES/029/2017, 15 September 2017).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
A.Y.O. is a member of the Pan African Network for Rapid Research, Response, and Preparedness for Infectious Diseases Epidemics consortium (PANDORA-ID-NET), supported by the European and Developing Countries Clinical Trials Partnership (EDCTP2) program (RIA2016E1609).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Maurin, M.; Raoult, D. Q Fever. Clin. Microbiol. Rev. 1999, 12, 518–553. [Google Scholar] [CrossRef] [PubMed]
- Kazar, J. Coxiella burnetii Infection. Ann. N. Y. Acad. Sci. 2005, 1063, 105–114. [Google Scholar] [CrossRef]
- Angelakis, E.; Raoult, D. Q Fever. Vet. Microbiol. 2010, 140, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Devaux, C.A.; Osman, I.O.; Million, M.; Raoult, D. Coxiella burnetii in Dromedary Camels (Camelus Dromedarius): A Possible Threat for Humans and Livestock in North Africa and the Near and Middle East? Front. Vet. Sci. 2020, 7, 558481. [Google Scholar] [CrossRef]
- Duron, O.; Sidi-Boumedine, K.; Rousset, E.; Moutailler, S.; Jourdain, E. The Importance of Ticks in Q Fever Transmission: What Has (and Has Not) Been Demonstrated? Trends Parasitol. 2015, 31, 536–552. [Google Scholar] [CrossRef] [PubMed]
- Klemmer, J.; Njeru, J.; Emam, A.; El-Sayed, A.; Moawad, A.A.; Henning, K.; Elbeskawy, M.A.; Sauter-Louis, C.; Straubinger, R.K.; Neubauer, H.; et al. Q Fever in Egypt: Epidemiological Survey of Coxiella burnetii Specific Antibodies in Cattle, Buffaloes, Sheep, Goats and Camels. PLoS ONE 2018, 13, e0192188. [Google Scholar] [CrossRef] [PubMed]
- Sahu, R.; Rawool, D.B.; Vinod, V.K.; Malik, S.V.S.; Barbuddhe, S.B. Current Approaches for the Detection of Coxiella burnetii Infection in Humans and Animals. J. Microbiol. Methods 2020, 179, 106087. [Google Scholar] [CrossRef]
- Gürtler, L.; Bauerfeind, U.; Blümel, J.; Burger, R.; Drosten, C.; Gröner, A.; Heiden, M.; Hildebrandt, M.; Jansen, B.; Offergeld, R.; et al. Coxiella Burnetii—Pathogenic Agent of Q (Query) Fever. Transfus. Med. Hemother. 2014, 41, 60–72. [Google Scholar] [CrossRef]
- Gale, P.; Kelly, L.; Mearns, R.; Duggan, J.; Snary, E.L. Q Fever through Consumption of Unpasteurised Milk and Milk Products—A Risk Profile and Exposure Assessment. J. Appl. Microbiol. 2015, 118, 1083–1095. [Google Scholar] [CrossRef]
- Hogema, B.M.; Slot, E.; Molier, M.; Schneeberger, P.M.; Hermans, M.H.; van Hannen, E.J.; van der Hoek, W.; Cuijpers, H.T.; Zaaijer, H.L. Coxiella burnetii Infection among Blood Donors during the 2009 Q-Fever Outbreak in the Netherlands. Transfusion 2012, 52, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Million, M.; Raoult, D. Recent Advances in the Study of Q Fever Epidemiology, Diagnosis and Management. J. Infect. 2015, 71, S2–S9. [Google Scholar] [CrossRef] [PubMed]
- Wegdam-Blans, M.C.A.; Vainas, T.; van Sambeek, M.R.; Cuypers, P.W.; Tjhie, H.T.J.; van Straten, A.H.M.; Teijink, J.A. Vascular Complications of Q-Fever Infections. Eur. J. Vasc. Endovasc. Surg. 2011, 42, 384–392. [Google Scholar] [CrossRef] [PubMed]
- Broos, P.P.H.L.; Hagenaars, J.C.J.P.; Kampschreur, L.M.; Wever, P.C.; Bleeker-Rovers, C.P.; Koning, O.H.J.; Teijink, J.A.W.; Wegdam-Blans, M.C.A. Vascular Complications and Surgical Interventions after World’s Largest Q Fever Outbreak. J. Vasc. Surg. 2015, 62, 1273–1280. [Google Scholar] [CrossRef] [PubMed]
- Wegdam-Blans, M.C.A.; Kampschreur, L.M.; Delsing, C.E.; Bleeker-Rovers, C.P.; Sprong, T.; van Kasteren, M.E.E.; Notermans, D.W.; Renders, N.H.M.; Bijlmer, H.A.; Lestrade, P.J.; et al. Chronic Q Fever: Review of the Literature and a Proposal of New Diagnostic Criteria. J. Infect. 2012, 64, 247–259. [Google Scholar] [CrossRef]
- Abdullah, H.H.A.M.; Hussein, H.A.; El-Razik, K.A.A.; Barakat, A.M.A.; Soliman, Y.A. Q Fever: A Neglected Disease of Camels in Giza and Cairo Provinces, Egypt. Vet. World 2019, 12, 1945–1950. [Google Scholar] [CrossRef]
- Schmatz, H.D.; Krauss, H.; Viertel, P.; Ismail, A.S.; Hussein, A.A. Seroepidemiological investigations in domestic ruminants from Egypt, Somalia and Jordan for the demonstration of complement fixing antibodies against Rickettsia and Chlamydia (author’s transl). Acta Trop. 1978, 35, 101–111. [Google Scholar] [PubMed]
- Gray, G.C.; Rodier, G.R.; Matras-Maslin, V.C.; Honein, M.A.; Ismail, E.A.; Botros, B.A.M.; Soliman, A.K.; Merrell, B.R.; Wang, S.P.; Grayston, J.T. Serologic Evidence of Respiratory and Rickettsial Infections among Somali Refugees. Am. J. Trop. Med. Hyg. 1995, 52, 349–353. [Google Scholar] [CrossRef]
- Botros, B.A.; Soliman, A.K.; Salib, A.W.; Olson, J.; Hibbs, R.G.; Williams, J.C.; Darwish, M.; el Tigani, A.; Watts, D.M. Coxiella burnetii Antibody Prevalences among Human Populations in North-East Africa Determined by Enzyme Immunoassay. J. Trop. Med. Hyg. 1995, 98, 173–178. [Google Scholar]
- Knap, N.; Žele, D.; Glinšek Biškup, U.; Avšič-Županc, T.; Vengušt, G. The Prevalence of Coxiella burnetii in Ticks and Animals in Slovenia. BMC Vet. Res. 2019, 15, 368. [Google Scholar] [CrossRef]
- Roest, H.I.J.; Bossers, A.; van Zijderveld, F.G.; Rebel, J.M.L. Clinical Microbiology of Coxiella Burnetii and Relevant Aspects for the Diagnosis and Control of the Zoonotic Disease Q Fever. Vet. Q. 2013, 33, 148–160. [Google Scholar] [CrossRef] [PubMed]
- van Roeden, S.E.; van Houwelingen, F.; Donkers, C.M.J.; Hogewoning, S.J.; de Lange, M.M.A.; van der Hoek, W.; Kampschreur, L.M.; Bonten, M.J.M.; Hoepelman, A.I.M.; Bleeker-Rovers, C.P.; et al. Exposure to Coxiella burnetii and Risk of Non-Hodgkin Lymphoma: A Retrospective Population-Based Analysis in the Netherlands. Lancet Haematol. 2018, 5, e211–e219. [Google Scholar] [CrossRef]
- Salifu, S.P.; Bukari, A.-R.A.; Frangoulidis, D.; Wheelhouse, N. Current Perspectives on the Transmission of Q Fever: Highlighting the Need for a Systematic Molecular Approach for a Neglected Disease in Africa. Acta Trop. 2019, 193, 99–105. [Google Scholar] [CrossRef]
- Njeru, J.; Henning, K.; Pletz, M.W.; Heller, R.; Neubauer, H. Q Fever Is an Old and Neglected Zoonotic Disease in Kenya: A Systematic Review. BMC Public Health 2016, 16, 297. [Google Scholar] [CrossRef]
- Prabhu, M.; Nicholson, W.L.; Roche, A.J.; Kersh, G.J.; Fitzpatrick, K.A.; Oliver, L.D.; Massung, R.F.; Morrissey, A.B.; Bartlett, J.A.; Onyango, J.J.; et al. Q Fever, Spotted Fever Group, and Typhus Group Rickettsioses among Hospitalized Febrile Patients in Northern Tanzania. Clin. Infect. Dis. 2011, 53, e8–e15. [Google Scholar] [CrossRef]
- Pilloux, L.; Baumgartner, A.; Jaton, K.; Lienhard, R.; Ackermann-Gäumann, R.; Beuret, C.; Greub, G. Prevalence of Anaplasma phagocytophilum and Coxiella burnetii in Ixodes ricinus Ticks in Switzerland: An Underestimated Epidemiologic Risk. New Microbes New Infect. 2019, 27, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, R.C.; Echaide, I.E.; Alves, R.N.; Beletti, M.E.; Nava, S.; Labruna, M.B. Coxiella burnetii in Ticks, Argentina. Emerg. Infect. Dis. 2013, 19, 344–346. [Google Scholar] [CrossRef] [PubMed]
- Tokarevich, N.K.; Panferova, Y.A.; Freylikhman, O.A.; Blinova, O.V.; Medvedev, S.G.; Mironov, S.V.; Grigoryeva, L.A.; Tretyakov, K.A.; Dimova, T.; Zaharieva, M.M.; et al. Coxiella burnetii in Ticks and Wild Birds. Ticks Tick-Borne Dis. 2019, 10, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Koka, H.; Sang, R.; Kutima, H.L.; Musila, L. Coxiella burnetii Detected in Tick Samples from Pastoral Communities in Kenya. BioMed Res. Int. 2018, 2018, 8158102. [Google Scholar] [CrossRef]
- Knobel, D.L.; Maina, A.N.; Cutler, S.J.; Ogola, E.; Feikin, D.R.; Junghae, M.; Halliday, J.E.B.; Richards, A.L.; Breiman, R.F.; Cleaveland, S.; et al. Coxiella burnetii in Humans, Domestic Ruminants, and Ticks in Rural Western Kenya. Am. J. Trop. Med. Hyg. 2013, 88, 513–518. [Google Scholar] [CrossRef]
- Mediannikov, O.; Fenollar, F.; Socolovschi, C.; Diatta, G.; Bassene, H.; Molez, J.-F.; Sokhna, C.; Trape, J.-F.; Raoult, D. Coxiella burnetii in Humans and Ticks in Rural Senegal. PLoS Negl. Trop. Dis. 2010, 4, e654. [Google Scholar] [CrossRef]
- Al-Hajjar, S.; Hussain Qadri, S.M.; Al-Sabban, E.; Jäger, C. Coxiella burnetii Endokarditis in a Child. Pediatr. Infect. Dis. J. 1997, 16, 911–913. [Google Scholar] [CrossRef] [PubMed]
- Arricau-Bouvery, N.; Hauck, Y.; Bejaoui, A.; Frangoulidis, D.; Bodier, C.C.; Souriau, A.; Meyer, H.; Neubauer, H.; Rodolakis, A.; Vergnaud, G. Molecular Characterization of Coxiella burnetii Isolates by Infrequent Restriction Site-PCR and MLVA Typing. BMC Microbiol. 2006, 6, 38. [Google Scholar] [CrossRef] [PubMed]
- Frangoulidis, D.; Walter, M.C.; Antwerpen, M.; Zimmermann, P.; Janowetz, B.; Alex, M.; Böttcher, J.; Henning, K.; Hilbert, A.; Ganter, M.; et al. Molecular Analysis of Coxiella burnetii in Germany Reveals Evolution of Unique Clonal Clusters. Int. J. Med. Microbiol. IJMM 2014, 304, 868–876. [Google Scholar] [CrossRef]
- Shriver, M.D.; Jin, L.; Boerwinkle, E.; Deka, R.; Ferrell, R.E.; Chakraborty, R. A Novel Measure of Genetic Distance for Highly Polymorphic Tandem Repeat Loci. Mol. Biol. Evol. 1995, 12, 914–920. [Google Scholar]
- Bellabidi, M.; Benaissa, M.H.; Bissati-Bouafia, S.; Harrat, Z.; Brahmi, K.; Kernif, T. Coxiella Burnetii in Camels (Camelus Dromedarius) from Algeria: Seroprevalence, Molecular Characterization, and Ticks (Acari: Ixodidae) Vectors. Acta Trop. 2020, 206, 105443. [Google Scholar] [CrossRef]
- Al-Deeb, M.A.; Frangoulidis, D.; Walter, M.C.; Kömpf, D.; Fischer, S.F.; Petney, T.; Muzaffar, S.B. Coxiella-like Endosymbiont in Argasid Ticks (Ornithodoros Muesebecki) from a Socotra Cormorant Colony in Umm Al Quwain, United Arab Emirates. Ticks Tick-Borne Dis. 2016, 7, 166–171. [Google Scholar] [CrossRef] [PubMed]
- McKinney, P. Puntland Food Security and Vulnerability Assessment April 2007; 2007. Available online: https://documents.wfp.org/stellent/groups/public/documents/ena/wfp144576.pdf (accessed on 8 April 2021).
- PLHDS. Puntland Report 2020 Somali Health and Demographic Survey; 2020. Available online: http://pl.statistics.so/wp-content/uploads/2020/11/Puntland_SHDS-Report-2020.pdf, (accessed on 8 April 2021).
- Shuaib, Y.A.; Elhag, A.M.-A.W.; Brima, Y.A.; Abdalla, M.A.; Bakiet, A.O.; Mohmed-Noor, S.E.-T.; Lemhöfer, G.; Bestehorn, M.; Poppert, S.; Schaper, S.; et al. Ixodid Tick Species and Two Tick-Borne Pathogens in Three Areas in the Sudan. Parasitol. Res. 2020, 119, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Springer, A.; Shuaib, Y.A.; Isaa, M.H.; Ezz-Eldin, M.I.-E.; Osman, A.Y.; Yagoub, I.A.; Abdalla, M.A.; Bakiet, A.O.; Mohmed-Noor, S.E.-T.; Schaper, S.; et al. Tick Fauna and Associated Rickettsia, Theileria, and Babesia spp. in Domestic Animals in Sudan (North Kordofan and Kassala States). Microorganisms 2020, 8, 1969. [Google Scholar] [CrossRef]
- Voltzit, O.V.; Keirans, J.E. A Review of African Amblyomma Species (Acari, Ixodida, Ixodidae). In Acarina 112135-214, 2003. Available online: http://publikationen.ub.uni-frankfurt.de/frontdoor/index/index/docId/11904, (accessed on 5 February 2021).
- Apanaskevich, D.A.; Horak, I.G. The Genus Hyalomma. VI. Systematics of H. (Euhyalomma) truncatum and the Closely Related Species, H. (E.) albiparmatum and H. (E.) nitidum (Acari: Ixodidae). Exp. Appl. Acarol. 2008, 44, 115–136. [Google Scholar] [CrossRef]
- Apanaskevich, D.A.; Horak, I.G. The Genus Hyalomma Koch, 1844: V. Re-Evaluation of the Taxonomic Rank of Taxa Comprising the H. (Euhyalomma) marginatum Koch Complex of Species (Acari: Ixodidae) with Redescription of All Parasitic Stages and Notes on Biology. Int. J. Acarol. 2008, 34, 13–42. [Google Scholar] [CrossRef]
- Apanaskevich, D.A.; Horak, I.G. The Genus Hyalomma Koch, 1844. IX. Redescription of All Parasitic Stages of H. (Euhyalomma) impeltatum Schulze & Schlottke, 1930 and H. (E.) somalicum Tonelli Rondelli, 1935 (Acari: Ixodidae). Syst. Parasitol. 2009, 73, 199–218. [Google Scholar] [CrossRef] [PubMed]
- Apanaskevich, D.A.; Schuster, A.L.; Horak, I.G. The Genus Hyalomma: VII. Redescription of All Parasitic Stages of H. (Euhyalomma) dromedarii and H. (E.) schulzei (Acari: Ixodidae). J. Med. Entomol. 2008, 45, 817–831. [Google Scholar] [CrossRef] [PubMed]
- Mangold, A.J.; Bargues, M.D.; Mas-Coma, S. Mitochondrial 16S rDNA Sequences and Phylogenetic Relationships of Species of Rhipicephalus and Other Tick Genera among Metastriata (Acari: Ixodidae). Parasitol. Res. 1998, 84, 478–484. [Google Scholar] [CrossRef] [PubMed]
- Duron, O. The IS1111 Insertion Sequence Used for Detection of Coxiella burnetii Is Widespread in Coxiella-like Endosymbionts of Ticks. FEMS Microbiol. Lett. 2015, 362, fnv132. [Google Scholar] [CrossRef]
- Jourdain, E.; Duron, O.; Barry, S.; González-Acuña, D.; Sidi-Boumedine, K. Molecular Methods Routinely Used to Detect Coxiella burnetii in Ticks Cross-React with Coxiella-like Bacteria. Infect. Ecol. Epidemiol. 2015, 5, 29230. [Google Scholar] [CrossRef]
- Frangoulidis, D.; Splettstoesser, W.D.; Landt, O.; Dehnhardt, J.; Henning, K.; Hilbert, A.; Bauer, T.; Antwerpen, M.; Meyer, H.; Walter, M.C.; et al. Microevolution of the Chromosomal Region of Acute Disease Antigen A (AdaA) in the Query (Q) Fever Agent Coxiella burnetii. PLoS ONE 2013, 8, e53440. [Google Scholar] [CrossRef]
- Fasemore, A.M.; Helbich, A.; Walter, M.C.; Dandekar, T.; Vergnaud, G.; Förstner, K.U.; Frangoulidis, D. CoxBase: An Online Platform for Epidemiological Surveillance, Visualization, Analysis and Typing of Coxiella burnetii Genomic Sequence. bioRxiv 2020. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).