
The Usefulness of a Duplex RT-qPCR during the Recent Yellow Fever Brazilian Epidemic: Surveillance of Vaccine Adverse Events, Epizootics and Vectors

 , ,
, ,  ,
,  , and
, and
Abstract
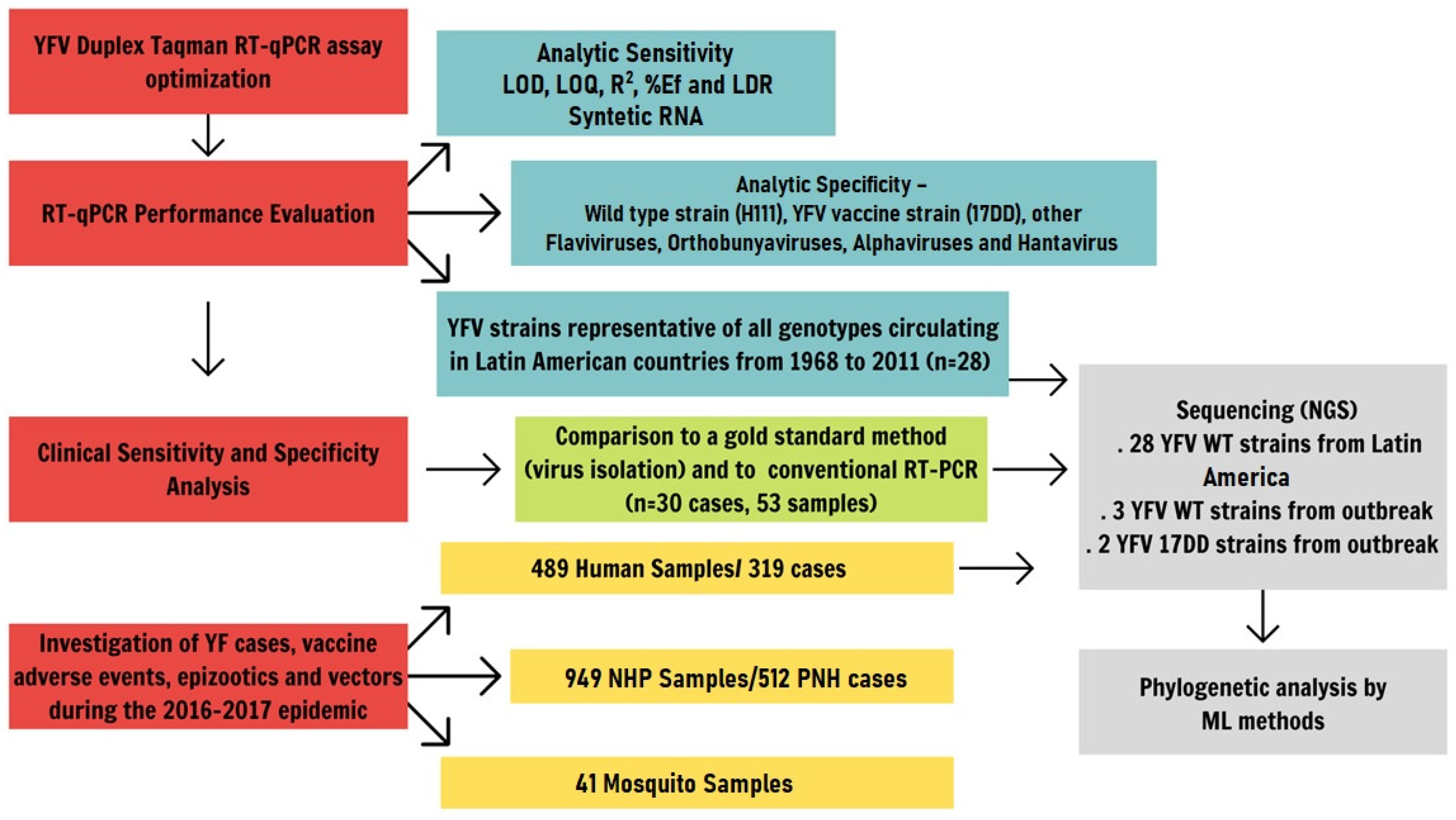
:1. Introduction
2. Results
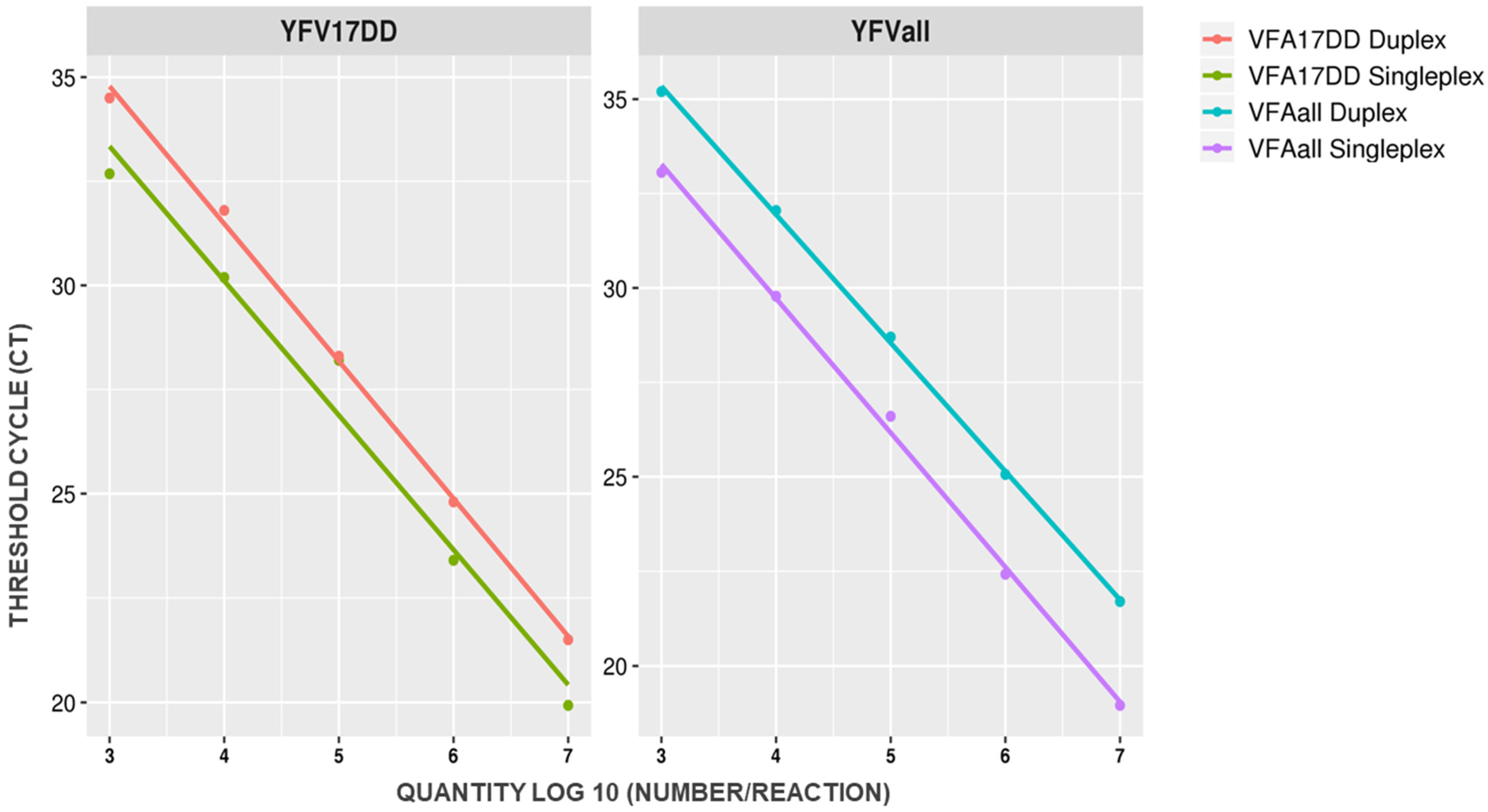
2.1. YFV Duplex Taqman RT-qPCR Assay Standardization
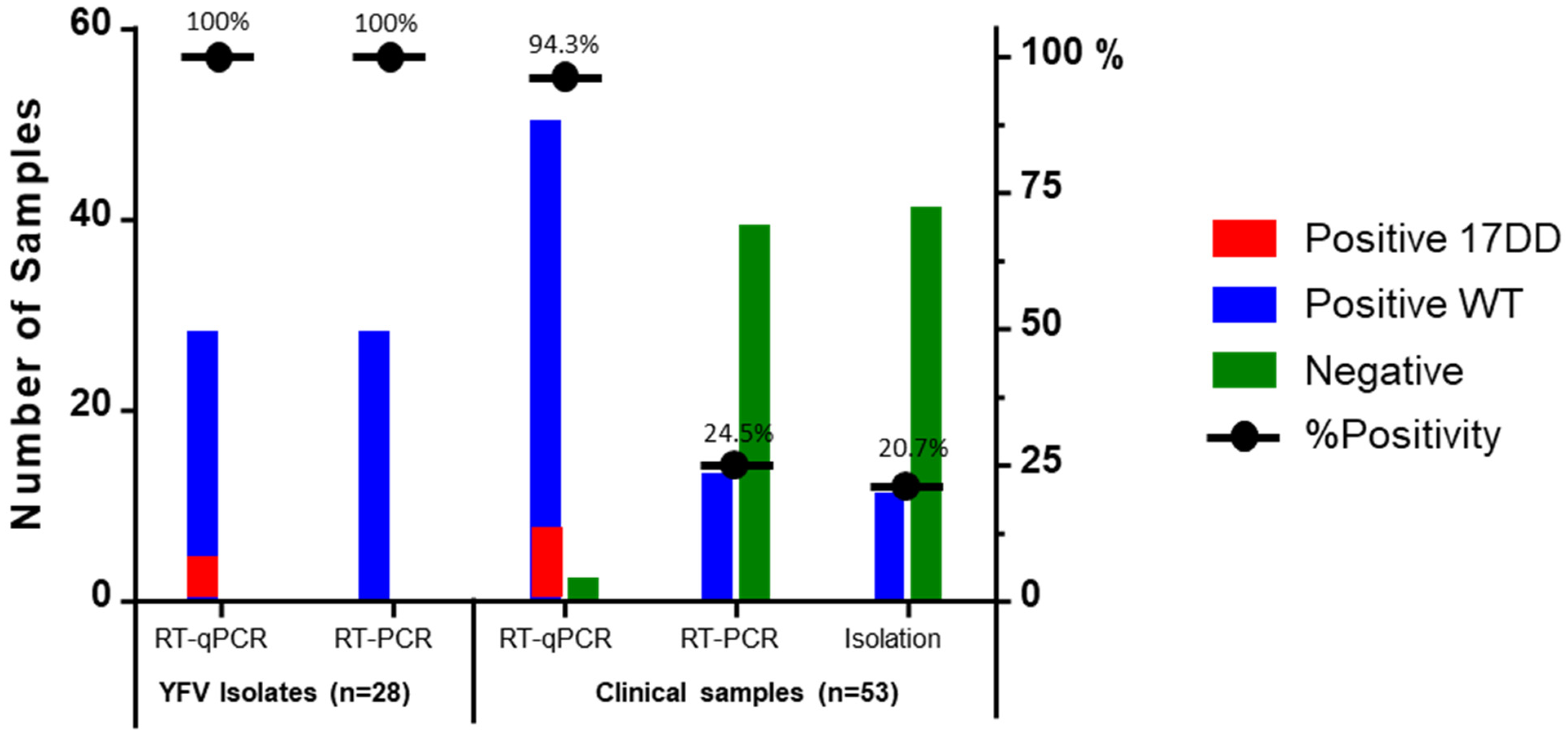
2.2. Duplex Taqman RT-qPCR Assay Clinical Evaluation
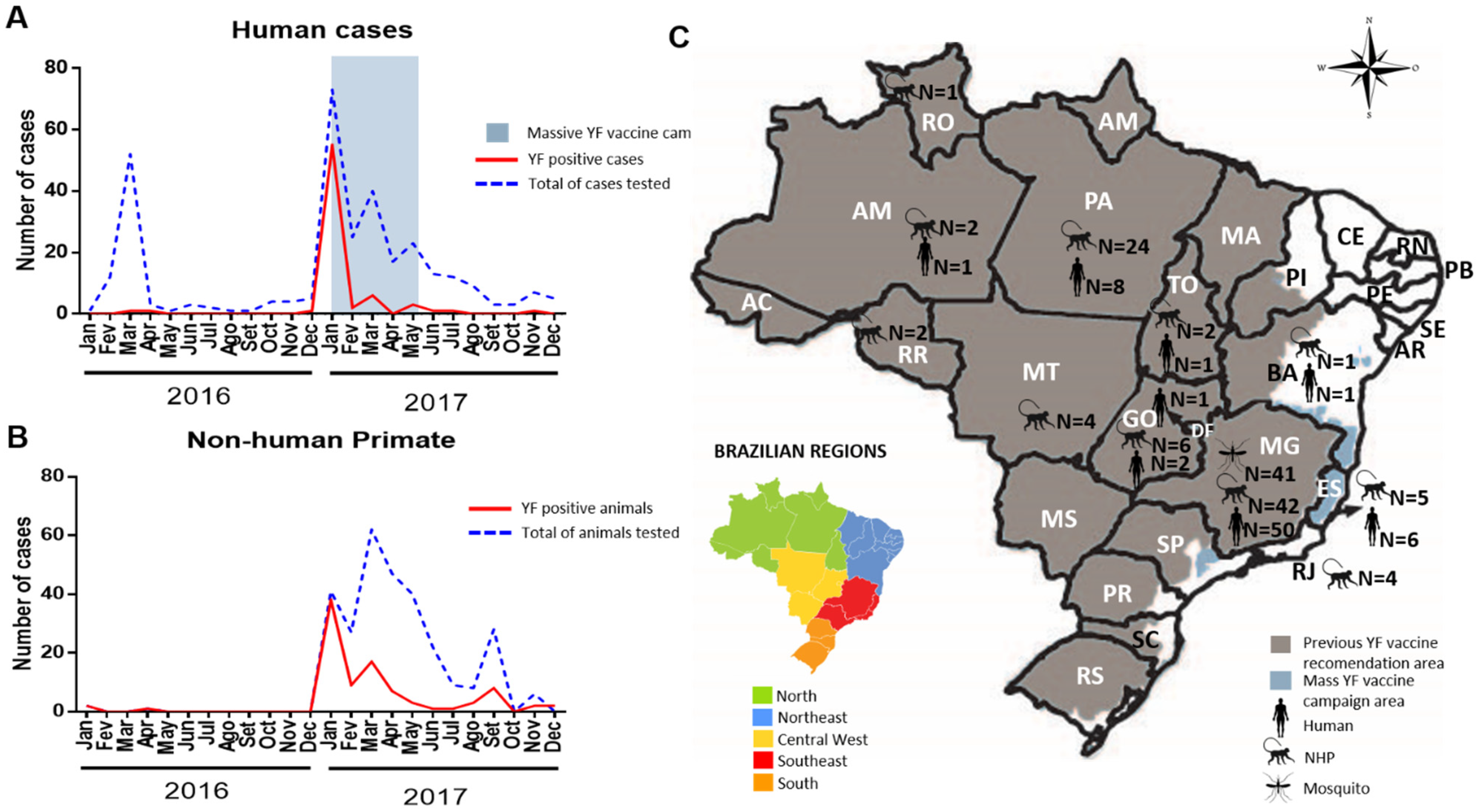
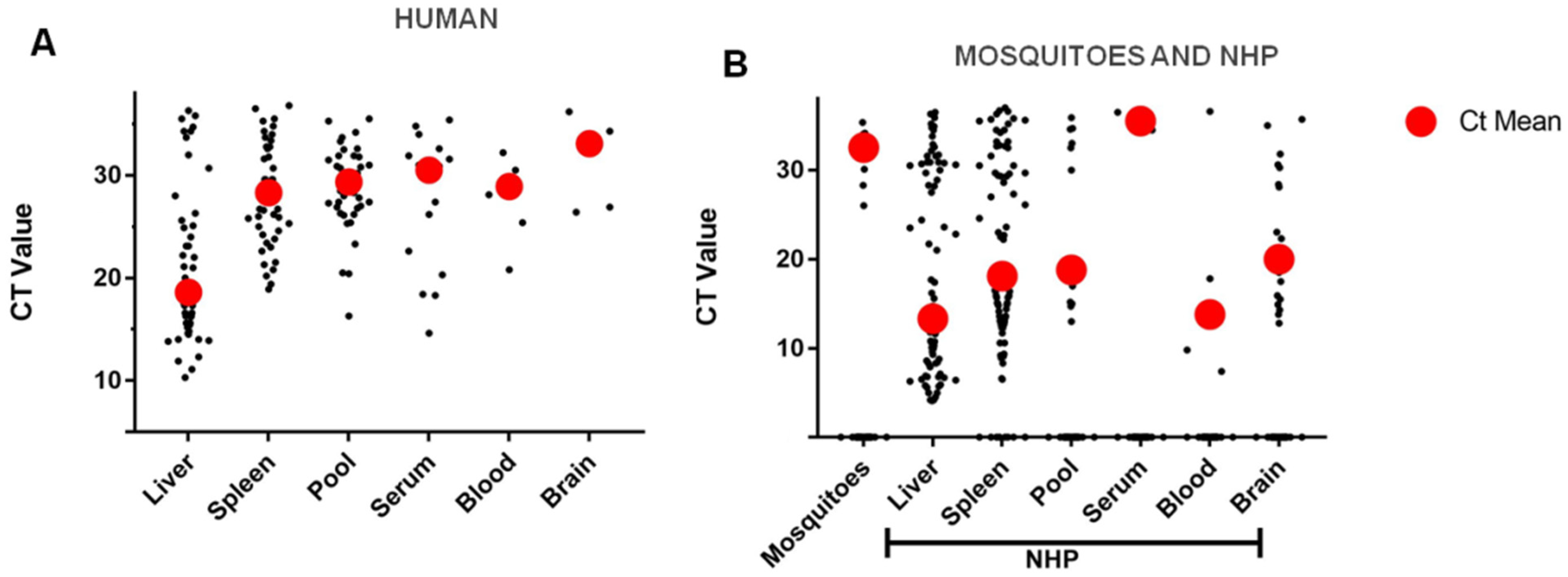
2.3. Investigation of YF Cases, Vaccine Adverse Events, Epizootics and Vectors during the 2016–2017 Epidemic
2.4. Phylogenetic Analysis
3. Discussion
4. Materials and Methods
4.1. Ethical Statement
4.2. Duplex TaqMan RT-qPCR Assay Establishment
4.3. Duplex TaqMan RT-qPCR Assay Evaluation
4.4. Investigation of YF Cases, Vaccine Adverse Events, Vectors and Epizootics during the 2016–2017 Epidemic
4.5. Next Generation Sequencing
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Chambers, T.J.; Hahn, C.S.; Galler, R.; Rice, C.M. Flavivirus Genome Organization, Expression, and Replication. Annu. Rev. Microbiol. 1990, 44, 649–688. [Google Scholar] [CrossRef]
- Lefkowitz, E.J.; Dempsey, D.M.; Hendrickson, R.C.; Orton, R.J.; Siddell, S.G.; Smith, D.B. Virus taxonomy: The database of the International Committee on Taxonomy of Viruses (ICTV). Nucleic Acids Res. 2018, 46, D708–D717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasconcelos, P.F.C.; Bryant, J.E.; da Rosa, A.P.A.T.; Tesh, R.B.; Rodrigues, S.G.; Barrett, A.D.T. Genetic Divergence and Dispersal of Yellow Fever Virus, Brazil. Emerg. Infect. Dis. 2004, 10, 1578–1584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monath, T.P.; Vasconcelos, P.F.C. Yellow fever. J. Clin. Virol. 2015, 64, 160–173. [Google Scholar] [CrossRef] [PubMed]
- Paules, C.I.; Fauci, A.S. Yellow Fever—Once Again on the Radar Screen in the Americas. N. Engl. J. Med. 2017, 376, 1397–1399. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.H.; Hamer, D.H. Vaccination Strategies during Shortages of Yellow Fever Vaccine—Reply. JAMA 2018, 319, 1280. [Google Scholar] [CrossRef]
- Silva, N.I.O.; Sacchetto, L.; de Rezende, I.M.; Trindade, G.d.S.; LaBeaud, A.D.; de Thoisy, B.; Drumond, B.P. Recent sylvatic yellow fever virus transmission in Brazil: The news from an old disease. Virol. J. 2020, 17, 9. [Google Scholar] [CrossRef] [Green Version]
- Grobbelaar, A.A.; Weyer, J.; Moolla, N.; van Vuren, P.J.; Moises, F.; Paweska, J.T. Resurgence of Yellow Fever in Angola, 2015–2016. Emerg. Infect. Dis. 2016, 22, 1854–1855. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Stepping up local efforts to stop global spread of yellow fever. Bull. World Health Organ. 2018, 96, 374–375. [Google Scholar] [CrossRef]
- Vasconcelos, P.F.C. Febre amarela: A doença ea vacina, uma história inacabada; Yellow fever: The disease and the vaccine, an unfinished history. Rev. Soc. Bras. Med. Trop. 2003, 36, 275–293. Available online: http://bases.bireme.br/cgi-bin/wxislind.exe/iah/online/?IsisScript=iah/iah.xis&src=google&base=LILACS&lang=p&nextAction=lnk&exprSearch=290004&indexSearch=ID (accessed on 25 March 2021). [CrossRef]
- Gossner, C.M.; Haussig, J.M.; Lary, C.D.B.D.S.; Aaslav, K.K.; Schlagenhauf, P.; Sudre, B. Increased risk of yellow fever infections among unvaccinated European travellers due to ongoing outbreak in Brazil, July 2017 to March 2018. Eurosurveillance 2018, 23, 18-00106. [Google Scholar] [CrossRef] [Green Version]
- Ministério da Saúde Brasil. Boletim Epidemiológico Secretaria—Emergência Epidemiológica de Febre Amarela No Brasil, No Período de Dezembro de 2016 a Junho de 2018; Ministério da Saúde: Brasilia, Brazil, 2018.
- Callender, D.M. Management and control of yellow fever virus: Brazilian outbreak January–April 2018. Glob. Public Health 2019, 14, 445–455. [Google Scholar] [CrossRef]
- Litvoc, M.N.; Novaes, C.T.G.; Lopes, M.I.B.F. Yellow fever. Rev. Assoc. Med. Bras. 2018, 64, 106–113. [Google Scholar] [CrossRef] [Green Version]
- Gardner, C.L.; Ryman, K.D. Yellow Fever: A Reemerging Threat. Clin. Lab. Med. 2010, 30, 237–260. [Google Scholar] [CrossRef] [Green Version]
- Pan American Health Organization/World Health Organization. 10 July 2017: Epidemiological Update: Yellow Fever; PAHO/WHO: Washington, DC, USA, 2017. [Google Scholar]
- Ministério da Saúde Brasil. Febre Amarela: Guia Para Profissionais de Saúde; Ministério da Saúde: Brasilia, Brazil, 2017.
- Vasconcelos, P.F.; Luna, E.J.; Galler, R.; Silva, L.J.; Coimbra, T.L.; Barros, V.L.; Monath, T.P.; Rodigues, S.G.; Laval, C.; Costa, Z.G.; et al. Serious adverse events associated with yellow fever 17DD vaccine in Brazil: A report of two cases. Lancet 2001, 358, 91–97. [Google Scholar] [CrossRef]
- Rossetto, E.V.; Angerami, R.N.; de Luna, E.J.A. What to expect from the 2017 yellow fever outbreak in Brazil? Rev. Inst. Med. Trop. Sao Paulo 2017, 59, 17–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benchimol, J.L. História da febre amarela no Brasil. História Ciências Saúde-Manguinhos 1994, 1, 121–124. [Google Scholar] [CrossRef] [Green Version]
- Vasconcelos, P.F.C. Yellow fever in Brazil: Thoughts and hypotheses on the emergence in previously free areas. Rev. Saude Publica 2010, 44, 1144–1149. [Google Scholar] [CrossRef] [Green Version]
- Ministério da Saúde Brasil; Secretaria de Vigilância em Saúde. Boletim Epidemiológico—Reemergência Da Febre Amarela Silvestre No Brasil, 2014/2015: Situação Epidemiológica e a Importância da Vacinação Preventiva e da Vigilância Intensificada, No Período Sazonal; Ministério da Saúde: Brasilia, Brazil, 2015.
- Ministério da Saúde Brasil. Informe 43/2017; Ministério da Saúde: Brasilia, Brazil, 2017.
- World Health Organization. Global Strategy to Eliminate Yellow Fever Epidemics (EYE); SAGE: Thousand Oaks, CA, USA, 2018. [Google Scholar]
- World Health Organization. Yellow fever in Africa and the Americas, 2016. Wkly. Epidemiol. Rec. 2017, 92, 442–452. [Google Scholar]
- Fox, J.P.; Lennette, E.H.; Manso, C.; Aguiar, J.R.S. Encephalitis in Man following Vaccination with 17D Yellow Fever Virus. Am. J. Hyg. 1942, 36, 117–142. [Google Scholar]
- Martin, M.; Tsai, T.F.; Cropp, B.; Chang, G.-J.J.; Holmes, D.A.; Tseng, J.; Shieh, W.-J.; Zaki, S.R.; Al-Sanouri, I.; Cutrona, A.F.; et al. Fever and multisystem organ failure associated with 17D-204 yellow fever vaccination: A report of four cases. Lancet 2001, 358, 98–104. [Google Scholar] [CrossRef]
- De Martins, R.M.; da Leal, M.L.F.; Homma, A. Serious adverse events associated with yellow fever vaccine. Hum. Vaccines Immunother. 2015, 11, 2183–2187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milazzo, M.L.; Cajimat, M.N.B.; Hanson, J.D.; Bradley, R.D.; Quintana, M.; Sherman, C.; Velásquez, R.T.; Fulhorst, C.F. Catacamas Virus, a Hantaviral Species Naturally Associated with Oryzomys Couesi (Coues’ Oryzomys) In Honduras. Am. J. Trop. Med. Hyg. 2006, 75, 1003–1010. [Google Scholar] [CrossRef]
- Negura, L.; Negura, A. Limits and pitfalls of SYBR Green detection in quantitative PCR. An. Ştiinţifice Univ. Alexandru Loan Cuza Iași Sect. II A Genet. Biol. Mol. 2012, 13–18. [Google Scholar]
- Fernandes-Monteiro, A.G.; Trindade, G.F.; Yamamura, A.M.Y.; Moreira, O.C.; de Paula, V.S.; Duarte, A.C.M.; Britto, C.; Lima, S.M.B. New approaches for the standardization and validation of a real-time qPCR assay using TaqMan probes for quantification of yellow fever virus on clinical samples with high quality parameters. Hum. Vaccines Immunother. 2015, 11, 1865–1871. [Google Scholar] [CrossRef] [Green Version]
- Bhat, S.; Herrmann, J.; Armishaw, P.; Corbisier, P.; Emslie, K.R. Single molecule detection in nanofluidic digital array enables accurate measurement of DNA copy number. Anal. Bioanal. Chem. 2009, 394, 457–467. [Google Scholar] [CrossRef]
- Corbisier, P.; Bhat, S.; Partis, L.; Xie, V.R.D.; Emslie, K.R. Absolute quantification of genetically modified MON810 maize (Zea mays L.) by digital polymerase chain reaction. Anal. Bioanal. Chem. 2010, 396, 2143–2150. [Google Scholar] [CrossRef]
- Whale, A.S.; Huggett, J.F.; Cowen, S.; Speirs, V.; Shaw, J.; Ellison, S.; Foy, C.A.; Scott, D.J. Comparison of microfluidic digital PCR and conventional quantitative PCR for measuring copy number variation. Nucleic Acids Res. 2012, 40, e82. [Google Scholar] [CrossRef]
- Whale, A.S.; Cowen, S.; Foy, C.A.; Huggett, J.F. Methods for Applying Accurate Digital PCR Analysis on Low Copy DNA Samples. PLoS ONE 2013, 8, e58177. [Google Scholar] [CrossRef] [Green Version]
- Rojas, A.; Diagne, C.T.; Stittleburg, V.D.; Mohamed-Hadley, A.; de Guillén, Y.A.; Balmaseda, A.; Faye, O.; Faye, O.; Sall, A.A.; Harris, E.; et al. Internally Controlled, Multiplex Real-Time Reverse Transcription PCR for Dengue Virus and Yellow Fever Virus Detection. Am. J. Trop. Med. Hyg. 2018, 98, 1833–1836. [Google Scholar] [CrossRef]
- Hoorfar, J.; Malorny, B.; Abdulmawjood, A.; Cook, N.; Wagner, M.; Fach, P. Practical Considerations in Design of Internal Amplification Controls for Diagnostic PCR Assays. J. Clin. Microbiol. 2004, 42, 1863–1868. [Google Scholar] [CrossRef] [Green Version]
- Santiago, G.A.; Vergne, E.; Quiles, Y.; Cosme, J.; Vazquez, J.; Medina, J.F.; Medina, F.; Colón, C.; Margolis, H.; Muñoz-Jordán, J.L. Analytical and Clinical Performance of the CDC Real Time RT-PCR Assay for Detection and Typing of Dengue Virus. PLoS Negl. Trop. Dis. 2013, 7, e2311. [Google Scholar] [CrossRef]
- Lanciotti, R.S.; Kosoy, O.L.; Laven, J.J.; Velez, J.O.; Lambert, A.J.; Johnson, A.J.; Stanfield, S.M.; Duffy, M.R. Genetic and Serologic Properties of Zika Virus Associated with an Epidemic, Yap State, Micronesia, 2007. Emerg. Infect. Dis. 2008, 14, 1232–1239. [Google Scholar] [CrossRef]
- Lanciotti, R.S.; Kosoy, O.L.; Laven, J.J.; Panella, A.J.; Velez, J.O.; Lambert, A.J.; Campbell, G.L. Chikungunya Virus in US Travelers Returning from India, 2006. Emerg. Infect. Dis. 2007, 13, 764–767. [Google Scholar] [CrossRef]
- Lanciotti, R.S.; Kerst, A.J.; Nasci, R.S.; Godsey, M.S.; Mitchell, C.J.; Savage, H.M.; Komar, N.; Panella, N.A.; Allen, B.C.; Volpe, K.E.; et al. Rapid Detection of West Nile Virus from Human Clinical Specimens, Field-Collected Mosquitoes, and Avian Samples by a TaqMan Reverse Transcriptase-PCR Assay. J. Clin. Microbiol. 2000, 38, 4066–4071. [Google Scholar] [CrossRef] [Green Version]
- Naveca, F.G.; Nascimento, V.A.D.; Souza, V.C.D.; de Nunes, B.T.; Rodrigues, D.S.G.; da Vasconcelos, P.F.C. Multiplexed reverse transcription real-time polymerase chain reaction for simultaneous detection of Mayaro, Oropouche, and Oropouche-like viruses. Memórias Inst. Oswaldo Cruz 2017, 112, 510–513. [Google Scholar] [CrossRef] [Green Version]
- Mantel, N.; Aguirre, M.; Gulia, S.; Girerd-Chambaz, Y.; Colombani, S.; Moste, C.; Barban, V. Standardized quantitative RT-PCR assays for quantitation of yellow fever and chimeric yellow fever–dengue vaccines. J. Virol. Methods 2008, 151, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Hughes, H.R.; Russell, B.J.; Mossel, E.C.; Kayiwa, J.; Lutwama, J.; Lambert, A.J. Development of a real-Time reverse transcription-PCR assay for global differentiation of yellow fever virus vaccine-Related adverse events from natural infections. J. Clin. Microbiol. 2018, 56, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galler, R.; Pugachev, K.; Santos, C.; Ocran, S.; Jabor, A.; Rodrigues, S.; Marchevsky, R.; Freire, M.; Almeida, L.; Cruz, A.; et al. Phenotypic and Molecular Analyses of Yellow Fever 17DD Vaccine Viruses Associated with Serious Adverse Events in Brazil. Virology 2001, 290, 309–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, C.; Torres, M.C.; Patel, P.; Moreira-Soto, A.; Gould, E.A.; Charrel, R.N.; de Lamballerie, X.; Nogueira, R.M.R.; Sequeira, P.C.; Rodrigues, C.D.S.; et al. Lineage-Specific Real-Time RT-PCR for Yellow Fever Virus Outbreak Surveillance, Brazil. Emerg. Infect. Dis. 2017, 23, 1867–1871. [Google Scholar] [CrossRef]
- Pulendran, B.; Oh, J.Z.; Nakaya, H.; Ravindran, R.; Kazmin, D.A. Immunity to viruses: Learning from successful human vaccines. Immunol. Rev. 2013, 255, 243–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quaresma, J.A.S.; Pagliari, C.; Medeiros, D.B.A.; Duarte, M.I.S.; Vasconcelos, P.F.C. Immunity and immune response, pathology and pathologic changes: Progress and challenges in the immunopathology of yellow fever. Rev. Med. Virol. 2013, 23, 305–318. [Google Scholar] [CrossRef]
- Lefeuvre, A.; Contamin, H.; Decelle, T.; Fournier, C.; Lang, J.; Deubel, V.; Marianneau, P. Host-cell interaction of attenuated and wild-type strains of yellow fever virus can be differentiated at early stages of hepatocyte infection. Microbes Infect. 2006, 8, 1530–1538. [Google Scholar] [CrossRef]
- Woodson, S.E.; Freiberg, A.N.; Holbrook, M.R. Coagulation factors, fibrinogen and plasminogen activator inhibitor-1, are differentially regulated by yellow fever virus infection of hepatocytes. Virus Res. 2013, 175, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Barba-Spaeth, G.; Longman, R.S.; Albert, M.L.; Rice, C.M. Live attenuated yellow fever 17D infects human DCs and allows for presentation of endogenous and recombinant T cell epitopes. J. Exp. Med. 2005, 202, 1179–1184. [Google Scholar] [CrossRef]
- Cong, Y.; McArthur, M.A.; Cohen, M.; Jahrling, P.B.; Janosko, K.B.; Josleyn, N.; Kang, K.; Zhang, T.; Holbrook, M.R. Characterization of Yellow Fever Virus Infection of Human and Non-human Primate Antigen Presenting Cells and Their Interaction with CD4+ T Cells. PLoS Negl. Trop. Dis. 2016, 10, e0004709. [Google Scholar] [CrossRef]
- McLinden, J.H.; Bhattarai, N.; Stapleton, J.T.; Chang, Q.; Kaufman, T.M.; Cassel, S.L.; Sutterwala, F.S.; Haim, H.; Houtman, J.C.; Xiang, J. Yellow Fever Virus, but Not Zika Virus or Dengue Virus, Inhibits T-Cell Receptor–Mediated T-Cell Function by an RNA-Based Mechanism. J. Infect. Dis. 2017, 216, 1164–1175. [Google Scholar] [CrossRef] [PubMed]
- Woodson, S.E.; Freiberg, A.N.; Holbrook, M.R. Differential cytokine responses from primary human Kupffer cells following infection with wild-type or vaccine strain yellow fever virus. Virology 2011, 412, 188–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khaiboullina, S.F.; Rizvanov, A.A.; Holbrook, M.R.; St Jeor, S. Yellow fever virus strains Asibi and 17D-204 infect human umbilical cord endothelial cells and induce novel changes in gene expression. Virology 2005, 342, 167–176. [Google Scholar] [CrossRef] [Green Version]
- Vallone, P.M.; Butler, J.M. AutoDimer: A screening tool for primer-dimer and hairpin structures. Biotechniques 2004, 37, 226–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rački, N.; Morisset, D.; Gutierrez-Aguirre, I.; Ravnikar, M. One-step RT-droplet digital PCR: A breakthrough in the quantification of waterborne RNA viruses. Anal. Bioanal. Chem. 2014, 406, 661–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menting, S.; Thai, K.T.D.; Nga, T.T.T.; Phuong, H.L.; Klatser, P.; Wolthers, K.C.; Binh, T.Q.; de Vries, P.J.; Beld, M. Internally Controlled, Generic Real-Time PCR for Quantification and Multiplex Real-Time PCR with Serotype-Specific Probes for Serotyping of Dengue Virus Infections. Adv. Virol. 2011, 2011, 514681. [Google Scholar] [CrossRef] [Green Version]
- Dreier, J.; Stormer, M.; Kleesiek, K. Use of Bacteriophage MS2 as an Internal Control in Viral Reverse Transcription-PCR Assays. J. Clin. Microbiol. 2005, 43, 4551–4557. [Google Scholar] [CrossRef] [Green Version]
- Svec, D.; Tichopad, A.; Novosadova, V.; Pfaffl, M.W.; Kubista, M. How good is a PCR efficiency estimate: Recommendations for precise and robust qPCR efficiency assessments. Biomol. Detect. Quantif. 2015, 3, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.; Leung, H.C.M.; Yiu, S.M.; Chin, F.Y.L. IDBA-UD: A de novo assembler for single-cell and metagenomic sequencing data with highly uneven depth. Bioinformatics 2012, 28, 1420–1428. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Huson, D.H.; Bryant, D. Application of Phylogenetic Networks in Evolutionary Studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PARAMETERS | SINGLEPLEX | DUPLEX | ||
|---|---|---|---|---|
| YFVall | YFV17DD | YFVall | YFV17DD | |
| Ct cut off | <38 | <38 | <38 | <38 |
| Slope | −3.359 | −3.226 | −3.314 | −3.304 |
| Y-inter | 42.856 | 46.86 | 45.039 | 44.778 |
| R2 | 0.993 | 0.993 | 0.997 | 0.997 |
| EF% | 98.48% | 104.1% | 100.34% | 100.76% |
| LDR | 103 to 107 copies/reaction | 103 to 107 copies/reaction | ||
| LOD | 102 copies/reaction | 102 copies/reaction | ||
| LOQ | 103 copies/reaction | 102 copies/reaction | ||
| Host | Specimen | Positive (%) |
|---|---|---|
| Human | Serum (n = 180) | 8.89 |
| Blood (n = 12) | 50.00 | |
| Liver (n = 117) | 45.30 | |
| Spleen (n = 72) | 69.44 | |
| Brain (n = 19) | 26.32 | |
| Pool (n = 83) | 48.19 | |
| Cerebrospinal Fluid (n = 6) | 0 | |
| TOTAL = 489 | ||
| NHP | Serum (n = 46) | 4.35 |
| Blood (n = 43) | 6.98 | |
| Liver (n = 315) | 26.98 | |
| Spleen (n = 339) | 22.71 | |
| Brain (n = 82) | 26.83 | |
| Pool (n = 124) | 10.48 | |
| TOTAL = 949 | ||
| Mosquitoes | Haemagogus spp. (n = 31) | 19.51 |
| Sabethes spp. (n = 10) | 0 | |
| TOTAL = 41 |
| Primers | Sequence (5′→3′) | Genome Position b |
| YFALL R | 5′- CTG CTA ATC GCT CAA MGA ACG -3′ | 83–103 |
| YFALL F | 5′- GCT AAT TGA GGT GYA TTG GTC TGC -3′ | 15–38 |
| VFA 17D R | 5′- TTT AAG TGC GGA GYC CGG TT- 3′ | 10448–10667 |
| VFA 17D F | 5′- TAC AAA CCA CGG GTG GAG AA -3′ | 10382–10401 |
| Probe | Sequence (5′→3′) a | Genome Position b |
| YFALL | 5′-FAM-ATC GAG TTG/ ZEN/ CTA GGC AAT AAA CAC-BHQ1 -3′ | 41–64 |
| VFA 17D | 5′-HEX-ACT TGA AAC/ ZEN/ CGG GAT ATA AAC CAC GGC TGG-BHQ1 -3′ | 10416–10445 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Queiroz, A.L.N.; Barros, R.S.; Silva, S.P.; Rodrigues, D.S.G.; Cruz, A.C.R.; dos Santos, F.B.; Vasconcelos, P.F.C.; Tesh, R.B.; Nunes, B.T.D.; Medeiros, D.B.A. The Usefulness of a Duplex RT-qPCR during the Recent Yellow Fever Brazilian Epidemic: Surveillance of Vaccine Adverse Events, Epizootics and Vectors. Pathogens 2021, 10, 693. https://doi.org/10.3390/pathogens10060693
Queiroz ALN, Barros RS, Silva SP, Rodrigues DSG, Cruz ACR, dos Santos FB, Vasconcelos PFC, Tesh RB, Nunes BTD, Medeiros DBA. The Usefulness of a Duplex RT-qPCR during the Recent Yellow Fever Brazilian Epidemic: Surveillance of Vaccine Adverse Events, Epizootics and Vectors. Pathogens. 2021; 10(6):693. https://doi.org/10.3390/pathogens10060693
Chicago/Turabian StyleQueiroz, Alice L. N., Rafael S. Barros, Sandro P. Silva, Daniela S. G. Rodrigues, Ana C. R. Cruz, Flávia B. dos Santos, Pedro F. C. Vasconcelos, Robert B. Tesh, Bruno T. D. Nunes, and Daniele B. A. Medeiros. 2021. "The Usefulness of a Duplex RT-qPCR during the Recent Yellow Fever Brazilian Epidemic: Surveillance of Vaccine Adverse Events, Epizootics and Vectors" Pathogens 10, no. 6: 693. https://doi.org/10.3390/pathogens10060693
APA StyleQueiroz, A. L. N., Barros, R. S., Silva, S. P., Rodrigues, D. S. G., Cruz, A. C. R., dos Santos, F. B., Vasconcelos, P. F. C., Tesh, R. B., Nunes, B. T. D., & Medeiros, D. B. A. (2021). The Usefulness of a Duplex RT-qPCR during the Recent Yellow Fever Brazilian Epidemic: Surveillance of Vaccine Adverse Events, Epizootics and Vectors. Pathogens, 10(6), 693. https://doi.org/10.3390/pathogens10060693