
Identification of Salicylic Acid Mechanism against Leaf Blight Disease in Oryza sativa by SR-FTIR Microspectroscopic and Docking Studies

 , ,
, ,
Abstract
1. Introduction
2. Results
2.1. The Effectiveness of the SA-Ricemate Elicitor on Inducing Resistance against LB
2.2. Accumulation of Endogenous SA Content
2.3. SR-FTIR Microspectroscopy
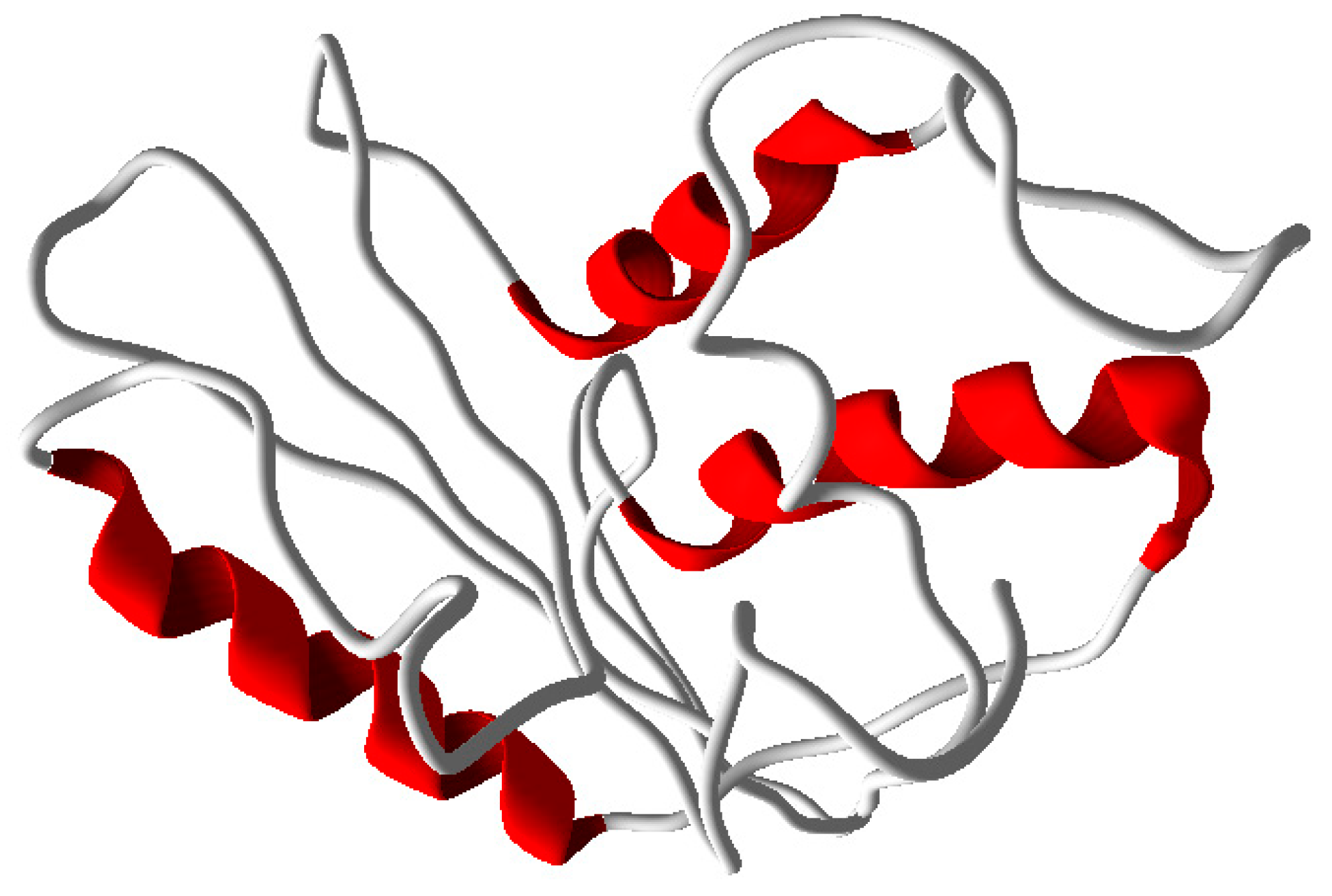
2.4. Homology Modeling of PR1b Domain
2.5. Identification of Template
2.6. Sequence Alignment
2.7. Prediction of the Three-Dimensional Structure
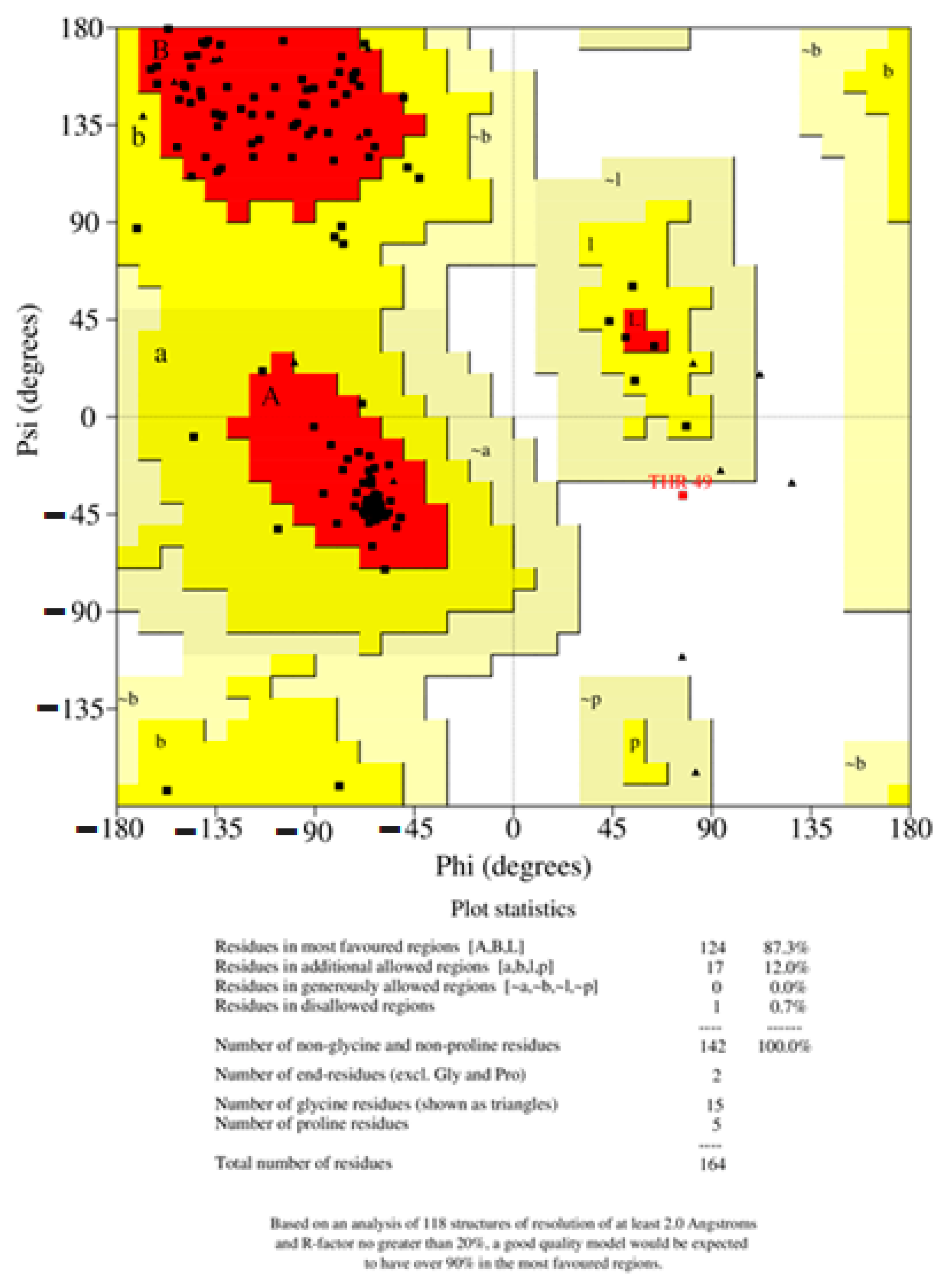
2.8. Structural Validation
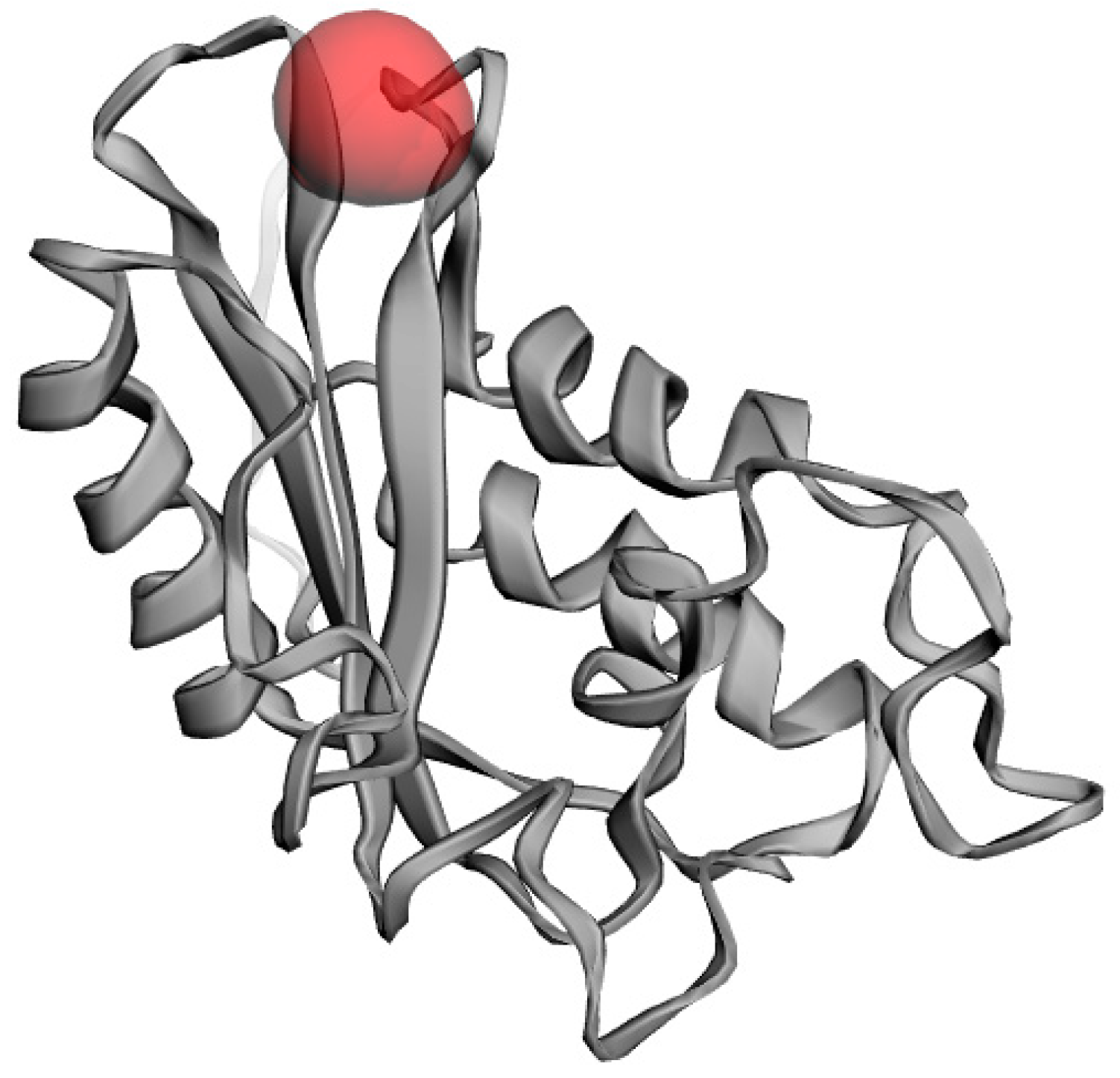
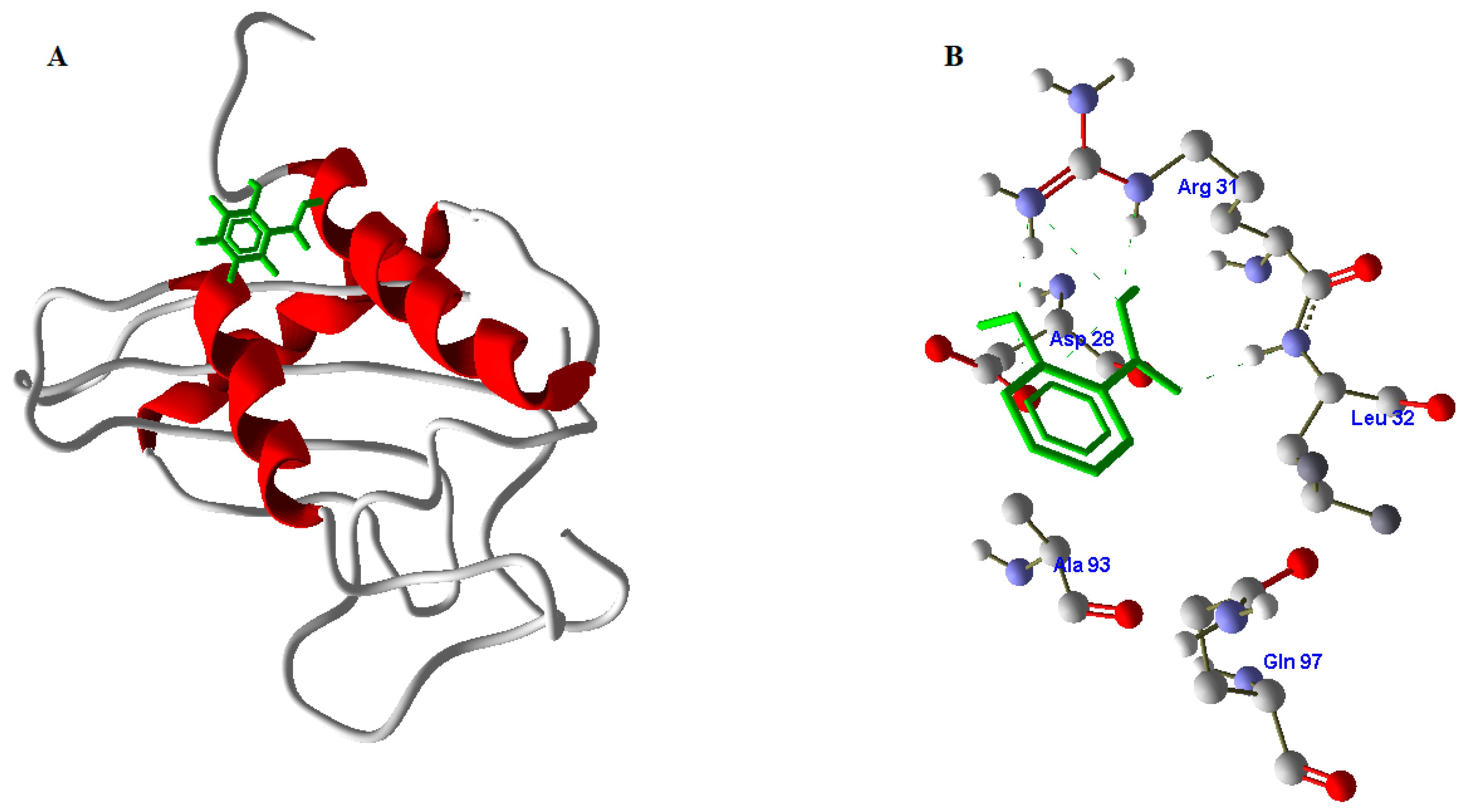
2.9. Active Site Identification
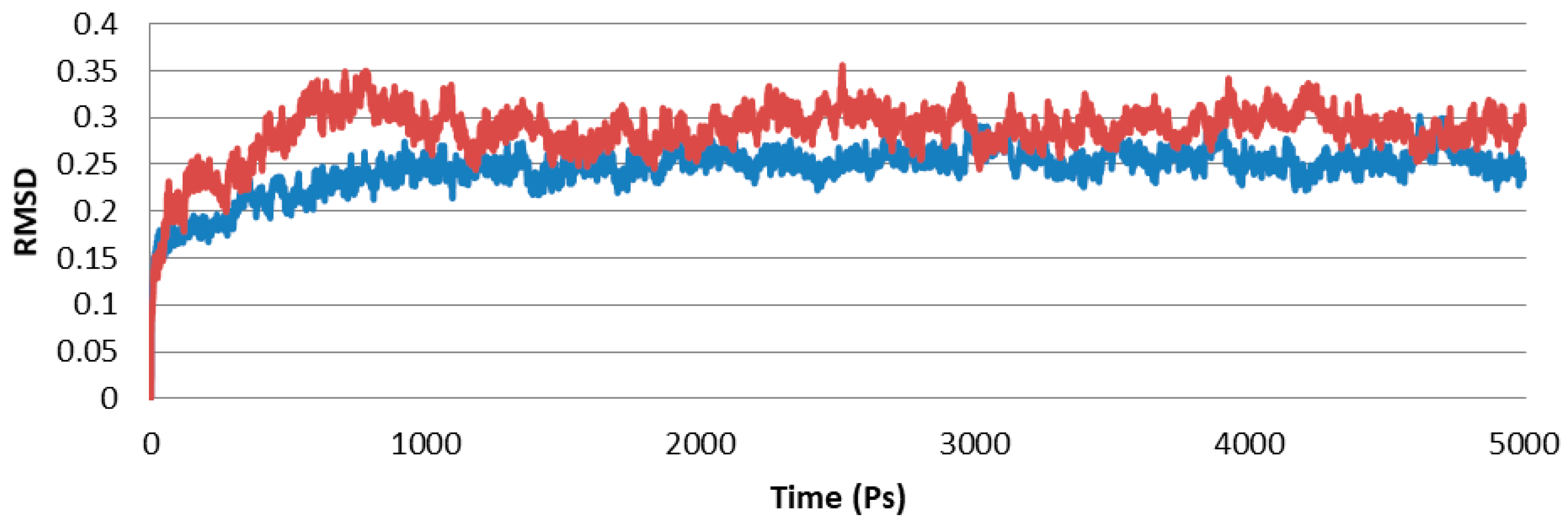
2.10. Docking Studies of SA with PR1b
3. Discussion
4. Materials and Methods
4.1. Rice Cultivar
4.2. Xanthomonas oryzae pv. oryzae Strains and Culture Conditions
4.3. Preparation of a Commercial Abiotic Elicitor Product (SA-Ricemate)
4.4. Efficacy of the SA-Ricemate Elicitor in Inducing Resistance Against LB
4.5. Determination of Endogenous SA
4.6. Biochemical Change Analyses Using SR-FTIR Microspectroscopy
4.7. Data Analysis of SR-FTIR Microspectroscopy
4.8. Domain Identification and Template Search
4.9. D Model Building of PR1b
4.10. Structure Validation of PR1b
4.11. Active site Identification of PR1b
4.12. Docking Studies with SA
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Walters, D.; Newton, A.C.; Lyon, G. Induced Resistance for Plant Defence: A Sustainable Approach to Crop Protection; John Wiley & Sons: Hoboken, NJ, USA, 2008. [Google Scholar]
- Shaheen, R.; Zahid Sharif, M.; Amrao, L.; Zheng, A.; Manzoor, M.; Majeed, D.; Kiran, H.; Jafir, M.; Ali, A. Investigation of bacterial leaf blight of rice through various detection tools and its impact on crop yield in Punjab, Pakistan. Pak. J. Bot. 2019, 51, 307–312. [Google Scholar] [CrossRef]
- Nino-Liu, D.O.; Ronald, P.C.; Bordanove, A.J. Xanthomonas oryzae pathovars: Model pathogens of a model crop. Mol. Plant Pathol. 2006, 7, 303–324. [Google Scholar] [CrossRef]
- Gnanamanickam, S.S.; Priyadarisini, V.B.; Narayanan, N.N.; Vasudevan, P.; Kavitha, S. An overview of bacterial blight disease of rice and strategies for its manament. Curr. Sci. 1999, 77, 1435–1443. [Google Scholar]
- Hilaire, E.; Young, S.A.; Willard, L.H.; McGee, J.D.; Sweat, T.; Chittoor, J.M.; Guikema, J.A.; Leach, J.E. Vascular defense responses in rice: Peroxidase accumulation in xylem parenchyma cells and xylem wall thickening. Mol. Plant Microbe Interact. 2001, 14, 1411–1419. [Google Scholar] [CrossRef]
- War, A.R.; Paulraj, M.G.; War, M.Y.; Ignacimuthu, S. Role of salicylic acid in induction of plant defense system in chickpea (Cicer arietinum L.). Plant Signal. Behav. 2011, 6, 1787–1792. [Google Scholar] [CrossRef] [PubMed]
- Trouvelot, S.; Varnier, A.L.; Allègre, M.; Mercier, L.; Baillieul, F.; Arnould, C.; Gianinazzi-Pearson, V.; Klarzynski, O.; Joubert, J.M.; Pugin, A.; et al. A beta-1,3 glucan sulfate induces resistance in grapevine against Plasmopara viticola through priming of defense responses, including HR-like cell death. Mol. Plant. Microbe Interact. 2008, 21, 232–243. [Google Scholar] [CrossRef] [PubMed]
- Bekkar, A.A.; Zaim, S.; Belabid, L. Induction of systemic resistance in chickpea against Fusarium wilt by Bacillus strains. Arch. Phytopathol. Pflanzenschutz 2018, 51, 70–80. [Google Scholar] [CrossRef]
- Lanna-Filho, R.; Souza, R.M.; Alves, E. Induced resistance in tomato plants promoted by two endophytic bacilli against bacterial speck. Trop. Plant Pathol. 2017, 42, 96–108. [Google Scholar] [CrossRef]
- Morris, S.W.; Vernooij, B.; Titatarn, S.; Starrett, M.; Thomas, S.; Wiltse, C.C.; Frederiksen, R.A.; Bhandhufalck, A.; Hulbert, S.; Uknes, S. Induced resistance responses in maize. Mol. Plant Microbe Interact. 1998, 11, 643–658. [Google Scholar] [CrossRef]
- Liyanage, S.; Dassanayake, R.S.; Bouyanfif, A.; Rajakaruna, E.; Ramalingam, L.; Moustaid-Moussa, N.; Abidi, N. Optimization and validation of cryostat temperature conditions for trans-reflectance mode FTIR microspectroscopic imaging of biological tissues. MethodsX 2017, 4, 118–127. [Google Scholar] [CrossRef]
- Thakur, M.; Sohal, B.S. Role of elicitors in inducing resistance in plants against pathogen infection: A review. ISRN Biochem. 2013, 10, 762412. [Google Scholar] [CrossRef]
- Zaynab, M.; Fatima, M.; Abbas, S.; Sharif, Y.; Umair, M.; Zafar, M.H.; Bahadar, K. Role of secondary metabolites in plant defense against pathogens. Microb. Pathog. 2018, 124, 198–202. [Google Scholar] [CrossRef]
- Shao, Z.; Li, Z.; Fu, Y.; Wen, Y.; Wei, S. Induction of defense responses against Magnaporthe oryzae in rice seedling by a new potential biocontrol agent Streptomyces JD211. J. Basic Microbiol. 2018, 58, 686–697. [Google Scholar] [CrossRef] [PubMed]
- Le Thanh, T.; Thumanu, K.; Wongkaew, S.; Boonkerd, N.; Teaumroong, N.; Phansak, P.; Buensanteai, N. Salicylic acid-induced accumulation of biochemical components associated with resistance against Xanthomonas oryzae pv. oryzae in rice. J. Plant Interact. 2017, 12, 108–120. [Google Scholar] [CrossRef]
- Yu, P. Application of advanced synchrotron radiation-based Fourier transform infrared (SR-FTIR) microspectroscopy to animal nutrition and feed science: A novel approach. Br. J. Nutr. 2004, 92, 869–885. [Google Scholar] [CrossRef] [PubMed]
- Theophilou, G.; Morais, C.L.M.; Halliwell, D.E.; Lima, K.M.G.; Drury, J.; Martin-Hirsch, P.L.; Stringfellow, H.F.; Hapangama, D.K.; Martin, F.L. Synchrotron- and focal plane array-based Fourier-transform infrared spectroscopy differentiates the basalis and functionalis epithelial endometrial regions and identifies putative stem cell regions of human endometrial glands. Anal. Bioanal. Chem. 2018, 410, 4541–4554. [Google Scholar] [CrossRef]
- Thumanu, K.; Sompong, M.; Phansak, P.; Nontapot, K.; Buensanteai, N. Use of infrared microspectroscopy to determine leaf biochemical composition of cassava in response to Bacillus subtilis CaSUT007. J. Plant Interact. 2015, 10, 270–279. [Google Scholar] [CrossRef]
- Sivakumar, S.; Khatiwada, C.P.; Sivasubramanian, J. Studies the alterations of biochemical and mineral contents in bone tissue of mus musculus due to aluminum toxicity and the protective action of desferrioxamine and deferiprone by FTIR, ICP-OES, SEM and XRD techniques. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2014, 126, 59–67. [Google Scholar] [CrossRef]
- Goto, S.; Sasakura-Shimoda, F.; Yamazaki, M.; Hayashi, N.; Suetsugu, M.; Ochiai, H.; Takatsuji, H. Development of disease-resistant rice by pathogen-responsive expression of WRKY45. Plant Biotech. J. 2016, 14, 1127–1138. [Google Scholar] [CrossRef]
- Block, A.; Schmelz, E.; O’Donnell, P.J.; Jones, J.B.; Klee, H.J. Systemic acquired tolerance to virulent bacterial pathogens in tomato. Plant Physiol. 2005, 138, 1481–1490. [Google Scholar] [CrossRef]
- Wani, A.B.; Chadar, H.; Wani, A.H.; Singh, S.; Upadhyay, N. Salicylic acid to decrease plant stress. Environ. Chem. Lett. 2017, 15, 101–123. [Google Scholar] [CrossRef]
- Yang, J.; Wang, Y.; Liu, L.; Liu, L.; Wang, C.; Wang, C.; Li, C. Effects of exogenous salicylic acid and pH on pathogenicity of biotrophy-associated secreted protein 1 (BAS1)-overexpressing strain, Magnaporthe oryzae. Environ. Sci. Pollut. Res. 2019, 26, 13725–13737. [Google Scholar] [CrossRef] [PubMed]
- Adam, A.L.; Nagy, Z.Á.; Kátay, G.; Mergenthaler, E.; Viczián, O. Signals of systemic immunity in plants: Progress and open questions. Int. J. Mol. Sci. 2018, 19, 1146. [Google Scholar] [CrossRef]
- Norouzi, M.; Beheshtizadeh, H.; Sattari, A. Bacterial Blight Resistance in Rice: A Review. Int. J. Sci. Res. Sci. Technol. 2015, 1, 2. [Google Scholar]
- Lopez-Gresa, M.P.; Lisón, P.; Yenush, L.; Conejero, V.; Rodrigo, I.; Bellés, J.M. Salicylic acid is involved in the basal resistance of tomato plants to Citrus Exocortis Viroid and Tomato Spotted Wilt Virus. PLoS ONE 2016, 11, e0166938. [Google Scholar] [CrossRef]
- Metraux, J.P.; Signer, H.; Ryals, J.; Ward, E.; Wyss-Benz, M.; Gaudin, J.; Raschdorf, K.; Schmid, E.; Blum, W.; Inverardi, B. Increase in salicylic acid at the onset of systemic acquired resistance in cucumber. Science 1990, 250, 1004–1006. [Google Scholar] [CrossRef] [PubMed]
- Aranega-Bou, P.; de la O Leyva, M.; Finiti, I.; García-Agustín, P.; González-Bosch, C. Priming of plant resistance by natural compounds. Hexanoic acid as a model. Front. Plant Sci. 2014, 5, 488. [Google Scholar] [CrossRef] [PubMed]
- Genzel, F.; Franken, P.; Witzel, K.; Grosch, R. Systemic induction of salicylic acid-related plant defences in potato in response to Rhizoctonia solani AG3PT. Plant Pathol. 2018, 67, 337–348. [Google Scholar] [CrossRef]
- Gharbi, E.; Martínez, J.; Ben Ahmed, H.; Dailly, H.; Quinet, M.; Lutts, S. The salicylic acid analog 2,6-dichloroisonicotinic acid has specific impact on the response of the halophyte plant species Solanum chilense to salinity. Plant Growth Regul. 2017, 82, 517–525. [Google Scholar] [CrossRef]
- Sahebani, N.; Hadavi, N. Induction of H2O2 and related enzymes in tomato roots infected with root knot nematode (M. javanica) by several chemical and microbial elicitors. Biocontrol Sci. Technol. 2009, 19, 301–313. [Google Scholar] [CrossRef]
- Gao, M.; Yin, X.; Yang, W.; Lam, S.M.; Tong, X.; Liu, J.; Wang, X.; Li, Q.; Shui, G.; He, Z. GDSL lipases modulate immunity through lipid homeostasis in rice. PLoS Pathog. 2017, 13, e1006724. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Xiao, S. Lipids in salicylic acid-mediated defense in plants: Focusing on the roles of phosphatidic acid and phosphatidylinositol 4-phosphate. Front. Plant Sci. 2015, 6, 387. [Google Scholar] [CrossRef] [PubMed]
- Thumanu, K.; Wongchalee, D.; Sompong, M.; Phansak, P.; Le Thanh, T.; Namanusart, W.; Vechklang, K.; Kaewnum, S.; Buensanteai, N. Synchrotron-based FTIR microspectroscopy of chili resistance induced by Bacillus subtilis strain D604 against anthracnose disease. J. Plant Interact. 2017, 12, 255–263. [Google Scholar] [CrossRef]
- Wu, C.T.; Leubner-Metzger, G.; Meins, F., Jr.; Bradford, K.J. Class I beta-1,3-glucanase and chitinase are expressed in the micropylar endosperm of tomato seeds prior to radicle emergence. Plant Physiol. 2001, 126, 1299–1313. [Google Scholar] [CrossRef] [PubMed]
- Leubner-Metzger, G.; Meins, F., Jr. Sense transformation reveals a novel role for class I β-1,3-glucanase in tobacco seed germination. Plant J. 2000, 23, 215–221. [Google Scholar] [CrossRef]
- Anita, B.; Samiyappan, R. Induction of systemic resistance in rice by Pseudomonas fluorescens against rice root knot nematode Meloidogyne graminicola. J. Biopestic. 2012, 5, 53–59. [Google Scholar]
- Lahlali, R.; Song, T.; Chu, M.; Yu, F.; Kumar, S.; Karunakaran, C.; Peng, G. Evaluating changes in cell-wall components associated with clubroot resistance using Fourier Transform Infrared Spectroscopy and RT-PCR. Int. J. Mol. Sci. 2017, 18, 2058. [Google Scholar] [CrossRef]
- Macoy, D.M.; Kim, W.-Y.; Lee, S.Y.; Kim, M.G. Biotic stress related functions of hydroxycinnamic acid amide in plants. J. Plant Biol. 2015, 58, 156–163. [Google Scholar] [CrossRef]
- Buchanan, E.G.; James, W.H.J., III; Choi, S.H.; Guo, L.; Gellman, S.H.; Müller, C.W.; Zwier, T.S. Single-conformation infrared spectra of model peptides in the amide I and amide II regions: Experiment-based determination of local mode frequencies and inter-mode coupling. Int. J. Chem. Phys. 2012, 137, 094301. [Google Scholar] [CrossRef]
- Bolton, M.D. Primary metabolism and plant defense—Fuel for the fire. Mol. Plant Microbe. Interact. 2009, 22, 487–497. [Google Scholar] [CrossRef]
- Trouvelot, S.; Héloir, M.-C.; Poinssot, B.; Gauthier, A.; Paris, F.; Guillier, C.; Combier, M.; Trdá, L.; Daire, X.; Adrian, M. Carbohydrates in plant immunity and plant protection: Roles and potential application as foliar sprays. Front. Plant Sci. 2014, 5, 592. [Google Scholar] [CrossRef] [PubMed]
- Tauzin, A.S.; Giardina, T. Sucrose and invertases, a part of the plant defense response to the biotic stresses. Front. Plant Sci. 2014, 5, 293. [Google Scholar] [CrossRef]
- Zhao, D.; You, Y.; Fan, H.; Zhu, X.; Wang, Y.; Duan, Y.; Xuan, Y.; Chen, L. The role of sugar transporter genes during early infection by root-knot nematodes. Int. J. Mol. Sci. 2018, 19, 302. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.K.; Srivastava, M.; Osmanoglu, Ö.; Dandekar, T. Genome-wide inference of the Camponotus floridanus protein-protein interaction network using homologous mapping and interacting domain profile pairs. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Durak, T.; Depciuch, J. Effect of plant sample preparation and measuring methods on ATR-FTIR spectra results. Environ. Exp. Bot. 2020, 169, 103915. [Google Scholar] [CrossRef]
- Daddam, J.R.; Dowlathabad, M.R.; Panthangi, S.; Jasti, P. Molecular docking and P-glycoprotein inhibitory activity of flavonoids. Interdiscip Sci. 2014, 6, 167–175. [Google Scholar] [CrossRef]
- Papathoti, N.K.; Saengchan, C.; Daddam, J.R.; Thongprom, N.; Tonpho, K.; Thanh, T.L.; Buensanteai, N. Plant systemic acquired resistance compound salicylic acid as a potent inhibitor against SCF (SKP1-CUL1-F-box protein) mediated complex in Fusarium oxysporum by homology modeling and molecular dynamics simulations. J. Biomol. Struct. Dyn. 2020, 1–8. [Google Scholar] [CrossRef]
- Daddam, J.R.; Sreenivasulu, B.; Peddanna, K.; Umamahesh, K. Designing, docking and molecular dynamics simulation studies of novel cloperastine analogues as anti-allergic agents: Homology modeling and active site prediction for the human histamine H1 receptor. RSC. Adv. 2020, 10, 4745–4754. [Google Scholar] [CrossRef]
- Daddam, J.R.; Sreenivasulu, B.; Umamahesh, K.; Peddanna, K.; Rao, D.M. In Silico studies on anti-stress compounds of ethanolic root extract of Hemidesmus indicus L. Curr. Pharm. Biotechnol. 2020, 21, 502–515. [Google Scholar] [CrossRef]
- Gupta, S.K.; Osmanoglu, Ö.; Srivastava, M.; Bencúrová, E.; Dandekar, T. Pathogen and Host-Pathogen Protein Interactions Provide a Key to Identify Novel Drug Targets. In Reference Module in Biomedical Sciences; Elsevier: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Bencurova, E.; Gupta, S.K.; Sarukhanyan, E.; Dandekar, T. Identification of antifungal targets based on computer modeling. J. Fungi 2018, 4, 81. [Google Scholar] [CrossRef]
- Kurjogi, M.; Satapute, P.; Jogaiah, S.; Abdelrahman, M.; Daddam, J.R.; Ramu, V.; Tran, L.-S.P. Computational modeling of the Staphylococcal enterotoxins and their interaction with natural antitoxin compounds. Int. J. Mol. Sci. 2018, 19, 133. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.N.; Swapna, T.H.; Khan, M.Y.; Daddam, J.R.; Hameeda, B. Molecular dynamics and protein interaction studies of lipopeptide (Iturin A) on α- amylase of Spodoptera litura. J. Theor. Biol. 2017, 415, 41–47. [Google Scholar] [CrossRef]
- Hussein, N.; Daddam, J.; Prasad, E.; Naidu, N. Evaluation of novel curcumin derivatives against methicillin resistant Staphylococcus aureus (MRSA). Int. J. Appl. Biol. Pharm. 2017, 8, 111–120. [Google Scholar] [CrossRef]
- Suresh, B.B.; Mangamoori, L.N.; Jayasimharayalu, D. In Silico docking studies of Elytraria acaulis Gas chromatography-Mass Spectroscopy derived compound against breast cancer target proteins. World J. Pharm. Pharm. Sci. 2017, 6, 1053–1062. [Google Scholar]
- Papathoti, N.; Lingampally, N.; Parameshwar, J.; Khan, M.; Rayalu, J.; Hameeda, B. In Silico and In Vitro studies of fungicidal nature of lipopeptide (Iturin A) from Bacillus amyloliquefaciens RHNK 22 and its plant growth promoting traits. Indian Phytopathol. 2016, 69, 569–574. [Google Scholar]
- Urama, D.; Tarigopula, S.; Pasha, K.; Daddam, J. Homology modelling and structure of Neplanocin A derivative. Onl. J. Bioinform. 2016, 17, 41–52. [Google Scholar]
- Daddam, J.R.; Rao, M.; Rao, D.S. Phytochemical screening and anti microbial studies of Hemidesmus indicus. Int. J. Appl. Biol. Pharm. 2015, 6, 10–13. [Google Scholar]
- Beda, P.; Vedula, S.; Daddam, J. Anti-tubercularl activity and molecular docking of dihydro-pyrimidinone derivatives. Onl. J. Bioinform. 2015, 16, 202–225. [Google Scholar]
- Masroor, H.; Parvateesam, M.; Daddam, J.; Naidu, N. In Silico docking of meliantrol, β-sitosterol, curcumin, vanillic and syringic acids to penicillin binding protein 2a on methicillin resistant Staphylococcus aureus. Onl. J. Bioinform. 2015, 16, 88–97. [Google Scholar]
- Shaik, A.H.; Shaik, S.R.; Daddam, J.R.; Ali, D.; Manoharadas, S.; Arafah, M.W.; Kodidhela, L.D. Maslinic acid and gallic acid protective efficacy on lipids, lipoproteins and lipid metabolizing enzymes against isoproterenol administered cardiotoxicity: An In Vivo and In Silico molecular docking evidences. J. King Saud Univ. Sci. 2021, 33, 101230. [Google Scholar] [CrossRef]
- Tsai, B.C.-K.; Kuo, W.-W.; Day, C.H.; Hsieh, D.J.-Y.; Kuo, C.-H.; Daddam, J.; Chen, R.-J.; Padma, V.V.; Wang, G.; Huang, C.-Y. The soybean bioactive peptide VHVV alleviates hypertension-induced renal damage in hypertensive rats via the SIRT1-PGC1α/Nrf2 pathway. J. Funct. Foods 2020, 75, 104255. [Google Scholar] [CrossRef]
- Smita, S.; Singh, K.P.; Akhoon, B.A.; Gupta, S.K. Bioinformatics Tools for Interpretation of Data Used in Molecular Identification. In Analyzing Microbes. Springer Protocols Handbooks; Arora, D., Das, S., Sukumar, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Krishnan, N.; Gandhi, K.; Peeran, M.F.; Muthurajan, R.; Kuppusamy, P.; Thiruvengadam, R. Management of bacterial leaf blight disease in rice with endophytic bacteria. World Appl. Sci. J. 2014, 28, 2229–2241. [Google Scholar]
- Buensanteai, N.; Yuen, G.; Prathuangwong, S. The biocontrol bacterium Bacillus amyloliquefaciens KPS46 produces auxin, surfactin and extracellular proteins for enhanced growth of soybean plant. Thai J. Agric. Sci. 2008, 41, 101–116. [Google Scholar]
- Mizobuchi, R.; Sato, H.; Fukuoka, S.; Tsushima, S.; Imbe, T.; Yano, M. Identification of qRBS1, a QTL involved in resistance to bacterial seedling rot in rice. TAG. Theor. Appl. Genet. 2013, 126, 2417–2425. [Google Scholar] [CrossRef] [PubMed]
- Udayashankar, A.C.; Chandra Nayaka, S.; Reddy, M.S.; Srinivas, C. Plant growth-promoting rhizobacteria mediate induced systemic resistance in rice against bacterial leaf blight caused by Xanthomonas oryzae pv. oryzae. Biol. Control 2011, 59, 114–122. [Google Scholar] [CrossRef]
- Xu, J.; Audenaert, K.; Hofte, M.; De Vleesschauwer, D. Abscisic acid promotes susceptibility to the rice leaf blight pathogen Xanthomonas oryzae pv. oryzae by suppressing salicylic acid-mediated defenses. PLoS ONE 2013, 8, e67413. [Google Scholar] [CrossRef]
- Ke, Y.; Hui, S.; Yuan, M. Xanthomonas oryzae pv. oryzae inoculation and growth rate on rice by leaf clipping method. Bioprotocol 2017, 7, e2568. [Google Scholar] [CrossRef]
- Ferreira, D.; Falahati, F.; Linden, C.; Buckley, R.F.; Ellis, K.A.; Savage, G.; Villemagne, V.L.; Rowe, C.C.; Ames, D.; Simmons, A.; et al. A ‘Disease Severity Index’ to identify individuals with subjective memory decline who will progress to mild cognitive impairment or dementia. Sci. Rep. 2017, 7, 44368. [Google Scholar] [CrossRef]
- Rozhon, W.; Petutschnig, E.; Wrzaczek, M.; Jonak, C. Quantification of free and total salicylic acid in plants by solid-phase extraction and isocratic high-performance anion-exchange chromatography. Anal. Bioanal. Chem. 2005, 382, 1620–1627. [Google Scholar] [CrossRef]
- Clayton, A.W.; Thiers, R.E. Direct spectrophotometric determination of salicylic acid, acetylsalicylic acid, salicylamide, caffeine, and phenacetin in tablets or powders. J. Pharm. Sci. 1966, 55, 404–407. [Google Scholar] [CrossRef]
- Wang, M.; Lu, X.; Yin, X.; Tong, Y.; Peng, W.; Wu, L.; Li, H.; Yang, Y.; Gu, J.; Xiao, T.; et al. Synchrotron radiation-based Fourier-transform infrared spectromicroscopy for characterization of the protein/peptide distribution in single microspheres. Acta Pharm. Sin. B 2015, 5, 270–276. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Thepbandit, W.; Buensanteai, N.; Thumanu, K.; Siriwong, S.; Toan Le, T.; Athinuwat, D. Salicylic acid elicitor inhibiting xanthomonas oryzae growth, motility, biofilm, polysaccharides production, and biochemical components during pathogenesis on rice. Chiang Mai J. Sci. 2021, 48, 341–353. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment 1 | Disease Severity (%) 2 | Control Efficacy % (%) | ||||
|---|---|---|---|---|---|---|
| 7 DPI | 14 DPI | 21 DPI | 7 DPI | 14 DPI | 21 DPI | |
| SA-Ricemate 50 ppm | 26.04 ± 2.75 c | 35.42 ± 4.54 c | 39.58 ± 2.75 c | 13.80 ± 7.75 a | 12.88 ± 4.96 a | 26.77 ± 10.83 a |
| SA-Ricemate 100 ppm | 17.71 ± 2.75 b | 18.75 ± 1.04 b | 21.88 ± 1.04 b | 41.39 ± 4.81 b | 53.88 ± 1.18 b | 59.53 ± 3.91 b |
| SA-Ricemate 150 ppm | 15.63 ± 3.12 b | 16.67 ± 1.04 b | 22.92 ± 1.52 b | 48.28 ± 8.51 b | 59.00 ± 3.12 b | 57.60 ± 5.19 b |
| SA-Ricemate 200 ppm | 16.67 ± 2.77 b | 19.79 ± 2.11 b | 21.88 ± 1.00 b | 44.83 ± 7.39 b | 51.31 ± 3.25 b | 59.53 ± 2.99 b |
| SA-Ricemate 250 ppm | 13.54 ± 2.71 b | 17.71 ± 2.08 b | 20.83 ± 2.18 b | 55.18 ± 7.73 b | 56.44 ± 3.49 b | 61.46 ± 3.99 b |
| SA-Ricemate 300 ppm | 12.50 ± 2.85 b | 16.67 ± 1.41 b | 17.71 ± 1.44 ab | 58.63 ± 6.58 b | 59.00 ± 5.86 b | 67.24 ± 3.95 cb |
| Commercial elicitor | 12.50 ± 2.14 b | 17.50 ± 1.61 b | 22.50 ± 1.12 b | 57.13 ± 5.98 b | 58.00 ± 3.60 b | 58.38 ± 3.12 b |
| Copper hydroxide 77% WP | 8.33 ± 1.04 a | 10.42 ± 1.24 a | 12.50 ± 1.20 a | 72.42 ± 4.13 c | 74.38 ± 2.88 c | 76.88 ± 1.71 c |
| Control (water) | 30.21 ± 4.10 c | 40.63 ± 2.98 d | 54.17 ± 3.75 d | 0 | 0 | 0 |
| Treatment 1 | Endogenous Salicylic Acid (µg g−1 of Fresh Weight) 2 | Increase of SA Activity (%) | |
|---|---|---|---|
| Pre Inoculation | Post Inoculation 24 h | ||
| SA-Ricemate 50 ppm | 12.12 ± 0.17 a | 17.60 ± 0.03 b | 30.78 ± 1.87 b |
| SA-Ricemate 100 ppm | 12.20 ± 0.18 a | 18.46 ± 0.10 b | 51.27 ± 3.10 b |
| SA-Ricemate 150 ppm | 12.39 ± 0.14 a | 18.34 ± 0.08 b | 48.00 ± 4.42 b |
| SA-Ricemate 200 ppm | 11.78 ± 0.26 a | 18.14 ± 0.21 b | 54.05 ± 6.24 b |
| SA-Ricemate 250 ppm | 12.28 ± 0.22 a | 18.28 ± 0.26 b | 48.84 ± 4.42 b |
| SA-Ricemate 300 ppm | 12.19 ± 0.21 a | 18.46 ± 0.25 b | 51.44 ± 4.64 b |
| Commercial elicitor | 11.99 ± 0.38 a | 16.61 ± 0.37 b | 38.33 ± 4.17 b |
| Copper hydroxide 77% WP | 11.69 ± 0.08 a | 13.03 ± 0.32 a | 5.19 ± 1.57 a |
| Control (water) | 13.21 ± 0.10 a | 13.89 ± 0.07 a | 11.40 ± 2.57 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thepbandit, W.; Papathoti, N.K.; Daddam, J.R.; Thumanu, K.; Siriwong, S.; Thanh, T.L.; Buensanteai, N. Identification of Salicylic Acid Mechanism against Leaf Blight Disease in Oryza sativa by SR-FTIR Microspectroscopic and Docking Studies. Pathogens 2021, 10, 652. https://doi.org/10.3390/pathogens10060652
Thepbandit W, Papathoti NK, Daddam JR, Thumanu K, Siriwong S, Thanh TL, Buensanteai N. Identification of Salicylic Acid Mechanism against Leaf Blight Disease in Oryza sativa by SR-FTIR Microspectroscopic and Docking Studies. Pathogens. 2021; 10(6):652. https://doi.org/10.3390/pathogens10060652
Chicago/Turabian StyleThepbandit, Wannaporn, Narendra Kumar Papathoti, Jayasimha Rayalu Daddam, Kanjana Thumanu, Supatcharee Siriwong, Toan Le Thanh, and Natthiya Buensanteai. 2021. "Identification of Salicylic Acid Mechanism against Leaf Blight Disease in Oryza sativa by SR-FTIR Microspectroscopic and Docking Studies" Pathogens 10, no. 6: 652. https://doi.org/10.3390/pathogens10060652
APA StyleThepbandit, W., Papathoti, N. K., Daddam, J. R., Thumanu, K., Siriwong, S., Thanh, T. L., & Buensanteai, N. (2021). Identification of Salicylic Acid Mechanism against Leaf Blight Disease in Oryza sativa by SR-FTIR Microspectroscopic and Docking Studies. Pathogens, 10(6), 652. https://doi.org/10.3390/pathogens10060652