
Detection of Tick-Borne Pathogens in Red Deer (Cervus elaphus), United Kingdom


Abstract
1. Introduction
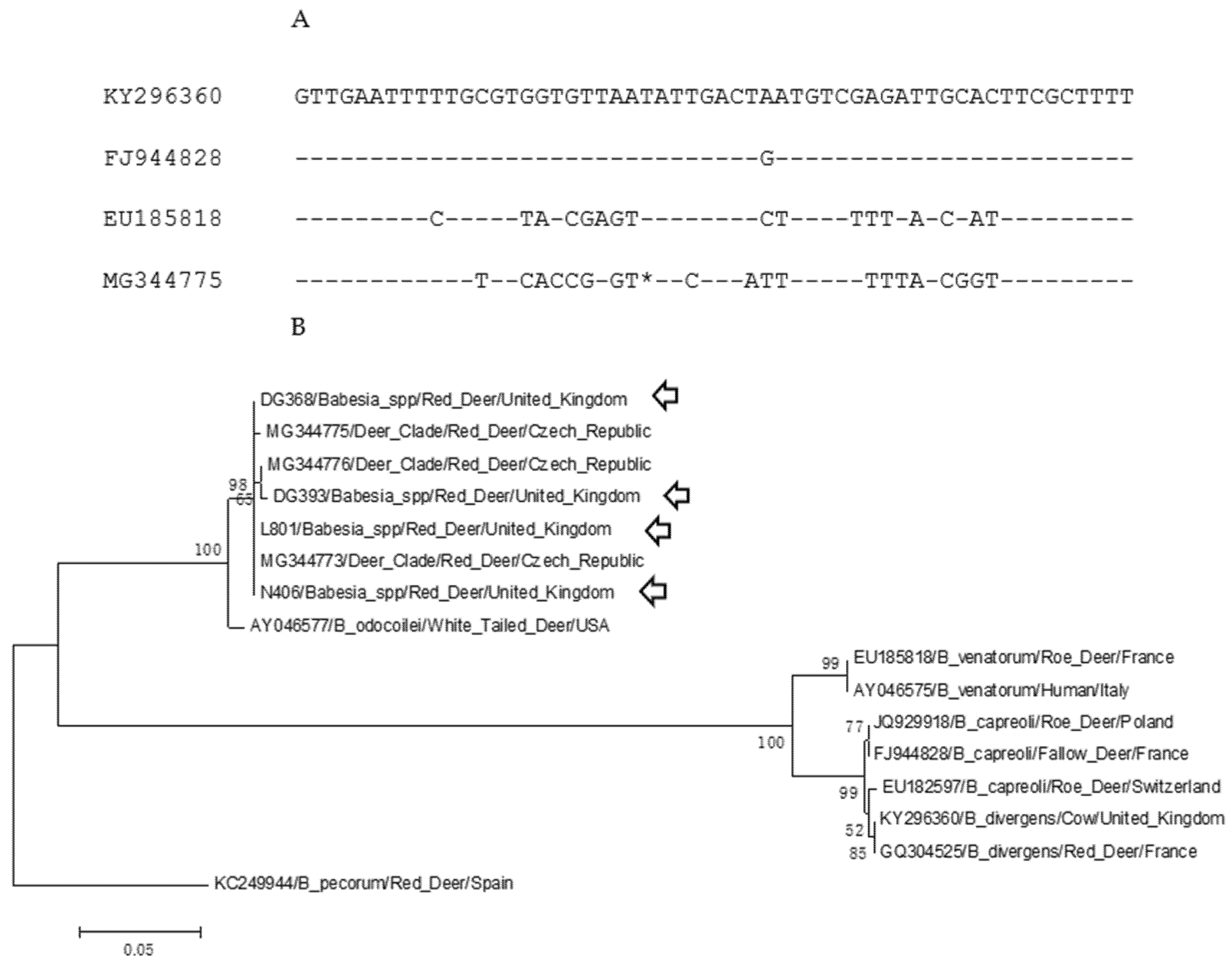
2. Results
3. Discussion
4. Materials and Methods
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Simpson, V.R. Wild animals as reservoirs of infectious diseases in the UK. Vet. J. 2002, 163, 128–146. [Google Scholar] [CrossRef]
- Böhm, M.; White, P.C.L.; Chambers, J.; Smith, L.; Hutchings, M.R. Wild deer as a source of infection for livestock and humans in the UK. Vet. J. 2007, 174, 260–276. [Google Scholar] [CrossRef] [PubMed]
- Ward, A.I. Expanding ranges of wild and feral deer in Great Britain. Mamm. Rev. 2005, 35, 165–173. [Google Scholar] [CrossRef]
- Schnittger, L.; Rodriguez, A.E.; Florin-Christensen, M.; Morrision, D.A. Babesia: A world emerging. Infect. Genet. Evol. 2012, 12, 1788–1809. [Google Scholar] [CrossRef] [PubMed]
- Phipps, L.P.; Hernández-Triana, L.M.; Goharriz, H.; Welchman, D.; Johnson, N. Detection of Theileria luwenshuni in sheep from Great Britain. Parasit. Vectors 2016, 9, 203. [Google Scholar] [CrossRef] [PubMed]
- M’Fadyean, J.; Stockman, S. A new species of piroplasm found in the blood of British Cattle. J. Comp. Pathol. 1911, 24, 340–354. [Google Scholar] [CrossRef]
- Zintl, A.; Mulcahy, G.; Skerrett, H.E.; Taylor, S.M.; Gray, J.S. Babesia divergens, a bovine blood parasite of veterinary and zoonotic importance. Clin. Microbiol. Rev. 2003, 16, 622–636. [Google Scholar] [CrossRef] [PubMed]
- Lewis, D.; Purnell, R.E.; Shaw, S.R. The isolation and characterization of human and bovine strains of Babesia divergens from Drumnadroit, Scotland. Parasitology 1980, 81, 145–155. [Google Scholar] [CrossRef]
- Lv, J.; Fernández de Marco, M.; Phipps, L.P.; Macrelli, M.; Otter, A.; Inman, B.; Mitchell, S.; Johnson, N. Molecular detection of Babesia divergens from an outbreak of Babesiosis in Holstein cows, England. J. Vet. Med. Res. 1917, 4, 1078. [Google Scholar]
- Brocklesby, D.W.; Zwart, S.; Perié, N.M. Serological evidence for the identification of Babesia major in Britain. Res. Vet. Sci. 1971, 12, 285–287. [Google Scholar] [CrossRef]
- Lewis, D.; Holman, M.R.; Purnell, R.E.; Young, E.R.; Herbert, I.C.; Bevan, W.J. Investigations on Babesia motasi isolated from Wales. Res. Vet. Sci. 1981, 31, 239–243. [Google Scholar] [CrossRef]
- Woldehiwet, Z. The natural history of Anaplasma phagocytophilum. Vet. Parasitol. 2010, 167, 108–122. [Google Scholar] [CrossRef]
- Azagi, T.; Hoogrstra, D.; Kremer, K.; Hovious, J.W.R.; Sprong, H. Evaluation of disease causality of rare Ixodes ricinus-borne infections in Europe. Pathogens 2020, 9, 150. [Google Scholar] [CrossRef]
- Alberdi, M.P.; Walker, A.R.; Urquhart, K.A. Field evidence that roe deer (Capreolus capreolus) are a natural host for Ehrlichia phagocytophila. Epidemiol. Infect. 2000, 124, 315–323. [Google Scholar] [CrossRef]
- Overzier, E.; Pfister, K.; Herb, I.; Mahling, M.; Böck, G., Jr.; Silaghi, C. Detection of tick-borne pathogens in roe deer (Capreolus capreolus), in questing ticks (Ixodes ricinus), and in tick infesting roe deer in southern Germany. Ticks Tick-Borne Dis. 2013, 4, 320–328. [Google Scholar] [CrossRef]
- Razanske, I.; Rosef, O.; Radzijevskaja, J.; Bratchikov, M.; Griciuviene, L.; Paulauskas, A. Prevalence and co-infection with tick-borne Anaplasma phagocytophilum and Babesia spp. in red deer (Cervus elephus) and roe deer (Capreolus capreolus) in southern Norway. Int. J. Parasitol. Parasites Wildl. 2019, 8, 127–134. [Google Scholar] [CrossRef]
- Ebani, V.V.; Rocchigiani, G.; Bertelloni, F.; Nardoni, S.; Leoni, A.; Nicoloso, S.; Mancianti, F. Molecular survey on the presence of zoonotic arthropod-borne pathogens in wild red deer (Cervus elephas). Comp. Immunol. Microbiol. Infect. Dis. 2016, 47, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Cézanne, R.; Mrowietz, N.; Eigner, B.; Duscher, G.G.; Glawischnig, W.; Feuhrer, H.-P. Molecular analysis of Anaplasma phagocytophilum and Babesia divergens in red deer (Cervus elephas) in Western Austria. Mol. Cell. Probes 2017, 31, 55–58. [Google Scholar] [CrossRef] [PubMed]
- Kauffman, M.; Rehbein, S.; Hamel, D.; Lutz, W.; Heddergott, M.; Pfister, K.; Silhagi, C. Anaplasma phagocytophilum and Babesia spp. in roe deer (Capreolus capreolus), fallow deer (Dama dama) and mouflon (Ovis musimon) in Germany. Mol. Cell. Probes 2017, 31, 46–54. [Google Scholar] [CrossRef]
- Latif, B.M.A.; Adam, K.M.G. Antibody to Babesia in Scottish red deer (Cervus elaphus). Nature 1973, 241, 476–477. [Google Scholar] [CrossRef] [PubMed]
- Adam, K.M.G.; Blewett, D.A.; Brocklesby, D.W.; Sharman, G.A.W. The isolation and characterization of a Babesia from red deer (Cervus elaphus). Parasitology 1976, 73, 1–11. [Google Scholar] [CrossRef]
- Enigk, K.; Freidhoff, K. Babesia capreoli n.sp. beim Reh (Capreolus capreolus). Z. Tropenmed. Parasitol. 1962, 13, 8–20. [Google Scholar]
- Duh, D.; Petrovec, M.; Bidovec, A.; Avsic-Zupanc, T. Cervids as Babesiae hosts, Slovenia. Emerg. Infect. Dis. 2003, 11, 1121–1123. [Google Scholar] [CrossRef] [PubMed]
- Herwaldt, B.L.; Caccio, S.; Gherlinzoni, F.; Aspock, H.; Slemenda, S.B.; Piccaluga, P.; Martinelli, G.; Edelhofer, R.; Hollenstein, U.; Poletti, G.; et al. Molecular characterization of non-Babesia divergens organism causing zoonotic babesiosis in Europe. Emerg. Infect. Dis. 2003, 9, 942–948. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, S.; Jouglin, M.; L’Hostis, M.; Chauvin, A. Babesia sp. EU1 from roe deer and transmission within Ixodes ricinus. Emerg. Infect. Dis. 2007, 13, 1208–1210. [Google Scholar] [CrossRef]
- Smith, F.D.; Ellse, L.; Wall, R. Prevalence of Babesia and Anaplasma in ticks infesting dogs in Great Britain. Vet. Parasitol. 2013, 198, 18–23. [Google Scholar] [CrossRef]
- Gray, A.; Capewell, P.; Loney, C.; Katzer, F.; Shiels, B.R.; Weir, W. Sheep as host species for zoonotic Babesia venatorum, United Kingdom. Emerg. Infect. Dis. 2019, 25, 2257–2260. [Google Scholar] [CrossRef]
- Emerson, H.R.; Wright, W.T. The isolation of a Babesia in white-tailed deer. Wildl. Dis. 1968, 4, 142. [Google Scholar] [CrossRef][Green Version]
- Perry, B.D.; Nichols, D.K.; Cullom, E.S. Babesia odocoilei Emerson and Wright, 1970 in white-tailed deer, Odocoileus virginianus (Zimmerman), in Virginia. J. Wildl. Dis. 1985, 21, 149–152. [Google Scholar] [CrossRef] [PubMed]
- Inokuma, H.; Yoshizaki, Y.; Shimada, Y.; Sakata, Y.; Okuda, M.; Onishi, T. Epidemiological survey of Babesia species in Japan performed with specimens from ticks collected from dogs and detection of new Babesia DNA closely related to Babesia odocoilei and Babesia divergens DNA. J. Clin. Microbiol. 2003, 41, 3494–3498. [Google Scholar] [CrossRef]
- Hilpertshauser, H.; Deplazes, P.; Schnyder, M.; Gern, L.; Mathis, A. Babesia spp. identified by PCR in ticks collected from domestic and wild domestic and wild ruminants in Switzerland. Appl. Environ. Microbiol. 2006, 72, 6503–6507. [Google Scholar] [CrossRef]
- García-Sanmartin, J.; Aurtenetxe, O.; Barral, M.; Marco, I.; Lavin, S.; Lavin, S.; García-Pérez, A.L.; Hurtado, A. Molecular detection and characterization of piroplasms infecting cervids and chamois in Northern Spain. Parasitology 2007, 134, 391–398. [Google Scholar] [CrossRef]
- Jouglin, M.; Fernández-de-Mera, I.G.; De al Cotte, N.; Ruis-Fons, F.; Gortázar, C.; Moreau, E.; Bastian, S.; de la Fuente, J.; Malandrin, L. Isolation and characterization of Babesia pecorum sp. nov. from farmed red deer (Cervus elephas). Vet. Res. 2014, 45, 78. [Google Scholar]
- Pereira, A.; Parreira, R.; Nunes, M.; Casadinho, A.; Viera, M.L.; Campino, L.; Maia, C. Molecular detection of tick-borne bacteria and protozoa in cervids and wild boar from Portugal. Parasit. Vectors 2016, 9, 251. [Google Scholar] [CrossRef]
- Holding, M.; Dowall, S.D.; Medlock, J.M.; Carter, D.P.; Pullan, S.T.; Lewis, J.; Vipond, R.; Rocchi, M.S.; Baylis, M.; Hewson, R. Tick-borne encephalitis virus, United Kingdom. Emerg. Infect. Dis. 2020, 26, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Zintl, A.; McGrath, G.; O’Grady, L.; Fanning, J.; Downing, K.; Roche, D.; Casey, M.; Gray, J.S. Changing incidence of bovine Babesiosis in Ireland. Ir. Vet. J. 2014, 67, 19. [Google Scholar] [CrossRef]
- Hradilová, K.; Rybářová, M.; Široký, P.; Votýpka, J.; Zintl, A.; Burgess, H.; Steinbauer, V.; Žákočík, V.; Modrý, D. Diversity of Babesia spp. in cervid ungulates based on the 18S rDNA and cytochrome c oxidase subunit 1 phylogenies. Infect. Genet. Evol. 2020, 77, 104060. [Google Scholar] [CrossRef]
- Duscher, G.G.; Battisti, E.; Hodžić, A.; Wäber, K.; Steinbach, P.; Stubbé, M.; Heddergott, M. First detection and molecular identification of Anaplasma phagocytophilum in an introduced population of Reeve’s muntjac (Muntiacus reevesi) in United Kingdom. Mol. Cell. Probes 2020, 52, 101528. [Google Scholar] [CrossRef]
- Langton, C.; Gray, J.S.; Water, P.F.; Holman, P.J. Naturally-acquired Babesiosis in a reindeer (Rangifer tarandus tarandus) herd in Great Britain. Parasitol. Res. 2003, 89, 194–198. [Google Scholar] [CrossRef] [PubMed]
- Malandrin, L.; Jouglin, M.; Sun, Y.; Brisseau, N.; Chauvin, A. Redescription of Babesia capreoli (Enigk and Freidhoff, 1962) from roe deer (Capreolus capreolus): Isolation, cultivation, host specificity, molecular characterization and differentiation from Babesia divergens. Int. J. Parasitol. 2010, 40, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Zintl, A.; Finnerty, E.J.; Murphy, T.M.; de Waal, T.; Gray, J.S. Babesias of red deer (Cervus elaphus) in Ireland. Vet. Res. 2011, 42, 7. [Google Scholar] [CrossRef] [PubMed]
- Folly, A.J.; Waller, E.S.L.; McCracken, F.; McElhinney, L.M.; Roberts, H.; Johnson, N. Equine seroprevalence of West Nile virus antibodies in the UK in 2019. Parasit. Vectors 2020, 13, 596. [Google Scholar] [CrossRef] [PubMed]
- Adam, K.M.; Beasley, S.J.; Blewett, D.A. The occurrence of antibody to Babesia and to the virus louping-ill in deer in Scotland. Res. Vet. Sci. 1977, 23, 133–138. [Google Scholar] [CrossRef]
- Courtney, J.W.; Kostelnik, L.M.; Zeidner, N.S.; Massung, R.E. Multiplex real-time PCR for detection of Anaplasma phagocytophilum and Borrelia burgdorferi. J. Clin. Microbiol. 2004, 42, 3164–3168. [Google Scholar] [CrossRef]
- Fernández de Marco, M.D.M.; Hernández-Triana, L.M.; Phipps, L.P.; Hansford, K.; Mitchell, E.S.; Cull, B.; Swainsbury, C.S.; Fooks, A.R.; Medlock, J.M.; Johnson, N. Emergence of Babesia canis in southern England. Parasit. Vectors 2017, 10, 241. [Google Scholar] [CrossRef]
- Johnson, N.; Phipps, L.P.; McFadzean, H.; Barlow, A.M. An outbreak of bovine Babesiosis in February, 2019, triggered by unusual high winter temperatures in southern England and co-infection with Babesia divergens and Anaplasma phagocytophilum. Parasit. Vectors 2020, 13, 305. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likliehood, evolutionary, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- Entrican, J.H.; Williams, H.; Cook, I.A.; Lancaster, W.M.; Clark, J.C.; Joyner, L.P.; Lewis, D. Babesiosis in man: A case from Scotland. Br. Med. J. 1979, 2, 474. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bläckberg, J.; Lazarevic, C.L.; Hunfeld, K.-P.; Persson, K.E.M. Low-virulent Babesia venatorum infection masquerading as hemophagocytic syndrome. Ann. Hematol. 2018, 97, 731–733. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Location | Year/Samples | Total | Anaplasma phagocytophilum PCR (%) [Code Numbers] | Pan-Piroplasm spp. PCR (%) [Code Numbers] |
|---|---|---|---|---|
| Norfolk (N) | 2020 0♂, 11♀ | 11 | 3/11 (27.3%) [N405, N406, N408] | 2/11 (18.2%) [N406, N408] |
| Dumfries and Galloway (DG) | 2020 50♂, 18♀ | 68 | 1/68 (1.5%) [DG422] | 2/68 (2.9%) [DG368, DG393] |
| Lancashire (L) | 2020 13♂, 13♀ | 26 | 1/26 (3.8%) [L811] | 1/26 (3.8%) [L801] |
| Totals | 105 | 5/105 (4.8%) | 5/105 (4.8%) |
| Location | West Nile Virus 1 (ID Screen® West Nile Competition Multi-Species) (%) | Tick-Borne Encephalitis Virus 2 (Immunozym FSME IgG All Species) (%) |
|---|---|---|
| Norfolk | 0/9 (0) | 0/9 (0) |
| Dumfries and Galloway | 0/14 (0) | 4/15 (26.7) |
| Lancashire | 0/16 (0) | 1/17 (5.9) |
| Total | 0/39 (0) | 5/41 (12.1) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johnson, N.; Golding, M.; Phipps, L.P. Detection of Tick-Borne Pathogens in Red Deer (Cervus elaphus), United Kingdom. Pathogens 2021, 10, 640. https://doi.org/10.3390/pathogens10060640
Johnson N, Golding M, Phipps LP. Detection of Tick-Borne Pathogens in Red Deer (Cervus elaphus), United Kingdom. Pathogens. 2021; 10(6):640. https://doi.org/10.3390/pathogens10060640
Chicago/Turabian StyleJohnson, Nicholas, Megan Golding, and Laurence Paul Phipps. 2021. "Detection of Tick-Borne Pathogens in Red Deer (Cervus elaphus), United Kingdom" Pathogens 10, no. 6: 640. https://doi.org/10.3390/pathogens10060640
APA StyleJohnson, N., Golding, M., & Phipps, L. P. (2021). Detection of Tick-Borne Pathogens in Red Deer (Cervus elaphus), United Kingdom. Pathogens, 10(6), 640. https://doi.org/10.3390/pathogens10060640