
Amplicon-Based Next Generation Sequencing for Rapid Identification of Rickettsia and Ectoparasite Species from Entomological Surveillance in Thailand


Abstract
1. Introduction
2. Results
2.1. A High Percentage of Rickettsia DNA in Fleas Collected from Domestic Dogs
2.2. Rickettsia Species in Rodent and Ectoparasite Pools
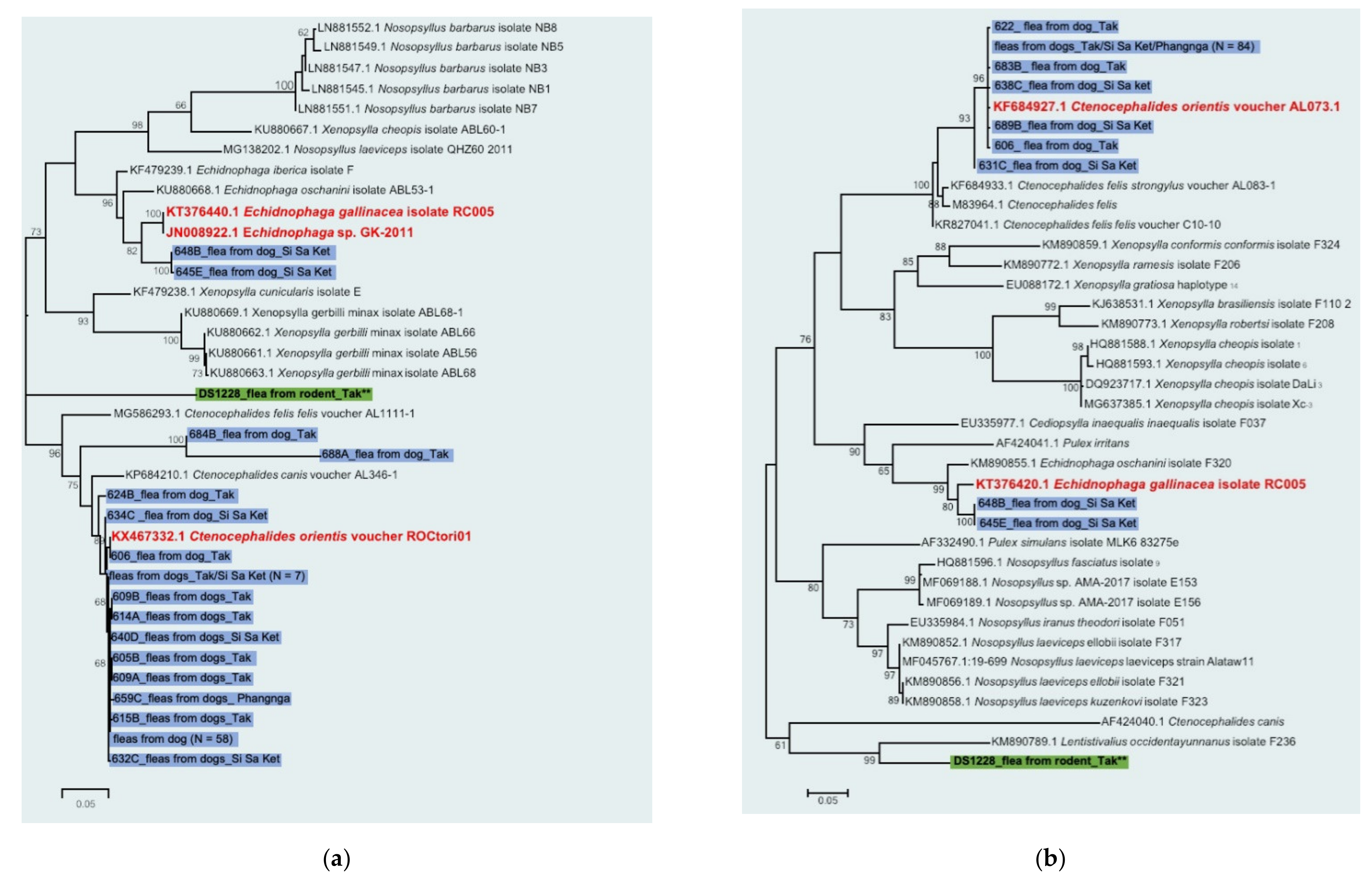
2.3. Ectoparasite Species Identification in Rickettsia-Positive Pools
3. Discussion
4. Materials and Methods
4.1. The Field Surveillance for Rickettsia in Rodents and Ectoparasites
4.2. Genomic DNA Extraction from Rodents and Ectoparasite Samples
4.3. Real-Time PCR Detection of Rickettsia
4.4. Amplicon-Based NGS Characterization of Rickettsia and Ectoparasite Species
4.5. Sequence Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Disclaimer
References
- Parte, A.C.; Sarda Carbasse, J.; Meier-Kolthoff, J.P.; Reimer, L.C.; Goker, M. List of Prokaryotic names with Standing in Nomenclature (LPSN) moves to the DSMZ. Int. J. Syst. Evol. Microbiol. 2020, 70, 5607–5612. [Google Scholar] [CrossRef]
- Blanton, L.S. The Rickettsioses: A Practical Update. Infect. Dis. Clin. N. Am. 2019, 33, 213–229. [Google Scholar] [CrossRef] [PubMed]
- Graves, S.R.; Stenos, J. Tick-borne infectious diseases in Australia. Med. J. Aust. 2017, 206, 320–324. [Google Scholar] [CrossRef] [PubMed]
- Zaharia, M.; Popescu, C.P.; Florescu, S.A.; Ceausu, E.; Raoult, D.; Parola, P.; Socolovschi, C. Rickettsia massiliae infection and SENLAT syndrome in Romania. Ticks Tick Borne Dis. 2016, 7, 759–762. [Google Scholar] [CrossRef]
- Parola, P.; Paddock, C.D.; Socolovschi, C.; Labruna, M.B.; Mediannikov, O.; Kernif, T.; Abdad, M.Y.; Stenos, J.; Bitam, I.; Fournier, P.E.; et al. Update on tick-borne rickettsioses around the world: A geographic approach. Clin. Microbiol. Rev. 2013, 26, 657–702. [Google Scholar] [CrossRef]
- Tarragona, E.L.; Soares, J.F.; Costa, F.B.; Labruna, M.B.; Nava, S. Vectorial competence of Amblyomma tonelliae to transmit Rickettsia rickettsii. Med. Vet. Entomol. 2016, 30, 410–415. [Google Scholar] [CrossRef]
- Drexler, N.A.; Yaglom, H.; Casal, M.; Fierro, M.; Kriner, P.; Murphy, B.; Kjemtrup, A.; Paddock, C.D. Fatal Rocky Mountain Spotted Fever along the United States-Mexico Border, 2013–2016. Emerg. Infect. Dis. 2017, 23, 1621–1626. [Google Scholar] [CrossRef]
- Straily, A.; Drexler, N.; Cruz-Loustaunau, D.; Paddock, C.D.; Alvarez-Hernandez, G. Notes from the Field: Community-Based Prevention of Rocky Mountain Spotted Fever—Sonora, Mexico, 2016. MMWR Morb. Mortal. Wkly. Rep. 2016, 65, 1302–1303. [Google Scholar] [CrossRef]
- Levin, M.L.; Killmaster, L.F.; Zemtsova, G.E. Domestic dogs (Canis familiaris) as reservoir hosts for Rickettsia conorii. Vector Borne Zoonotic Dis. 2012, 12, 28–33. [Google Scholar] [CrossRef]
- Jensenius, M.; Fournier, P.E.; Kelly, P.; Myrvang, B.; Raoult, D. African tick bite fever. Lancet Infect. Dis. 2003, 3, 557–564. [Google Scholar] [CrossRef]
- Robinson, M.T.; Satjanadumrong, J.; Hughes, T.; Stenos, J.; Blacksell, S.D. Diagnosis of spotted fever group Rickettsia infections: The Asian perspective. Epidemiol. Infect. 2019, 147, e286. [Google Scholar] [CrossRef]
- Sirisanthana, T.; Pinyopornpanit, V.; Sirisanthana, V.; Strickman, D.; Kelly, D.J.; Dasch, G.A. First cases of spotted fever group rickettsiosis in Thailand. Am. J. Trop. Med. Hyg. 1994, 50, 682–686. [Google Scholar] [CrossRef]
- Jiang, J.; Sangkasuwan, V.; Lerdthusnee, K.; Sukwit, S.; Chuenchitra, T.; Rozmajzl, P.J.; Eamsila, C.; Jones, J.W.; Richards, A.L. Human Infection with Rickettsia honei, Thailand. Emerg. Infect. Dis. 2005, 11, 1473–1475. [Google Scholar] [CrossRef] [PubMed]
- Gaywee, J.; Sunyakumthorn, P.; Rodkvamtook, W.; Ruang-areerate, T.; Mason, C.J.; Sirisopana, N. Human infection with Rickettsia sp. related to R. japonica, Thailand. Emerg. Infect. Dis. 2007, 13, 657–659. [Google Scholar] [CrossRef]
- Robertson, R.G.; Wisseman, C.L., Jr. Tick-borne rickettsiae of the spotted fever group in West Pakistan. II. Serological classification of isolates from West Pakistan and Thailand: Evidence for two new species. Am. J. Epidemiol. 1973, 97, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Takada, N.; Fujita, H.; Kawabata, H.; Ando, S.; Sakata, A.; Takano, A.; Chaithong, U. Spotted fever group Rickettsia sp. closely related to Rickettsia japonica, Thailand. Emerg. Infect. Dis. 2009, 15, 610–611. [Google Scholar] [CrossRef] [PubMed]
- Luvira, V.; Silachamroon, U.; Piyaphanee, W.; Lawpoolsri, S.; Chierakul, W.; Leaungwutiwong, P.; Thawornkuno, C.; Wattanagoon, Y. Etiologies of Acute Undifferentiated Febrile Illness in Bangkok, Thailand. Am. J. Trop. Med. Hyg. 2019, 100, 622–629. [Google Scholar] [CrossRef]
- Kuo, C.C.; Wardrop, N.; Chang, C.T.; Wang, H.C.; Atkinson, P.M. Significance of major international seaports in the distribution of murine typhus in Taiwan. PLoS Negl. Trop. Dis. 2017, 11, e0005430. [Google Scholar] [CrossRef]
- Blanton, L.S.; Idowu, B.M.; Tatsch, T.N.; Henderson, J.M.; Bouyer, D.H.; Walker, D.H. Opossums and Cat Fleas: New Insights in the Ecology of Murine Typhus in Galveston, Texas. Am. J. Trop. Med. Hyg. 2016, 95, 457–461. [Google Scholar] [CrossRef]
- Maina, A.N.; Fogarty, C.; Krueger, L.; Macaluso, K.R.; Odhiambo, A.; Nguyen, K.; Farris, C.M.; Luce-Fedrow, A.; Bennett, S.; Jiang, J.; et al. Rickettsial Infections among Ctenocephalides felis and Host Animals during a Flea-Borne Rickettsioses Outbreak in Orange County, California. PLoS ONE 2016, 11, e0160604. [Google Scholar] [CrossRef] [PubMed]
- Foongladda, S.; Inthawong, D.; Kositanont, U.; Gaywee, J. Rickettsia, Ehrlichia, Anaplasma, and Bartonella in ticks and fleas from dogs and cats in Bangkok. Vector Borne Zoonotic Dis. 2011, 11, 1335–1341. [Google Scholar] [CrossRef] [PubMed]
- Mongkol, N.; Suputtamongkol, Y.; Taweethavonsawat, P.; Foongladda, S. Molecular Evidence of Rickettsia in Human and Dog Blood in Bangkok. Vector Borne Zoonotic Dis. 2018, 18, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Phomjareet, S.; Chaveerach, P.; Suksawat, F.; Jiang, J.; Richards, A.L. Spotted Fever Group Rickettsia Infection of Cats and Cat Fleas in Northeast Thailand. Vector Borne Zoonotic Dis. 2020, 20, 566–571. [Google Scholar] [CrossRef]
- Papa, A.; Tsioka, K.; Kontana, A.; Papadopoulos, C.; Giadinis, N. Bacterial pathogens and endosymbionts in ticks. Ticks Tick Borne Dis. 2017, 8, 31–35. [Google Scholar] [CrossRef]
- Pesquera, C.; Portillo, A.; Palomar, A.M.; Oteo, J.A. Investigation of tick-borne bacteria (Rickettsia spp., Anaplasma spp., Ehrlichia spp. and Borrelia spp.) in ticks collected from Andean tapirs, cattle and vegetation from a protected area in Ecuador. Parasit. Vectors 2015, 8, 46. [Google Scholar] [CrossRef]
- Kurilshikov, A.; Livanova, N.N.; Fomenko, N.V.; Tupikin, A.E.; Rar, V.A.; Kabilov, M.R.; Livanov, S.G.; Tikunova, N.V. Comparative Metagenomic Profiling of Symbiotic Bacterial Communities Associated with Ixodes persulcatus, Ixodes pavlovskyi and Dermacentor reticulatus Ticks. PLoS ONE 2015, 10, e0131413. [Google Scholar] [CrossRef]
- Portillo, A.; Palomar, A.M.; de Toro, M.; Santibanez, S.; Santibanez, P.; Oteo, J.A. Exploring the bacteriome in anthropophilic ticks: To investigate the vectors for diagnosis. PLoS ONE 2019, 14, e0213384. [Google Scholar] [CrossRef]
- Takhampunya, R.; Korkusol, A.; Pongpichit, C.; Yodin, K.; Rungrojn, A.; Chanarat, N.; Promsathaporn, S.; Monkanna, T.; Thaloengsok, S.; Tippayachai, B.; et al. Metagenomic Approach to Characterizing Disease Epidemiology in a Disease-Endemic Environment in Northern Thailand. Front. Microbiol. 2019, 10, 319. [Google Scholar] [CrossRef]
- Galan, M.; Razzauti, M.; Bard, E.; Bernard, M.; Brouat, C.; Charbonnel, N.; Dehne-Garcia, A.; Loiseau, A.; Tatard, C.; Tamisier, L.; et al. 16S rRNA Amplicon Sequencing for Epidemiological Surveys of Bacteria in Wildlife. mSystems 2016, 1. [Google Scholar] [CrossRef]
- Perez-Losada, M.; Arenas, M.; Castro-Nallar, E. Microbial sequence typing in the genomic era. Infect. Genet. Evol. 2018, 63, 346–359. [Google Scholar] [CrossRef]
- Kalkofen, U.P.; Greenberg, J. Echidnophaga gallinacea infestation in dogs. J. Am. Vet. Med. Assoc. 1974, 165, 447–448. [Google Scholar]
- Hii, S.F.; Lawrence, A.L.; Cuttell, L.; Tynas, R.; Abd Rani, P.A.; Slapeta, J.; Traub, R.J. Evidence for a specific host-endosymbiont relationship between ‘Rickettsia sp. genotype RF2125’ and Ctenocephalides felis orientis infesting dogs in India. Parasit. Vectors 2015, 8, 169. [Google Scholar] [CrossRef]
- Maina, A.N.; Jiang, J.; Luce-Fedrow, A.; St John, H.K.; Farris, C.M.; Richards, A.L. Worldwide Presence and Features of Flea-Borne Rickettsia asembonensis. Front. Vet. Sci. 2018, 5, 334. [Google Scholar] [CrossRef]
- Parola, P.; Cornet, J.P.; Sanogo, Y.O.; Miller, R.S.; Thien, H.V.; Gonzalez, J.P.; Raoult, D.; Telford, I.S.; Wongsrichanalai, C. Detection of Ehrlichia spp., Anaplasma spp., Rickettsia spp., and other eubacteria in ticks from the Thai-Myanmar border and Vietnam. J. Clin. Microbiol. 2003, 41, 1600–1608. [Google Scholar] [CrossRef]
- Rodkvamtook, W.; Pholpetch, S.; Yokanit, W.; Thongphun, A.; Somsri, M.; Somsri, K.; Boonsom, V.; Thimhaipol, C.; Siriwatthanakul, K.; Amradee, B.; et al. Rickettsia asembonensis causing febrile disease in southern Thailand, 2016–2018. In Proceedings of the 29th Meeting of the American Society for Rickettsiology, Milwaukee, WI, USA, 16–19 June 2018. [Google Scholar]
- Palacios-Salvatierra, R.; Caceres-Rey, O.; Vasquez-Dominguez, A.; Mosquera-Visaloth, P.; Anaya-Ramirez, E. Rickettsial species in human cases with non-specific acute febrile syndrome in Peru. Rev. Peru Med. Exp. Salud. Publica 2018, 35, 630–635. [Google Scholar] [CrossRef]
- Kho, K.L.; Koh, F.X.; Singh, H.K.; Zan, H.A.; Kukreja, A.; Ponnampalavanar, S.; Tay, S.T. Spotted Fever Group Rickettsioses and Murine Typhus in a Malaysian Teaching Hospital. Am. J. Trop. Med. Hyg. 2016, 95, 765–768. [Google Scholar] [CrossRef]
- Kolo, A.O.; Sibeko-Matjila, K.P.; Maina, A.N.; Richards, A.L.; Knobel, D.L.; Matjila, P.T. Molecular Detection of Zoonotic Rickettsiae and Anaplasma spp. in Domestic Dogs and Their Ectoparasites in Bushbuckridge, South Africa. Vector Borne Zoonotic Dis. 2016, 16, 245–252. [Google Scholar] [CrossRef]
- Tay, S.T.; Koh, F.X.; Kho, K.L.; Sitam, F.T. Rickettsial infections in monkeys, Malaysia. Emerg. Infect. Dis. 2015, 21, 545–547. [Google Scholar] [CrossRef]
- Liu, H.; Li, Q.; Zhang, X.; Li, Z.; Wang, Z.; Song, M.; Wei, F.; Wang, S.; Liu, Q. Characterization of rickettsiae in ticks in northeastern China. Parasit. Vectors 2016, 9, 498. [Google Scholar] [CrossRef]
- Igolkina, Y.; Rar, V.; Vysochina, N.; Ivanov, L.; Tikunov, A.; Pukhovskaya, N.; Epikhina, T.; Golovljova, I.; Tikunova, N. Genetic variability of Rickettsia spp. in Dermacentor and Haemaphysalis ticks from the Russian Far East. Ticks Tick Borne Dis. 2018, 9, 1594–1603. [Google Scholar] [CrossRef]
- Muul, I. Lekagul, Boonsong, Jeffrey, A. McNeely. Mammals of thailand. Association for the Conservation of Wildlife*, Bangkok, Thailand. J. Mammal. 1979, 60, 241–242. [Google Scholar] [CrossRef]
- Fleas, B.M.R.C.O.; Hopkins, G.H.E.; Siphonaptera, B.M.D.O.E.; Collection, B.M.R.; Rothschild, M.; Mardon, D.K.; Smit, F.G.A.M. An Illustrated Catalogue of the Rothschild Collection of Fleas (Siphonaptera) in the British Museum (Natural History): With Keys and Short Descriptions for the Identification of Families, Genera, Species and Subspecies of the Order. Hystrichopsyllidae (Ctenophthalminae, Dinopsyllinae, Doratopsyllinae and Listropsyllinae); Trustees of the British Museum (Natural History): London, UK, 1966. [Google Scholar]
- Tanskull, P.; Inlao, I. Keys to the adult ticks of Haemaphysalis Koch, 1844, in Thailand with notes on changes in taxonomy (Acari: Ixodoidea: Ixodidae). J. Med. Entomol. 1989, 26, 573–601. [Google Scholar] [CrossRef]
- Durden, L.A.M.; Guy, G. The sucking lice (Insecta, Anoplura) of the world: A taxonomic checklist with records of mammalian hosts and geographical distributions. Bull. AMNH 1994, 218, 1–90. [Google Scholar]
- Whiting, M.F. Mecoptera is paraphyletic: Multiple genes and phylogeny of Mecoptera and Siphonaptera. Zool. Scr. 2002, 31, 93–104. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Sample Type/Provinces | Rickettsia-PCR Positive Pools (% Positive) | |||||
|---|---|---|---|---|---|---|
| Phangnga 8°41′08.5″ N 98°21′07.8″ E | Ranong 10°21′54.7″ N 98°49′01.8″ E | Si Sa Ket 14°34′13.9″ N 104°20′25.9″ E | Tak 16°23′41.4″ N 98°44′03.7″ E | Udon Thani 17°36′44.3″ N 102°20′38.3″ E | Total | |
| Rodent | 0/516 (0%) | 0/137 (0%) | 0/220 (0%) | 1/452* (0.2%) | 1/59* (1.7%) | 2/1384 (0.1%) |
| Rodent Fleas | 0/3 (0%) | NA. | 0/15 (0%) | 1/22 (4.5%) | 0/12 (0%) | 1/52 (1.9%) |
| Rodent Tick | 0/8 (0%) | NA. | 0/12 (0%) | 0/17 (0%) | NA. | 0/37 (0%) |
| Rodent Lice | 0/2 (0%) | NA. | 0/1 (0%) | 0/3 (0%) | NA. | 0/6 (0%) |
| DA Flea1 | 16/21 (76.2%) | NA. | 31/57 (54.4%) | 50/95 (52.6%) | NA. | 97/173 (56.1) |
| DA Tick2 | 2/88 (2%) | NA. | 1/69 (1.4%) | NA. | NA. | 3/157 (1.9%) |
| DA Lice3 | 0/5 (0%) | NA. | 0/6 (0%) | 0/5 (0%) | NA. | 0/16 (0%) |
| Total | 18/643 (2.8%) | 0/137 (0%) | 32/380 (8.4%) | 52/594 (8.8%) | 1/71 (1.4%) | 103/1825 (5.6%) |
| Rickettsia Species Identified | Sample Type | Total Positive Samples (% of Overall Positive Samples) | |||
|---|---|---|---|---|---|
| Rodent | Rodent Flea | DA Flea | DA Tick | ||
| “Ca. R. senegalensis” | 0 | 0 | 16 | 1 | 17 (16.8%) |
| R. asembonensis | 0 | 1 | 79 | 0 | 80 (79.2%) |
| R. helongjiangensis | 0 | 0 | 0 | 2 | 2 (2.0%) |
| R. typhi | 2 | 0 | 0 | 0 | 2 (2.0%) |
| Total | 2 | 1 | 95 | 3 | 101 (100%) |
| Host | Ectoparasite Species | Rickettsia Species | Total | ||
|---|---|---|---|---|---|
| “Ca. R. senegalensis” | R. asembonensis | R. heilongjiangensis | |||
| Rodent | Flea- Unknown species | 0 | 1 | 0 | 1 |
| Dog | Flea- Ctenocephalides f. orientis | 15 | 74 | 0 | 89 |
| Flea- Echidnophaga gallinacea | 1 | 1 | 0 | 2 | |
| Tick- Unknown * | 1 | 0 | 0 | 1 | |
| Tick- Haemaphysalis hystricis | 0 | 0 | 2 | 2 | |
| Total | All species | 17 | 76 | 2 | 95 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaorattanakawee, S.; Korkusol, A.; Tippayachai, B.; Promsathaporn, S.; Poole-Smith, B.K.; Takhampunya, R. Amplicon-Based Next Generation Sequencing for Rapid Identification of Rickettsia and Ectoparasite Species from Entomological Surveillance in Thailand. Pathogens 2021, 10, 215. https://doi.org/10.3390/pathogens10020215
Chaorattanakawee S, Korkusol A, Tippayachai B, Promsathaporn S, Poole-Smith BK, Takhampunya R. Amplicon-Based Next Generation Sequencing for Rapid Identification of Rickettsia and Ectoparasite Species from Entomological Surveillance in Thailand. Pathogens. 2021; 10(2):215. https://doi.org/10.3390/pathogens10020215
Chicago/Turabian StyleChaorattanakawee, Suwanna, Achareeya Korkusol, Bousaraporn Tippayachai, Sommai Promsathaporn, Betty K. Poole-Smith, and Ratree Takhampunya. 2021. "Amplicon-Based Next Generation Sequencing for Rapid Identification of Rickettsia and Ectoparasite Species from Entomological Surveillance in Thailand" Pathogens 10, no. 2: 215. https://doi.org/10.3390/pathogens10020215
APA StyleChaorattanakawee, S., Korkusol, A., Tippayachai, B., Promsathaporn, S., Poole-Smith, B. K., & Takhampunya, R. (2021). Amplicon-Based Next Generation Sequencing for Rapid Identification of Rickettsia and Ectoparasite Species from Entomological Surveillance in Thailand. Pathogens, 10(2), 215. https://doi.org/10.3390/pathogens10020215