Experimental Study of the Potential Role of Salmonella enterica subsp. diarizonae in the Diarrhoeic Syndrome of Lambs

 , ,
, ,  ,
,  ,
,  , and
, and 
Abstract
1. Introduction
2. Results
2.1. Clinical Findings
2.2. Haematological Findings
2.3. Bacteriological Findings
2.3.1. Faecal Swab Samples from Lambs
2.3.2. Buccal Swab Samples from Lambs
2.3.3. Faecal Swab Samples from Ewes
2.3.4. Milk Samples from Ewes
2.3.5. Tissue Samples from Lambs
2.3.6. Identification of Isolates Recovered and Serological Typing
2.4. Molecular Findings
2.5. Pathological Findings
2.5.1. Gross-Pathological Findings
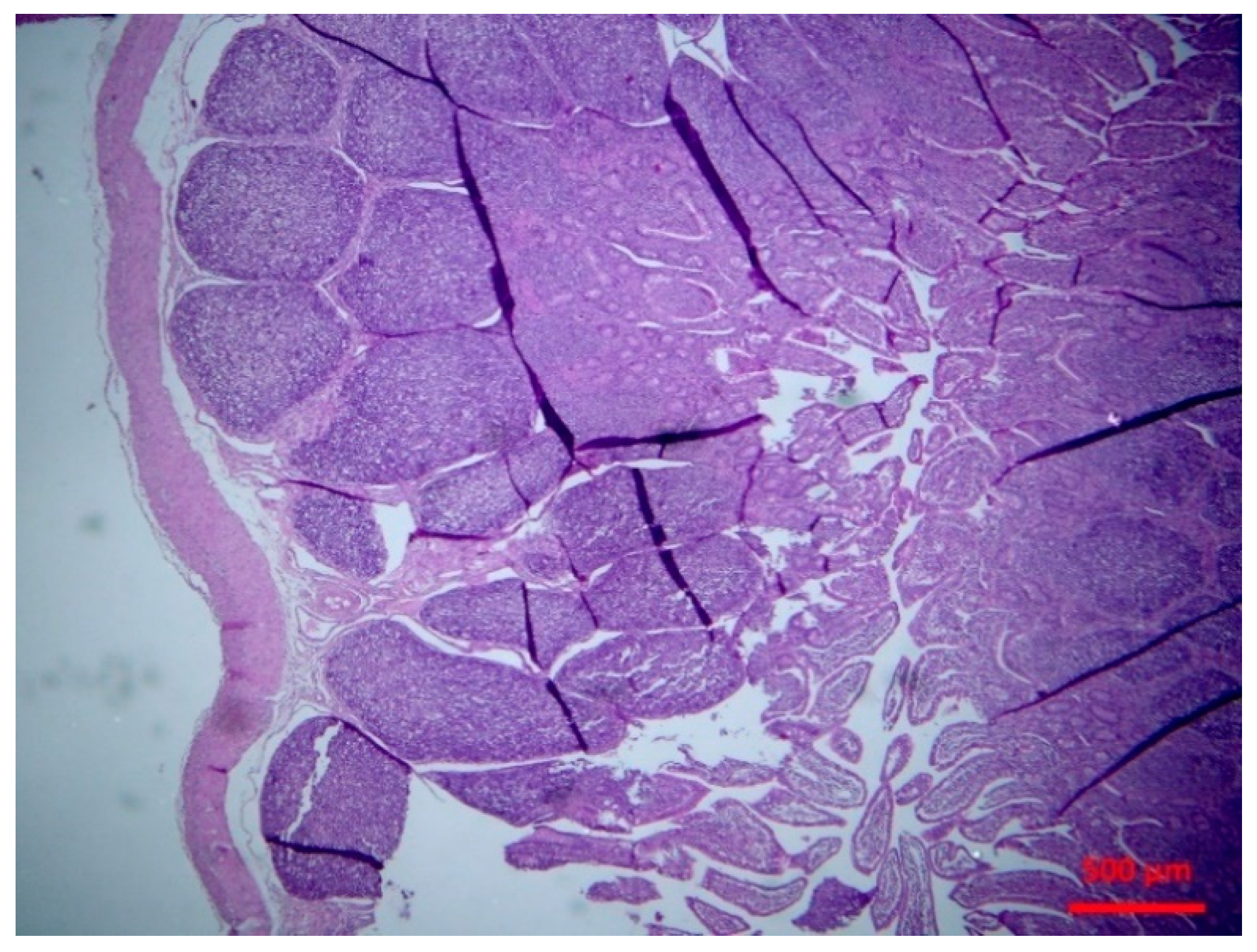
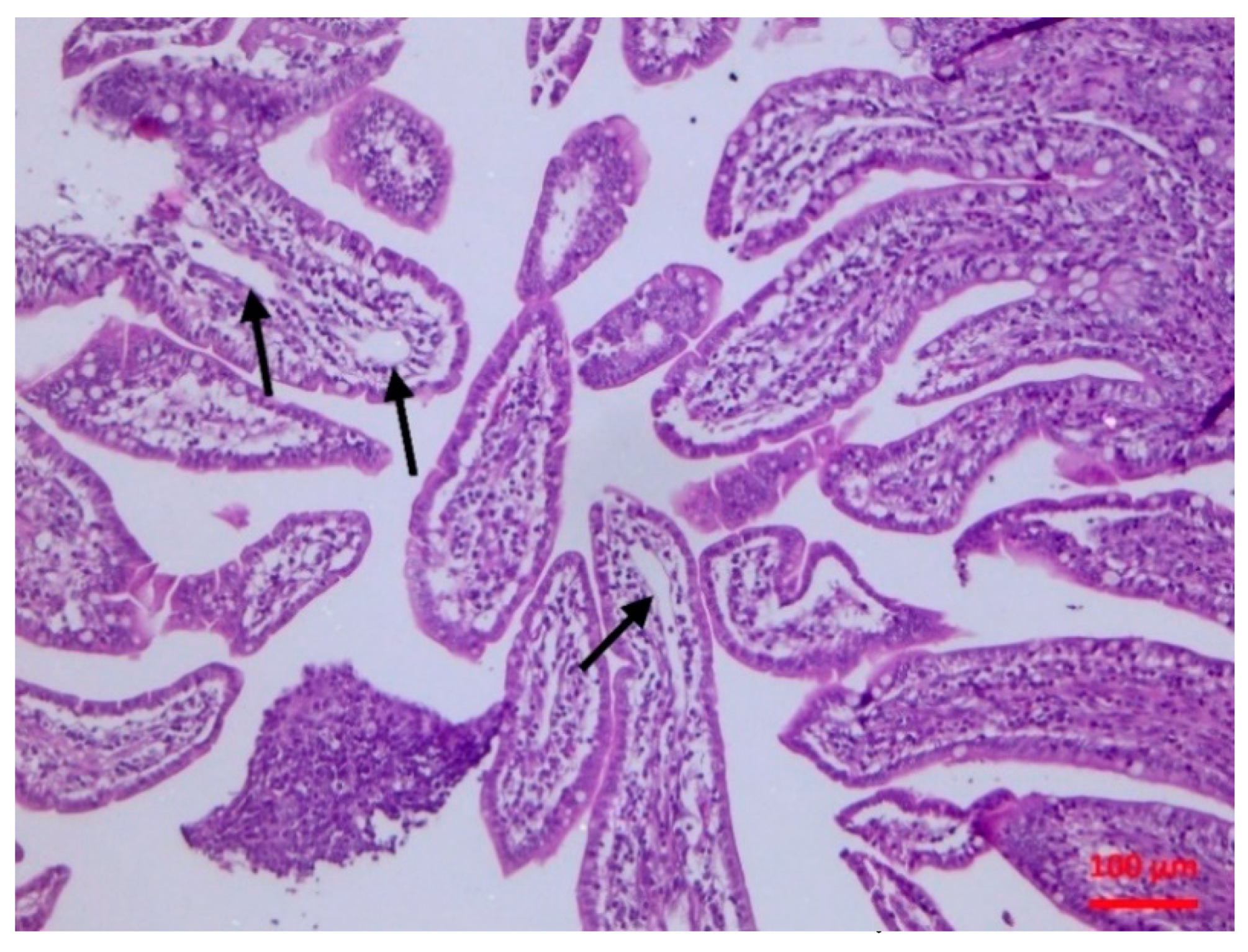
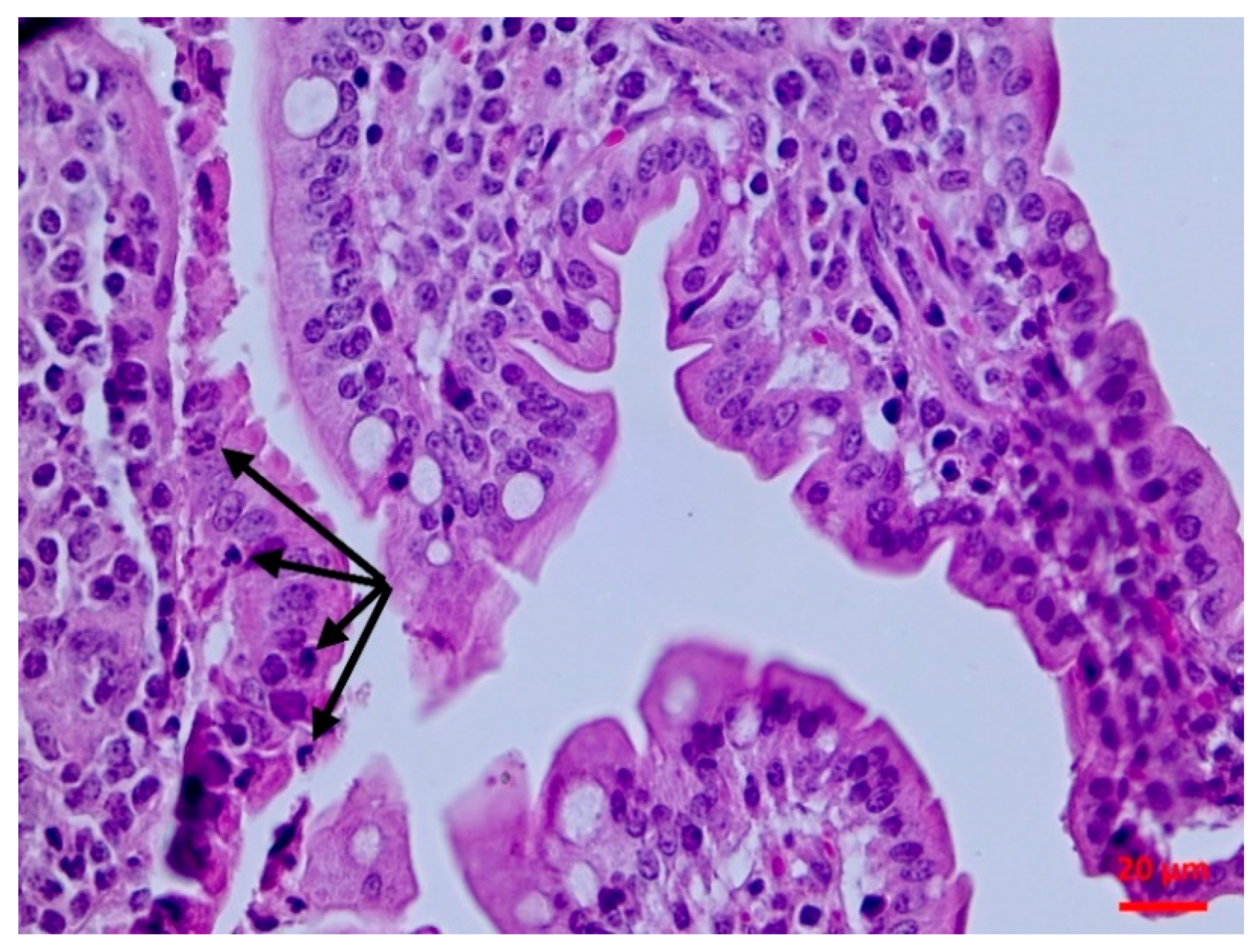
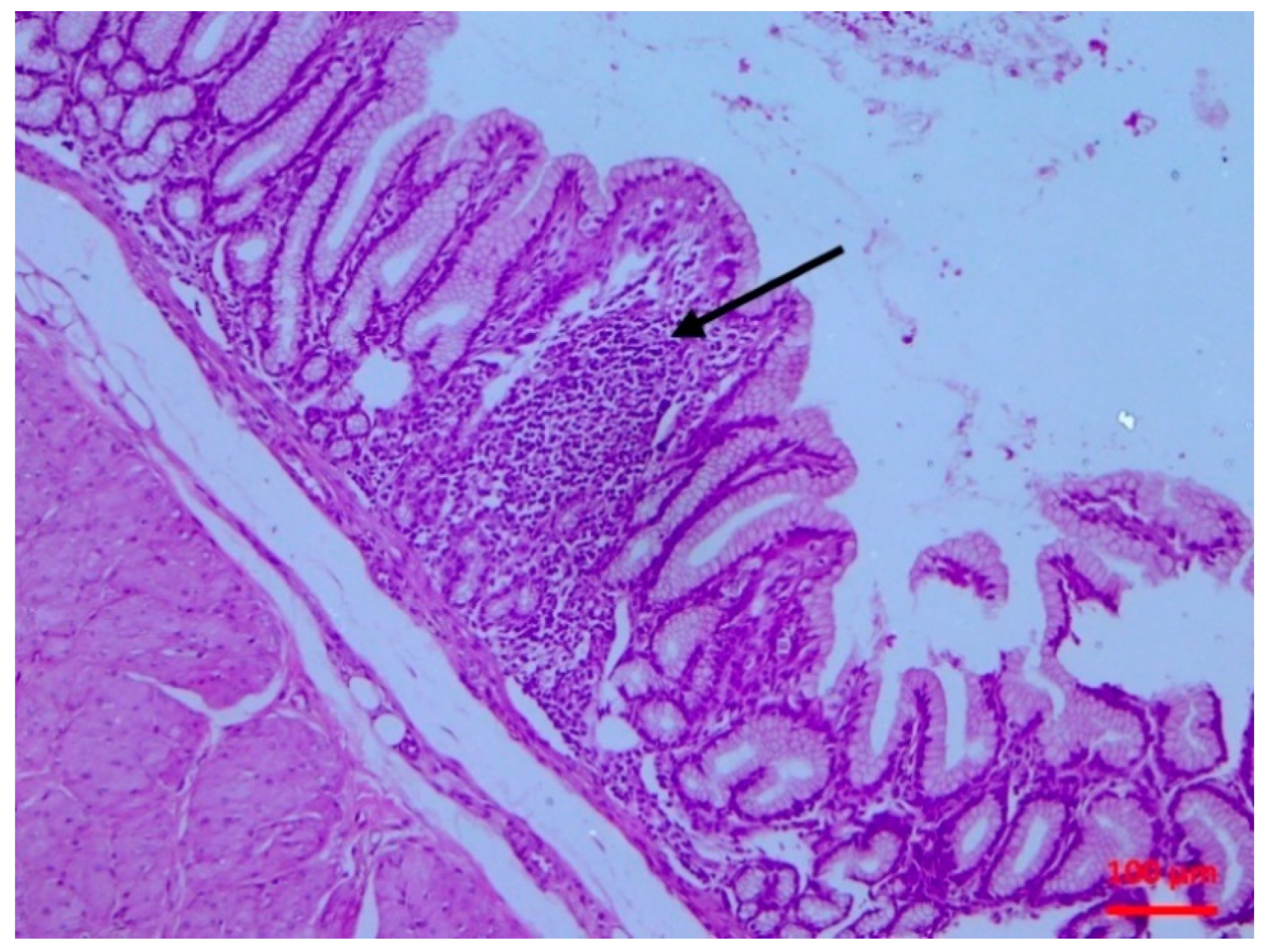
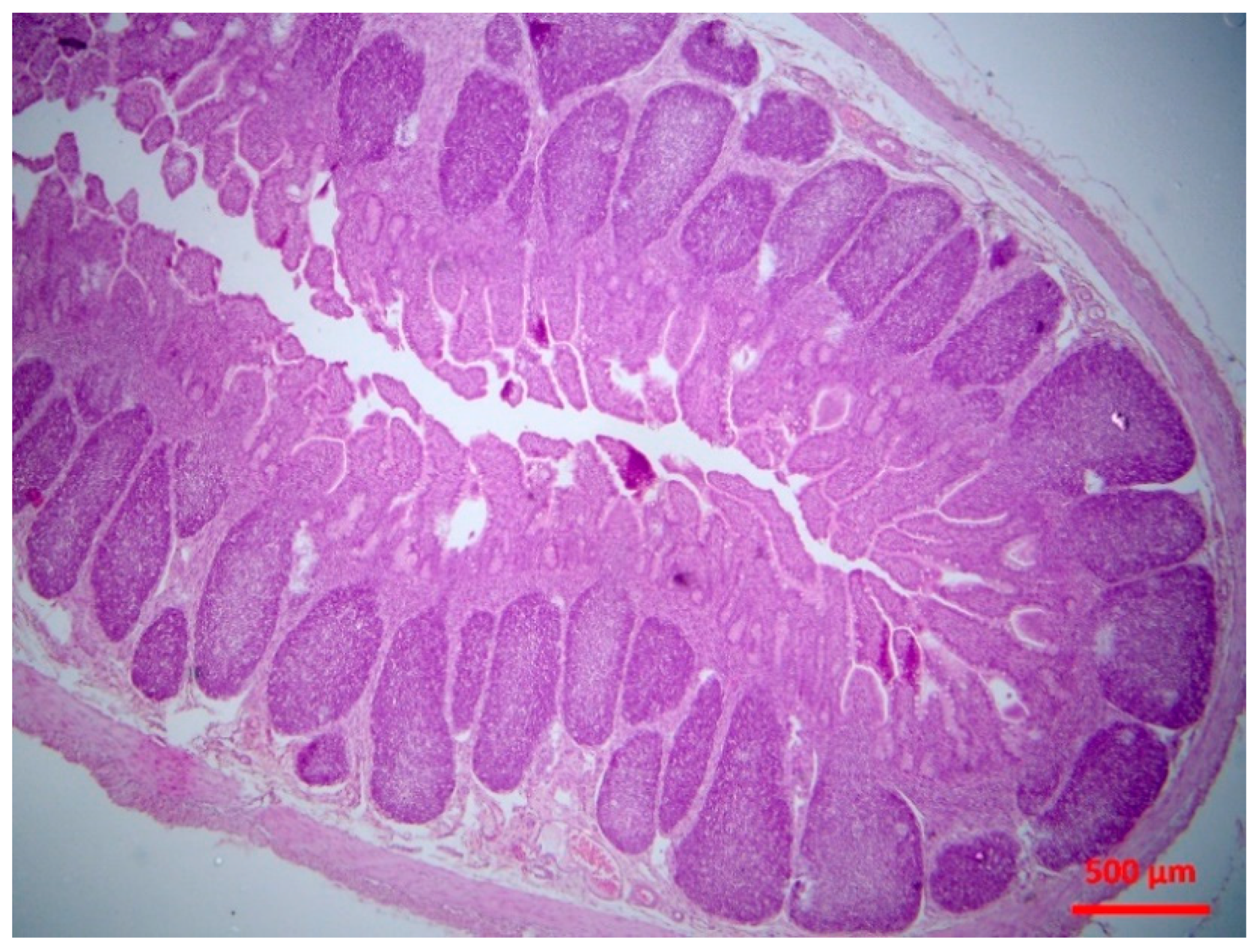
2.5.2. Histopathological findings
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Inoculation Procedure
4.3. Examination of Animals-Samplings
4.3.1. Lambs
4.3.2. Ewes
4.4. Laboratory Examinations
4.4.1. Haematological Examination
4.4.2. Bacteriological Examination
4.4.3. Molecular Examination for Presence of Salmonella spp.
DNA Extraction
PCR Amplification
4.4.4. Histopathological Examination
4.5. Data Management and Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Caldwell, M.E.; Ryerson, D.L. Salmonellosis in certain reptiles. J. Infect. Dis. 1939, 65, 242–245. [Google Scholar] [CrossRef]
- Tindall, B.J.; Grimont, P.A.D.; Garrity, G.M.; Euzeby, J. Nomenclature and taxonomy of the genus Salmonella. Int. J. System. Evol. Microbiol. 2005, 55, 521–524. [Google Scholar] [CrossRef] [PubMed]
- Grimont, P.A.D.; Weill, F.X. Antigenic Formulae of the Salmonella Serovars. In WHO Collaborating Center for Reference and Research on Salmonella, 9th ed.; Institut Pasteur: Paris, France, 2007. [Google Scholar]
- Lamas, A.; Miranda, J.M.; Regal, P.; Vázquez, B.; Franco, C.M.; Cepeda, A. A comprehensive review of non-enterica subspecies of Salmonella enterica. Microbiolog. Res. 2018, 206, 60–73. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, K.; Lassen-Nielsen, A.M.; Nordentoft, S.; Hammer, A.S. Serovars of Salmonella from captive reptiles. Zoon. Publ. Health 2009, 56, 238–242. [Google Scholar] [CrossRef]
- Lukac, M.; Pedersen, K.; Prukner-Radovcic, E. Prevalence of Salmonella in captive reptiles from Croatia. J. Zoo Wildl. Med. 2015, 46, 234–240. [Google Scholar] [CrossRef]
- Chiari, M.; Zanoni, M.; Tagliabue, S.; Lavazza, A.; Alborali, L.G. Salmonella serotypes in wild boars (Sus scrofa) hunted in northern Italy. Acta Vet. Scand. 2013, 55, 42. [Google Scholar] [CrossRef]
- Sanno, A.; Aspan, A.; Hestvik, G.; Jacobson, M. Presence of Salmonella spp., Yersinia enterocolitica, Yersinia pseudotuberculosis and Escherichia coli O157:H7 in wild boars. Epidemiol. Inf. 2014, 142, 2542–2547. [Google Scholar] [CrossRef]
- Hall, M.L.; Rowe, B. Arizona 26:29:30 in sheep in the United Kingdom. Vet. Rec. 1980, 107, 581–582. [Google Scholar]
- Davies, R.H.; Evans, S.J.; Perece, B.E.; Chappell, S.; Kidd, S.; Jones, Y.E. Increase in Salmonella enterica subspecies diarizonae serovar 61:k:1,5,(7) in sheep. Vet. Rec. 2001, 149, 555–557. [Google Scholar] [CrossRef]
- Alvseike, O.; Skjerve, E. Prevalence of Salmonella subspecies diarizonae in Norwegian sheep herds. Pr. Vet. Med. 2002, 52, 277–285. [Google Scholar] [CrossRef]
- Sandberg, M.; Alvseike, O.; Skjerve, E. The prevalence and dynamics of Salmonella enterica IIIb 61:k:1,5,(7) in sheep flocks in Norway. Pr. Vet. Med. 2002, 52, 267–275. [Google Scholar] [CrossRef]
- Methner, U.; Moog, U. Occurrence and characterisation of Salmonella enterica subspecies diarizonae serovar 61:k: 1, 5, (7) in sheep in the federal state of Thuringia, Germany. BMC Vet. Res. 2018, 14, 401. [Google Scholar] [CrossRef] [PubMed]
- Milnes, A.S.; Stewart, I.; Clifton-Hadley, F.A.; Davies, R.H.; Newell, D.G.; Sayers, A.R.; Cheasty, T.; Cassar, C.; Ridley, A.; Cook, A.J.C.; et al. Intestinal carriage of verocytotoxigenic Escherichia coli O157, Salmonella, thermophilic Campylobacter and Yersinia enterocolitica, in cattle, sheep and pigs at slaughter in Great Britain during 2003. Epidemiol. Inf. 2008, 136, 739–751. [Google Scholar] [CrossRef] [PubMed]
- Bonke, R.; Wacheck, S.; Bumann, C.; Thum, C.; Stüber, E.; König, M. High prevalence of Salmonella enterica subsp. diarizonae in tonsils of sheep at slaughter. Food Res. Int. 2012, 45, 880–884. [Google Scholar]
- Sörén, K.; Lindblad, M.; Jernberg, C.; Eriksson, E.; Melin, L.; Wahlström, H.; Lundh, M. Changes in the risk management of Salmonella enterica subspecies diarizonae serovar 61:(k):1,5,(7) in Swedish sheep herds and sheep meat due to the results of a prevalence study 2012. Acta Vet. Scand. 2015, 57, 6. [Google Scholar] [CrossRef][Green Version]
- Chatzopoulos, D.C. Significance of Salmonella enterica subsp. diarizonae in the Diarrhoeic Syndrome of Lambs. Ph.D. Thesis, University of Thessaly, Volos, Greece, 2019. [Google Scholar]
- Harp, J.A.; Myers, L.L.; Rich, J.E.; Gates, N.L. Role of Salmonella arizonae and other infective agents in enteric disease of lambs. Am. J. Vet. Res. 1981, 42, 596–599. [Google Scholar]
- Hannam, D.A.; Wray, C.; Harbourne, J.F. Experimental Salmonella arizonae infection of sheep. Br. Vet. J. 1986, 142, 458–466. [Google Scholar] [CrossRef]
- Chatzopoulos, D.C.; Sarrou, S.; Vasileiou, N.G.C.; Ioannidi, K.S.; Peteinaki, E.; Valiakos, G.; Tsokana, K.; Papadopoulos, E.; Spyrou, V.; Giannakopoulos, A.; et al. Dissemination of intestinal pathogens between lambs and puppies in sheep farms. Small Rumin. Res. 2016, 41, 5–10. [Google Scholar] [CrossRef]
- Greenfield, J.; Greenway, J.A.; Bigland, C.H. Arizona infections in sheep associated with gastroenteritis and abortion. Vet. Rec. 1973, 92, 400–401. [Google Scholar] [CrossRef]
- Meehan, J.T.; Brogden, K.A.; Courtney, C.; Cutlip, R.C.; Lehmkuhl, H.D. Chronic proliferative rhinitis associated with Salmonella arizonae in sheep. Vet. Pathol. 1992, 29, 556–559. [Google Scholar] [CrossRef]
- Brogden, K.; Meehan, J.; Lehmkuhl, H. Salmonella arizonae infection and colonisation of the upper respiratory tract of sheep. Vet. Rec. 1994, 135, 410–411. [Google Scholar] [CrossRef] [PubMed]
- Roger, P.A. The impact of disease and disease prevention on sheep welfare. Small Rumin. Res. 2008, 76, 104–111. [Google Scholar] [CrossRef]
- Henderson, D.C. Neonatal conditions. In Diseases of Sheep, 4th ed.; Aitken, I.D., Ed.; Blackwell: Oxford, UK, 2007; pp. 81–87. [Google Scholar]
- Katribe, E.; Bogomolnaya, L.M.; Wingert, H.; Andrews-Polymenis, H. Subspecies IIIa and IIIb Salmonellae are defective for colonization of murine models of salmonellosis compared to Salmonella enterica subsp. I serovar typhimurium. J. Bacteriol. 2009, 191, 2843–2850. [Google Scholar] [CrossRef] [PubMed]
- Lacasta, D.; Figueras, L.; Bueso, J.P.; De las Heras, M.; Ramos, J.; Ferrer, L.; González, J.; de Arcaute, M.R.; Ortín, A.; Marteles, D. Experimental infection with Salmonella enterica subsp. diarizonae serotype 61:k:1,5,(7) in sheep: Study of cell mediated immune response. Small Rumin. Res. 2017, 149, 28–33. [Google Scholar]
- Titchen, D.A. Cineradiographic studies of swallowing in the suckled lamb. Ann. Rech. Vet. 1977, 8, 483. [Google Scholar]
- Gougoulis, D.A.; Kyriazakis, I.; Tzora, A.; Taitzoglou, I.A.; Skoufos, J.; Fthenakis, G.C. Effects of lamb sucking on the bacterial flora of teat duct and mammary gland of ewes. Reprod. Dom. Anim. 2008, 43, 22–26. [Google Scholar]
- Kyriakis, C.S.; Billinis, C.; Papadopoulos, E.; Vasileiou, N.G.; Athanasiou, L.V.; Fthenakis, G.C. Bluetongue in small ruminants: An opinionated review, with a brief appraisal of the 2014 outbreak of the disease in Greece and the south-east Europe. Vet. Microbiol. 2015, 181, 66–74. [Google Scholar] [CrossRef]
- Vasileiou, N.G.C.; Fthenakis, G.C.; Amiridis, G.S.; Athanasiou, L.V.; Birtsas, P.; Chatzopoulos, D.C.; Chouzouris, T.M.; Giannakopoulos, A.; Ioannidi, K.S.; Kalonaki, S.N.; et al. Experiences from the 2014 outbreak of bluetongue in Greece. Small Rumin. Res. 2016, 142, 61–68. [Google Scholar] [CrossRef]
- Sunar, N.M.; Stentiford, E.I.; Stewart, D.I.; Fletcher, L.A. Molecular techniques to characterize the invA genes of Salmonella spp. for pathogen inactivation study in composting. arXiv 2014, 1404, 5208. [Google Scholar]
- Mejía, W.; Casal, J.; Zapata, D.; Sánchez, G.J.; Martín, M.; Mateu, E. Epidemiology of salmonella infections in pig units and antimicrobial susceptibility profiles of the strains of Salmonella species isolated. Vet. Rec. 2006, 159, 271–276. [Google Scholar] [CrossRef]
- Barbagianni, M.S.; Mavrogianni, V.S.; Katsafadou, A.I.; Spanos, S.A.; Tsioli, V.; Galatos, A.D.; Nakou, M.; Valasi, I.; Gouletsou, P.G.; Fthenakis, G.C. Pregnancy toxaemia as predisposing factor for development of mastitis in sheep during the immediately post-partum period. Small Rumin. Res. 2015, 130, 246–251. [Google Scholar] [CrossRef]
- Köhler, H.; Burkert, B.; Pavlik, I.; Diller, R.; Geue, L.; Conraths, F.J.; Martin, G. Evaluation of five ELISA test kits for the measurement of antibodies against Mycobacterium avium subspecies paratuberculosis in bovine serum. Berliner und Münch. Tierärztl. Wochensch. 2008, 21, 203–210. [Google Scholar]
- Vandenbussche, F.; Vanbinst, T.; Verheyden, B.; Van Dessel, W.; Demeestere, L.; Houdart, P.; Bertels, G.; Praet, N.; Berkvens, D.; Mintiens, K.; et al. Evaluation of antibody-ELISA and real-time RT-PCR for the diagnosis and profiling of bluetongue virus serotype 8 during the epidemic in Belgium in 2006. Vet. Microbiol. 2008, 129, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Michiels, R.; Van Mael, E.; Quinet, C.; Adjadj, N.R.; Cay, A.B.; De Regge, N. Comparative analysis of different serological and molecular tests for the detection of Small Ruminant Lentiviruses (SRLVs) in Belgian sheep and goats. Viruses 2018, 10, 696. [Google Scholar] [CrossRef] [PubMed]
- International Organization for Standardization. Microbiology of the Food Chain—Horizontal Method for the Detection, Enumeration and Serotyping of Salmonella—Part 1: Detection of Salmonella spp. EN ISO 6579-1; International Organization for Standardization: Geneva, Switzerland, 2017. [Google Scholar]
- Miles, A.A.; Misra, S.S.; Irwin, J.O. The estimation of the bactericidal power of the blood. J. Hyg. Camb. 1938, 38, 732–749. [Google Scholar] [CrossRef] [PubMed]
- Fthenakis, G.C. Prevalence and aetiology of subclinical mastitis in ewes of Southern Greece. Small Rumin. Res. 1994, 13, 293–300. [Google Scholar] [CrossRef]
- Athanasiou, L.V.; Giannakopoulos, C.G.; Polizopoulou, Z.S.; Katsogiannou, E.; Kontos, V.I. A comparative study of the ovine haemogram: Cell-Dyn 3500 versus manual methods. Am. J. Anim. Vet. Sci. 2013, 8, 203–209. [Google Scholar] [CrossRef]
- Rahn, K.; De Grandis, S.; Clarke, R.; McEwen, S.; Galan, J.; Ginocchio, C. Amplification of an invA gene sequence of Salmonella typhimurium by polymerase chain reaction as a specific method of detection of Salmonella spp. Mol. Cell. Probes 1992, 6, 271–279. [Google Scholar] [CrossRef]
- Mavrogianni, V.S.; Cripps, P.J.; Fthenakis, G.C. Bacterial flora and risk of infection of the ovine teat duct and mammary gland throughout lactation. Prev. Vet. Med. 2007, 79, 163–173. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Haematocrit (%) | Erythrocytes (×106 cells μL−1) | Haemoglobin (g dL−1) | MCV (fL) 1 | MCHC (g dL−1) 1 | ||||||
| Sampling occasion | Inoculated lambs | Uninfected controls | Inoculated lambs | Uninfected controls | Inoculated lambs | Uninfected controls | Inoculated lambs | Uninfected controls | Inoculated lambs | Uninfected controls |
| D0 | 33.8 | 26.1 | 8.2 | 10.2 | 10.5 | 11.1 | 40.4 | 45.2 | 12.8 | 12.4 |
| D0 + 6 h | 30.6 | 28.0 | 7.6 | 10.4 | 9.8 | 10.7 | 39.7 | 44.6 | 12.7 | 12.7 |
| D0 + 12 h | 28.9 | 28.1 | 7.3 | 11.1 | 9.5 | 10.9 | 39.0 | 44.2 | 12.7 | 12.9 |
| D1 | 30.0 | 28.3 | 7.7 | 11.4 | 9.4 | 11.1 | 39.2 | 45.0 | 12.4 | 12.4 |
| D2 | 30.4 | 27.7 | 7.7 | 10.7 | 9.7 | 10.6 | 39.6 | 44.2 | 12.7 | 11.0 |
| D4 | 28.1 | 28.9 | 7.2 | 11.0 | 9.0 | 11.4 | 38.7 | 41.2 | 12.3 | 11.0 |
| D7 | 28.9 | 29.1 | 7.7 | 11.9 | 9.1 | 12.0 | 35.1 | 41.7 | 11.9 | 11.6 |
| D10 | 28.6 | 29.2 | 8.1 | 9.8 | 8.8 | 11.4 | 35.2 | 40.4 | 10.9 | 10.5 |
| D14 | 27.8 | 28.3 | 8.7 | 9.9 | 8.3 | 10.5 | 32.7 | 40.7 | 9.8 | 9.6 |
| D21 | 34.4 | 28.7 | 10.5 | 10.1 | 10.3 | 10.5 | 33.2 | 41.3 | 10.0 | 10.3 |
| Total leucocytes (cells μL−1) | Neutrophils (cells μL−1) | Neutrophils (% leucocytes) | Lymphocytes (cells μL−1) | Lymphocytes (% leucocytes) | ||||||
| Sampling occasion | Inoculated lambs | Uninfected controls | Inoculated lambs | Uninfected controls | Inoculated lambs | Uninfected controls | Inoculated lambs | Uninfected controls | Inoculated lambs | Uninfected controls |
| D0 | 4740 | 6425 | 2100 | 3300 | 42.1 | 51.0 | 2610 | 2620 | 49.7 | 40.9 |
| D0 + 6 h | 4650 | 7010 | 1945 | 4040 | 37.9 | 57.6 | 2815 | 2540 | 55.3 | 36.2 |
| D0 + 12 h | 4720 | 8200 | 1460 | 5280 | 29.1 | 64.4 | 2720 | 2650 | 63.2 | 32.3 |
| D1 | 5270 | 5350 | 1550 | 3400 | 31.8 | 63.5 | 3330 | 1730 | 63.1 | 32.3 |
| D2 | 5300 | 5470 | 1790 | 1800 | 32.2 | 40.6 | 3440 | 2215 | 60.4 | 51.2 |
| D4 | 4970 | 5000 | 1480 | 730 | 29.5 | 14.6 | 3050 | 2000 | 60.8 | 80.0 |
| D7 | 4935 | 5085 | 1230 | 2255 | 27.2 | 44.4 | 3475 | 2555 | 66.1 | 50.0 |
| D10 | 5395 | 4650 | 990 | 1845 | 17.9 | 40.0 | 4025 | 2520 | 74.8 | 53.6 |
| D14 | 7280 | 4105 | 2330 | 1580 | 27.5 | 38.2 | 5260 | 2290 | 63.8 | 56.2 |
| D21 | 10,725 | 9490 | 3475 | 3250 | 34.3 | 34.4 | 5675 | 2325 | 53.0 | 55.8 |
| Monocytes (cells μL−1) | Monocytes (% leucocytes) | Eosinophils (cells μL−1) | Eosinophils (% leucocytes) | |||||||
| Sampling occasion | Inoculated lambs | Uninfected controls | Inoculated lambs | Uninfected controls | Inoculated lambs | Uninfected controls | Inoculated lambs | Uninfected controls | ||
| D0 | 155 | 65 | 2.6 | 0.9 | 50 | 285 | 1.0 | 4.6 | ||
| D0 + 6 h | 120 | 20 | 3.2 | 0.3 | 70 | 260 | 1.3 | 3.7 | ||
| D0 + 12 h | 60 | 20 | 1.6 | 0.2 | 40 | 110 | 1.0 | 1.3 | ||
| D1 | 60 | 20 | 1.5 | 0.4 | 60 | 70 | 1.5 | 1.2 | ||
| D2 | 245 | 180 | 4.0 | 4.0 | 40 | 45 | 0.7 | 1.0 | ||
| D4 | 80 | 20 | 1.6 | 0.4 | 50 | 40 | 1.0 | 0.8 | ||
| D7 | 70 | 100 | 1.3 | 2.0 | 50 | 45 | 0.9 | 0.9 | ||
| D10 | 115 | 100 | 1.8 | 2.2 | 35 | 85 | 0.8 | 2.1 | ||
| D14 | 190 | 50 | 2.1 | 1.2 | 130 | 120 | 1.7 | 2.2 | ||
| D21 | 150 | 105 | 1.4 | 1.1 | 1140 | 600 | 9.1 | 6.4 | ||
| Basophils (cells μL−1) | Basophils (% leucocytes) | Thrombocytes (cells μL−1) | ||||||||
| Sampling occasion | Inoculated lambs | Uninfected controls | Inoculated lambs | Uninfected controls | Inoculated lambs | Uninfected controls | ||||
| D0 | 90 | 110 | 1.9 | 1.7 | 545 | 763 | ||||
| D0 + 6 h | 120 | 100 | 2.1 | 1.4 | 556 | 893 | ||||
| D0 + 12 h | 100 | 90 | 2.0 | 1.0 | 512 | 844 | ||||
| D1 | 90 | 80 | 2.0 | 1.5 | 549 | 922 | ||||
| D2 | 90 | 70 | 1.9 | 1.7 | 598 | 1030 | ||||
| D4 | 110 | 180 | 2.2 | 3.6 | 934 | 1045 | ||||
| D7 | 120 | 95 | 2.6 | 1.8 | 1028 | 1770 | ||||
| D10 | 95 | 55 | 1.7 | 1.2 | 1146 | 1503 | ||||
| D14 | 90 | 45 | 1.2 | 1.1 | 1151 | 1167 | ||||
| D21 | 185 | 100 | 1.2 | 1.0 | 1082 | 1070 | ||||
| Before Challenge | Day After Challenge | Cumulative | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| D0 | D0 + 6 h | D0 + 12 h | D1 | D2 | D4 | D7 | D10 | D14 | D21 | |
| Faecal samples | ||||||||||
| 0/12 | 8/12 | 12/12 | 11/12 | 9/11 | 2/10 | 2/8 | 1/6 | 0/4 | 0/2 | 45/77 |
| Buccal samples | ||||||||||
| 0/12 | 0/12 | 0/12 | 6/12 | 2/11 | 1/10 | 1/8 | 0/6 | 0/4 | 0/2 | 10/77 |
| Tissue | Day After Challenge | Cumulative | ||||||
|---|---|---|---|---|---|---|---|---|
| D1 | D2 | D4 | D7 | D10 | D14 | D21 | ||
| small intestine | 1/1 | 1/1 | 0/2 | 1/2 | 1/2 | 2/2 | 0/2 | 6/12 |
| abomasum | 1/1 | 1/1 | 2/2 | 0/2 | 0/2 | 0/2 | 0/2 | 4/12 |
| liver | 0/1 | 0/1 | 1/2 | 0/2 | 0/2 | 1/2 | 1/2 | 3/12 |
| gallbladder | 0/1 | 0/1 | 1/2 | 1/2 | 0/2 | 0/2 | 0/2 | 2/12 |
| Cumulative | 2/4 | 2/4 | 4/8 | 2/8 | 1/8 | 3/8 | 1/8 | 15/48 |
| Recovery from Faecal Swab Samples | |||
|---|---|---|---|
| Yes | No | ||
| Recovery from Small Intestine Tissue Samples | Yes | 2 | 4 |
| No | 0 | 6 | |
| Recovery from Any Tissue Samples | Yes | 3 | 8 |
| No | 1 | 0 | |
| Before Challenge | Day After Challenge | Cumulative | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| D0 | D0 + 6 h | D0 + 12 h | D1 | D2 | D4 | D7 | D10 | D14 | D21 | |
| Faecal samples | ||||||||||
| 0/12 | 9/12 | 12/12 | 11/12 | 10/11 | 10/10 | 8/8 | 6/6 | 4/4 | 0/2 | 70/77 |
| Buccal samples | ||||||||||
| 0/12 | 3/12 | 3/12 | 8/12 | 5/11 | 3/10 | 2/8 | 0/6 | 0/4 | 0/2 | 24/77 |
| Animals | Type of Samples | No. of Animals in Samples from which Detected | No. of Samples in which Detected |
|---|---|---|---|
| Lambs | Faeces | 12/12 (1.000) | 70/77 (0.909) |
| Lambs | Buccal cavity swab | 9/12 (0.750) | 24/77 (0.312) |
| Ewes | Faeces | 0/6 (0.000) | 0/39 (0.000) |
| Ewes | Milk | 2/6 (0.333) | 3/78 (0.038) 1 |
| Lambs | Small intestine tissue | 12/12 (1.000) | 12/12 (1.000) |
| Lambs | Abomasum tissue | 8/12 (0.667) | 8/12 (0.667) |
| Lambs | Liver tissue | 12/12 (1.000) | 12/12 (1.000) |
| Lambs | Gallbladder tissue | 8/12 (0.667) | 8/12 (0.667) |
| Tissue | Day After Challenge | Cumulative | ||||||
|---|---|---|---|---|---|---|---|---|
| D1 | D2 | D4 | D7 | D10 | D14 | D21 | ||
| small intestine | 1/1 | 1/1 | 2/2 | 2/2 | 2/2 | 2/2 | 2/2 | 12/12 |
| abomasum | 1/1 | 1/1 | 2/2 | 2/2 | 1/2 | 1/2 | 0/2 | 10/12 |
| liver | 1/1 | 1/1 | 2/2 | 2/2 | 2/2 | 2/2 | 2/2 | 12/12 |
| gallbladder | 0/1 | 0/1 | 1/2 | 1/2 | 2/2 | 2/2 | 2/2 | 8/12 |
| Cumulative | 3/4 | 3/4 | 7/8 | 7/8 | 7/8 | 7/8 | 6/8 | 15/48 |
| Primer Sequence | Concentration (μM) | Product Size (bp) | AT 1 (°C) | Reference |
|---|---|---|---|---|
| Fw-GTGAAATTATCGCCACGTTCGGGCAA | 0.25 | 284 | 55.0 | [42] |
| Rv-TCATCGCACCGTCAAAGGAACC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chatzopoulos, D.C.; Vasileiou, N.G.C.; Ioannidi, K.S.; Katsafadou, A.I.; Mavrogianni, V.S.; Michael, C.K.; Katsarou, E.I.; Karavanis, E.; Papadopoulos, N.; Sbiraki, A.; et al. Experimental Study of the Potential Role of Salmonella enterica subsp. diarizonae in the Diarrhoeic Syndrome of Lambs. Pathogens 2021, 10, 113. https://doi.org/10.3390/pathogens10020113
Chatzopoulos DC, Vasileiou NGC, Ioannidi KS, Katsafadou AI, Mavrogianni VS, Michael CK, Katsarou EI, Karavanis E, Papadopoulos N, Sbiraki A, et al. Experimental Study of the Potential Role of Salmonella enterica subsp. diarizonae in the Diarrhoeic Syndrome of Lambs. Pathogens. 2021; 10(2):113. https://doi.org/10.3390/pathogens10020113
Chicago/Turabian StyleChatzopoulos, Dimitris C., Natalia G. C. Vasileiou, Katerina S. Ioannidi, Angeliki I. Katsafadou, Vasia S. Mavrogianni, Charalambia K. Michael, Eleni I. Katsarou, Emmanouil Karavanis, Nikolaos Papadopoulos, Afroditi Sbiraki, and et al. 2021. "Experimental Study of the Potential Role of Salmonella enterica subsp. diarizonae in the Diarrhoeic Syndrome of Lambs" Pathogens 10, no. 2: 113. https://doi.org/10.3390/pathogens10020113
APA StyleChatzopoulos, D. C., Vasileiou, N. G. C., Ioannidi, K. S., Katsafadou, A. I., Mavrogianni, V. S., Michael, C. K., Katsarou, E. I., Karavanis, E., Papadopoulos, N., Sbiraki, A., Athanasiou, L. V., Billinis, C., & Fthenakis, G. C. (2021). Experimental Study of the Potential Role of Salmonella enterica subsp. diarizonae in the Diarrhoeic Syndrome of Lambs. Pathogens, 10(2), 113. https://doi.org/10.3390/pathogens10020113