
Ascaris suum Nutrient Uptake and Metabolic Release, and Modulation of Host Intestinal Nutrient Transport by Excretory-Secretory and Cuticle Antigens In Vitro

 , ,
, , 

Abstract
:1. Introduction
2. Results
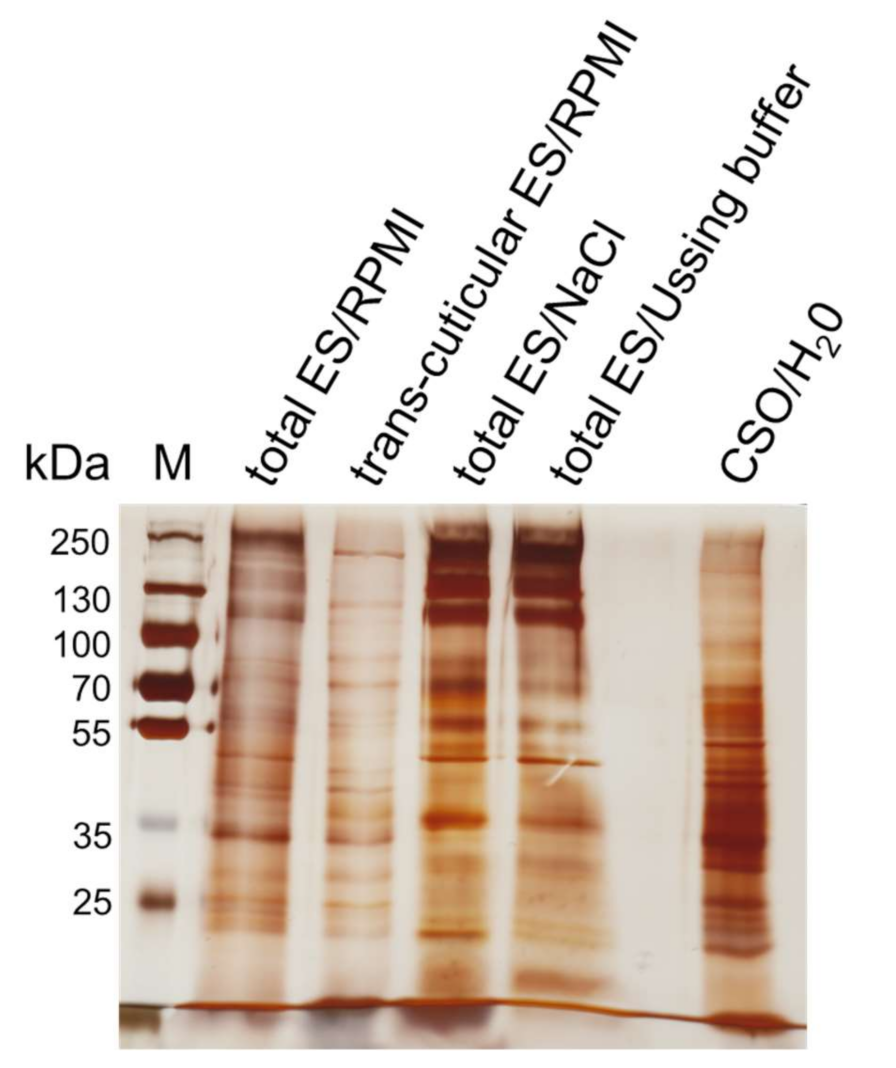
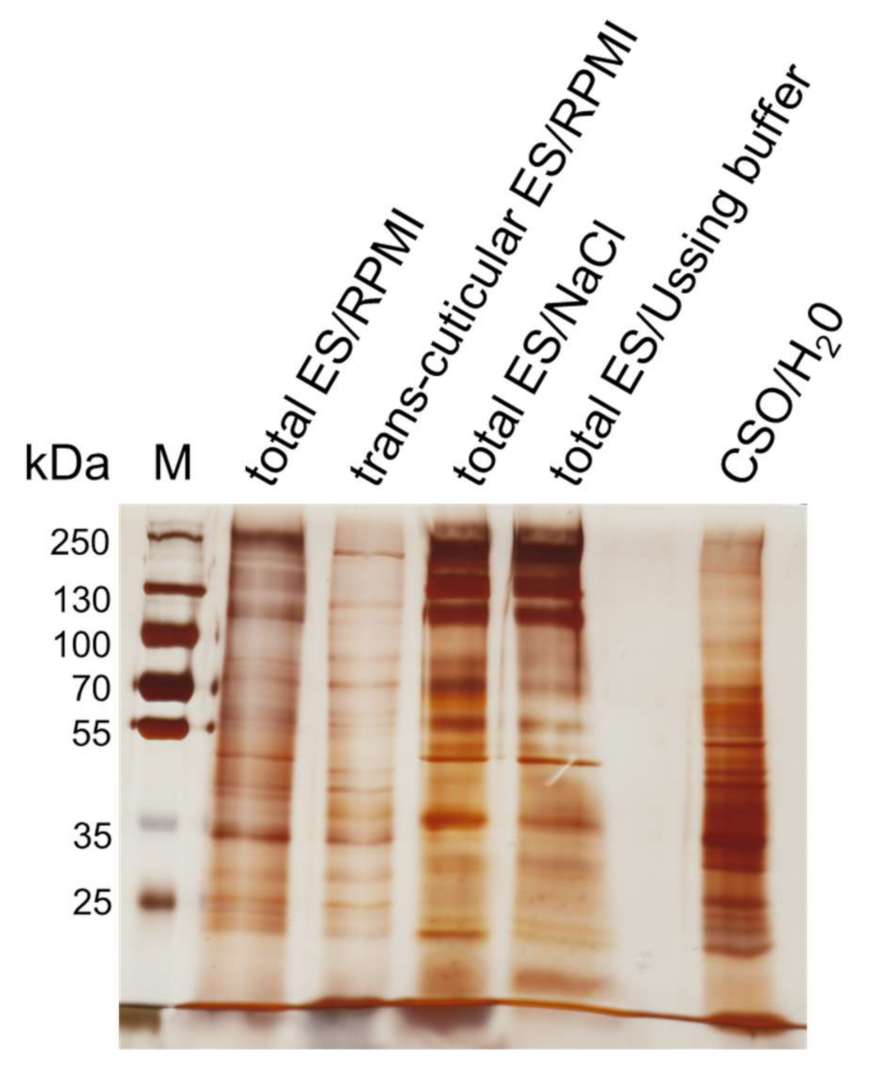
2.1. Nutrient Uptake and ES Pattern of In Vitro Cultivated Worms
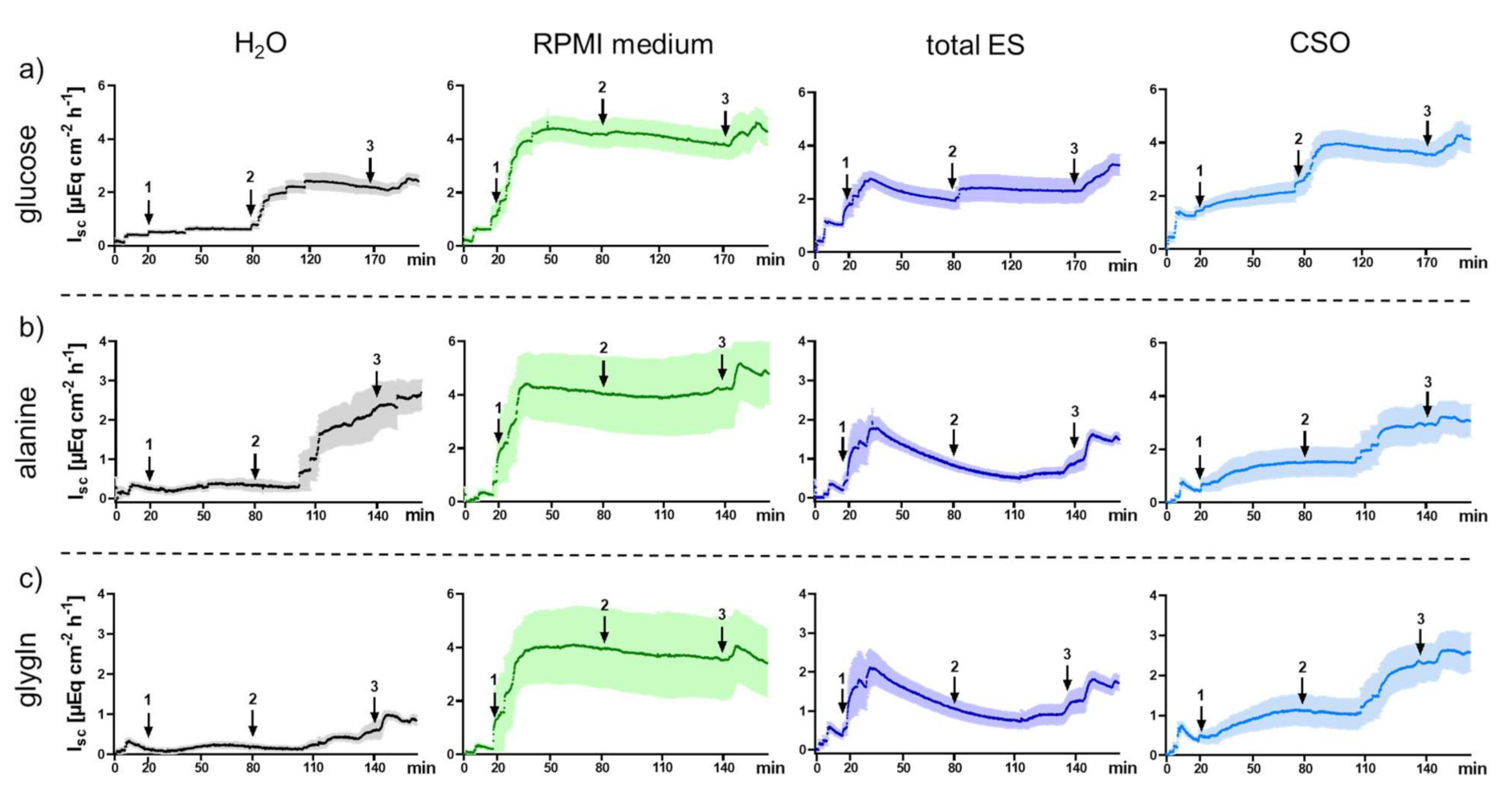
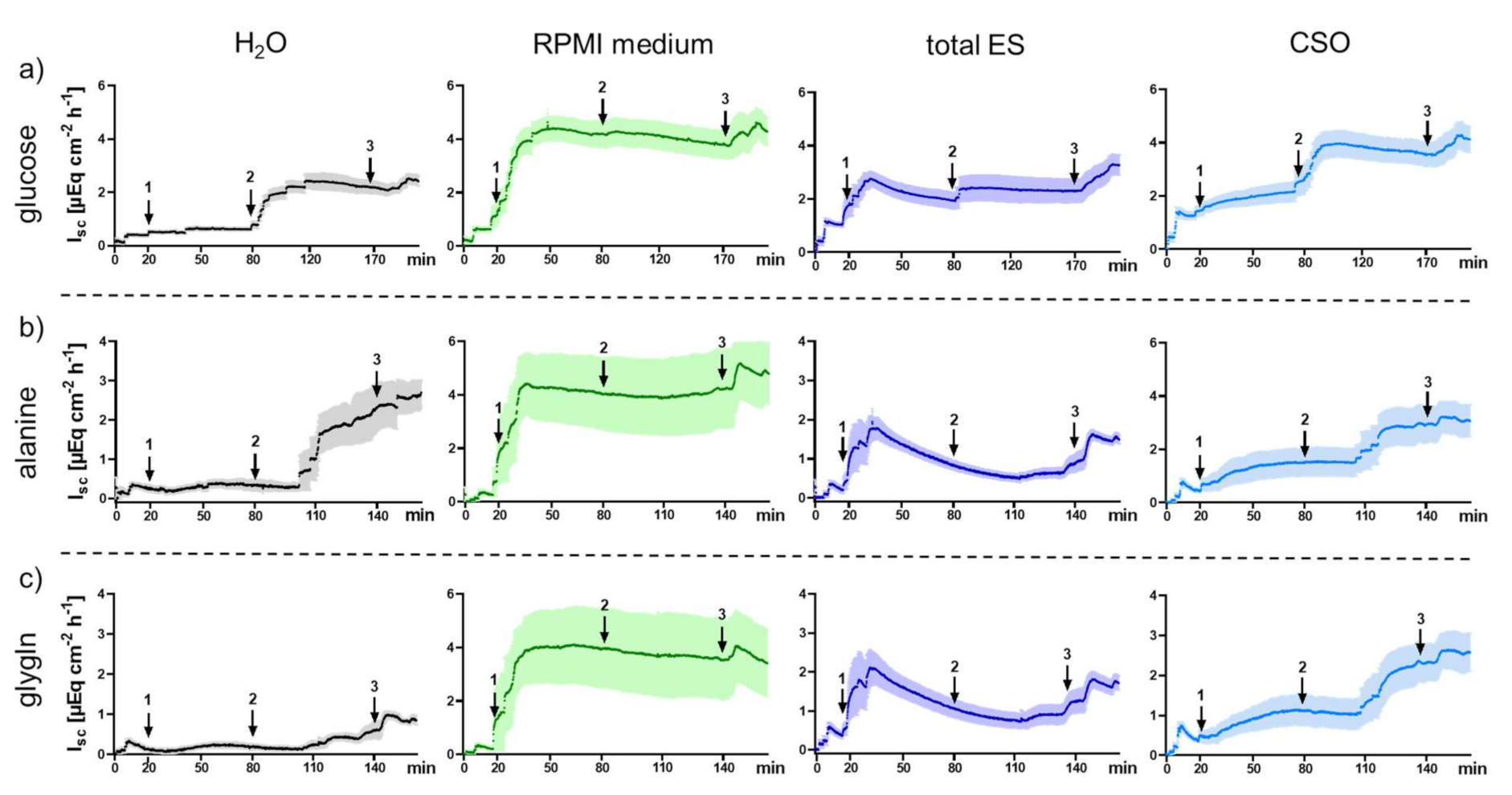
2.2. Functional Measurements upon Exposure of Porcine Jejunal Tissue to Adult A. suum Total ES and CSO Antigens
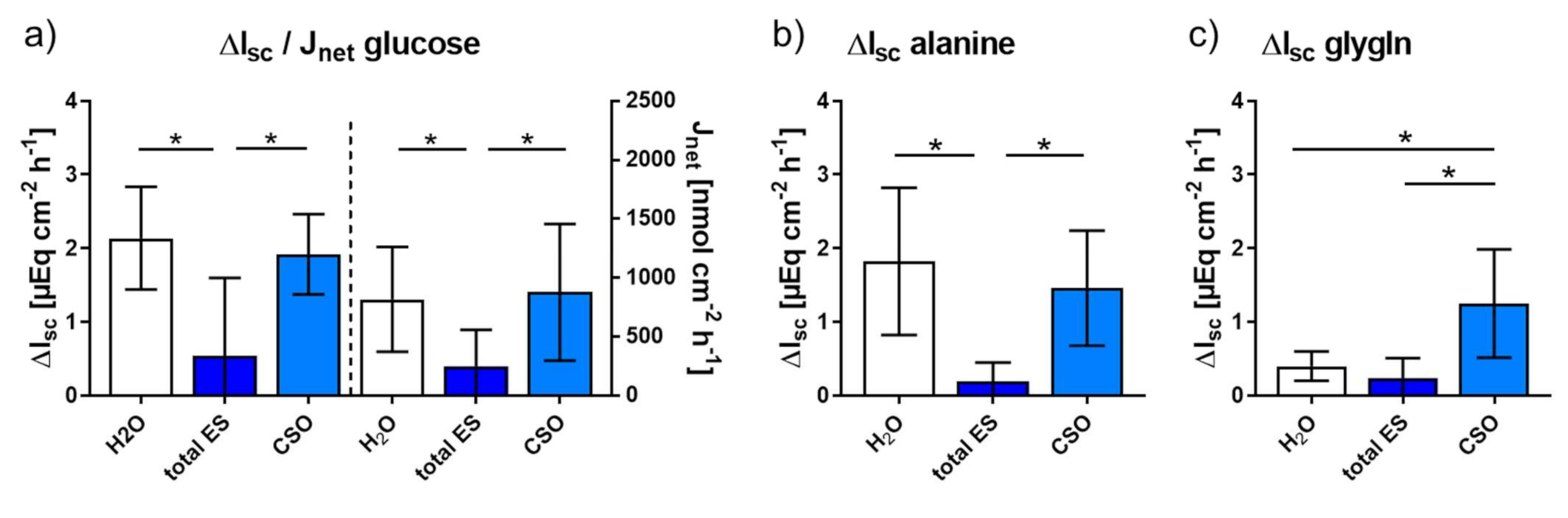
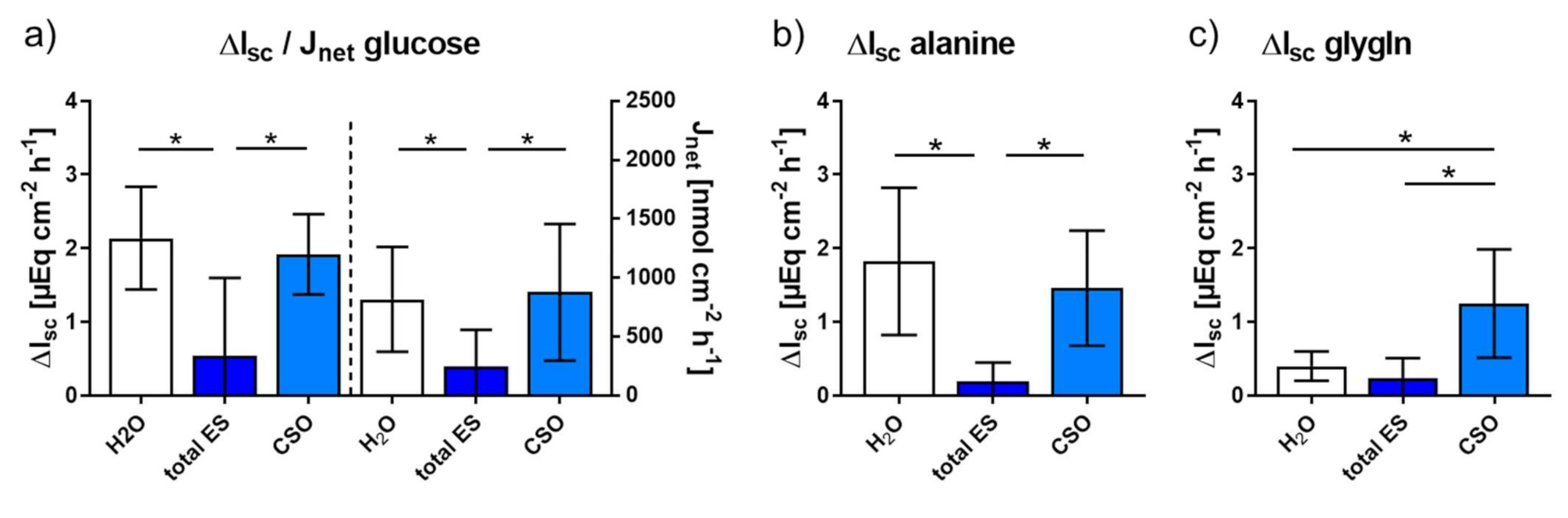
2.2.1. Impact of Adult A. suum Total ES and CSO Antigens on Jejunal Short-Circuit Current (Isc/ΔIsc) and 3H-D-Glucose Net Flux Rates (Jnet)
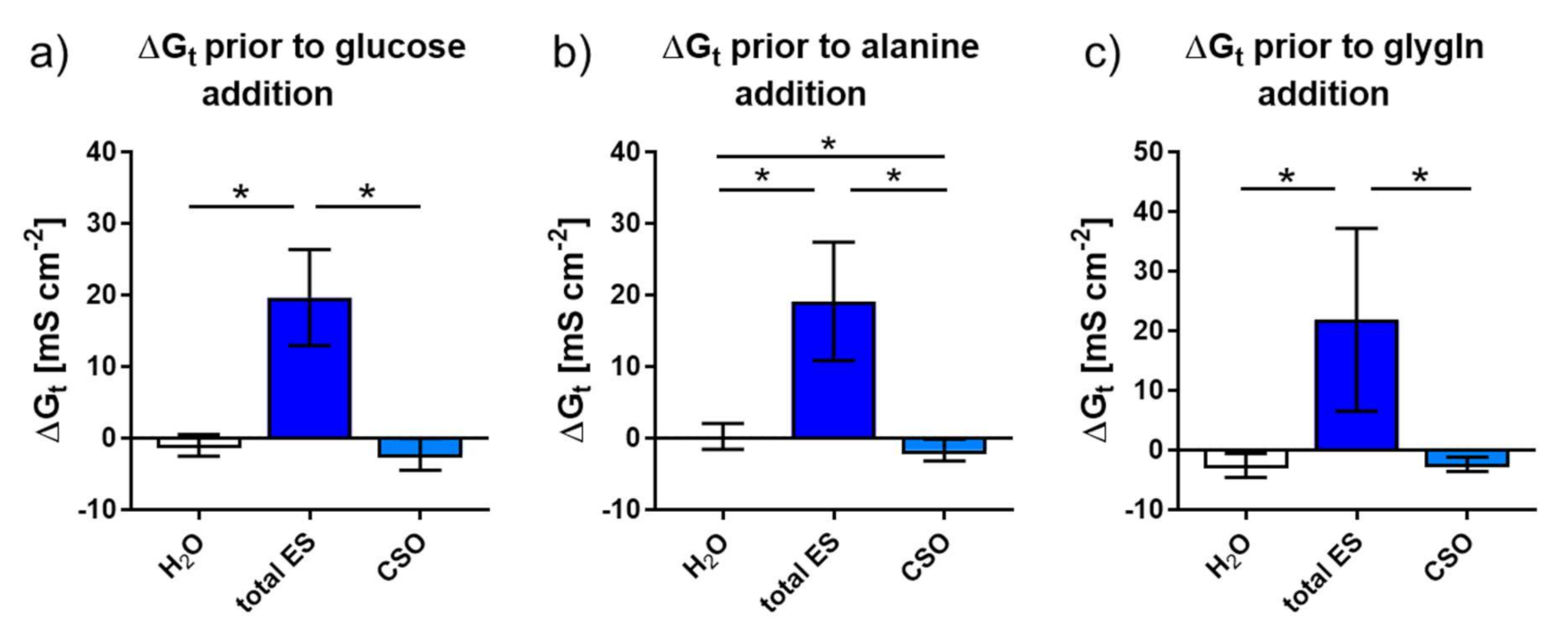
2.2.2. Tissue Conductance (Gt) in Response to Adult A. suum total ES and CSO Antigens
3. Discussion
4. Materials and Methods
4.1. Production of A. suum Total ES, Trans-Cuticular ES and CSO Antigens
4.2. Nutrient Uptake and ES Pattern of In Vitro Cultivated Adult Worms
4.3. Animals and Sample Collection
4.4. Ussing Chamber Measurements
4.5. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhou, C.; Guo, T.; Deng, Y.; He, J.; Ouyang, S.; Wu, X. Mitochondrial phylogenomics of human-type Ascaris, pig-type Ascaris, and hybrid Ascaris populations. Vet. Parasitol. 2020, 287, 109256. [Google Scholar] [CrossRef]
- Kurze, S.; Wesemeier, H.H. Ascaridiasis and discarded livers in Pigs—Data from veterinary practice and economic consequences. Prakt. Tierarzt 2006, 87, 128–137. [Google Scholar]
- Hale, O.M.; Stewart, T.B.; Marti, O.G. Influence of an experimental infection of Ascaris suum on performance of pigs. J. Anim. Sci. 1985, 60, 220–225. [Google Scholar] [CrossRef] [Green Version]
- Clark, P.; Bilkei, G. Production losses due to ascarid infestation of outdoor maintained pigs. Tierarztl Umschau 2003, 58, 425–431. [Google Scholar]
- Roepstorff, A.; Eriksen, L.; Slotved, H.C.; Nansen, P. Experimental Ascaris suum infection in the pig: Worm population kinetics following single inoculations with three doses of infective eggs. Parasitology 1997, 115 Pt 4, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Sinniah, B. Daily egg production of Ascaris lumbricoides: The distribution of eggs in the faeces and the variability of egg counts. Parasitology 1982, 84, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Dold, C.; Holland, C.V. Ascaris and ascariasis. Microbes Infect. 2011, 13, 632–637. [Google Scholar] [CrossRef]
- Fleming, M.W.; Fetterer, R.H. Ascaris suum: Continuous perfusion of the pseudocoelom and nutrient absorption. Exp. Parasitol. 1984, 57, 142–148. [Google Scholar] [CrossRef]
- Schwarz, A.; Gauly, M.; Abel, H.; Das, G.; Humburg, J.; Rohn, K.; Breves, G.; Rautenschlein, S. Immunopathogenesis of Ascaridia galli infection in layer chicken. Dev. Comp. Immunol. 2011, 35, 774–784. [Google Scholar] [CrossRef]
- Notari, L.; Riera, D.C.; Sun, R.; Bohl, J.A.; McLean, L.P.; Madden, K.B.; van Rooijen, N.; Vanuytsel, T.; Urban, J.F., Jr.; Zhao, A.; et al. Role of macrophages in the altered epithelial function during a type 2 immune response induced by enteric nematode infection. PLoS ONE 2014, 9, e84763. [Google Scholar] [CrossRef] [PubMed]
- Sekikawa, S.; Kawai, Y.; Fujiwara, A.; Takeda, K.; Tegoshi, T.; Uchikawa, R.; Yamada, M.; Arizono, N. Alterations in hexose, amino acid and peptide transporter expression in intestinal epithelial cells during Nippostrongylus brasiliensis infection in the rat. Int. J. Parasitol. 2003, 33, 1419–1426. [Google Scholar] [CrossRef]
- Dawson, H.D.; Beshah, E.; Nishi, S.; Solano-Aguilar, G.; Morimoto, M.; Zhao, A.; Madden, K.B.; Ledbetter, T.K.; Dubey, J.P.; Shea-Donohue, T.; et al. Localized multigene expression patterns support an evolving Th1/Th2-like paradigm in response to infections with Toxoplasma gondii and Ascaris suum. Infect. Immun. 2005, 73, 1116–1128. [Google Scholar] [CrossRef] [Green Version]
- Solano-Aguilar, G.; Shea-Donohue, T.; Madden, K.B.; Quinones, A.; Beshah, E.; Lakshman, S.; Xie, Y.; Dawson, H.; Urban, J.F. Bifidobacterium animalis subspecies lactis modulates the local immune response and glucose uptake in the small intestine of juvenile pigs infected with the parasitic nematode Ascaris suum. Gut Microbes 2018, 9, 422–436. [Google Scholar] [CrossRef]
- Kennedy, M.W.; Qureshi, F. Stage-specific secreted antigens of the parasitic larval stages of the nematode Ascaris. Immunology 1986, 58, 515–522. [Google Scholar]
- Andrade, M.A.; Siles-Lucas, M.; Lopez-Aban, J.; Carranza, C.; Perez-Arellano, J.L.; Muro, A. Antigens from Ascaris suum trigger in vitro macrophage NO production. Parasite Immunol. 2005, 27, 235–242. [Google Scholar] [CrossRef]
- Antunes, M.F.; Titz, T.O.; Batista, I.F.; Marques-Porto, R.; Oliveira, C.F.; Alves de Araujo, C.A.; Macedo-Soares, M.F. Immunosuppressive PAS-1 is an excretory/secretory protein released by larval and adult worms of the ascarid nematode Ascaris suum. J. Helminthol. 2015, 89, 367–374. [Google Scholar] [CrossRef]
- Lightowlers, M.W.; Rickard, M.D. Excretory-secretory products of helminth parasites: Effects on host immune responses. Parasitology 1988, 96, S123–S166. [Google Scholar] [CrossRef] [PubMed]
- Diosdado, A.; Simon, F.; Morchon, R.; Gonzalez-Miguel, J. Host-parasite relationships in porcine ascariosis: Anticoagulant potential of the third larval stage of Ascaris suum as a possible survival mechanism. Animals 2021, 11, 804. [Google Scholar] [CrossRef]
- Leles, D.; Gardner, S.L.; Reinhard, K.; Iniguez, A.; Araujo, A. Are Ascaris lumbricoides and Ascaris suum a single species? Parasit Vectors 2012, 5, 42. [Google Scholar] [CrossRef] [Green Version]
- Dass, P.D.; Donahue, M.J. Gamma-Glutamyl transpeptidase activity in Ascaris suum. Mol. Biochem. Parasitol. 1986, 20, 233–236. [Google Scholar] [CrossRef]
- Hussein, A.S.; Walter, R.D. Purification and characterization of gamma-glutamyl transpeptidase from Ascaris Suum. Mol. Biochem. Parasitol. 1996, 77, 41–47. [Google Scholar] [CrossRef]
- Van den Bossche, H.; De Nollin, S. Effects of mebendazole on the absorption of low molecular weight nutrients by Ascaris suum. Int. J. Parasitol. 1973, 3, 401–407. [Google Scholar] [CrossRef]
- Ho, N.F.; Geary, T.G.; Raub, T.J.; Barsuhn, C.L.; Thompson, D.P. Biophysical transport properties of the cuticle of Ascaris suum. Mol. Biochem. Parasitol. 1990, 41, 153–165. [Google Scholar] [CrossRef]
- Pollak, J.K.; Fairbairn, D. The metabolism of Ascaris lumbricoides ovaries. I. Nitrogen distribution. Can. J. Biochem. Physiol. 1955, 33, 297–306. [Google Scholar] [CrossRef]
- Pollak, J.K.; Fairbairn, D. The metabolism of Ascaris lumbricoides ovaries. II. Amino acid metabolism. Can. J. Biochem. Physiol. 1955, 33, 307–316. [Google Scholar] [CrossRef]
- Abbas, M.; Foor, W.E. Ascaris suum: Free amino acids and proteins in the pseudocoelom, seminal vesicle, and glandular vas deferens. Exp. Parasitol. 1978, 45, 263–273. [Google Scholar] [CrossRef]
- Donahue, M.J.; Yacoub, N.J.; Kaeini, M.R.; Harris, B.G. Activity of enzymes regulating glycogen metabolism in perfused muscle-cuticle sections of Ascaris suum (Nematoda). J. Parasitol. 1981, 67, 362–367. [Google Scholar] [CrossRef] [PubMed]
- Rasero, F.S.; Monteoliva, M.; Mayor, F. Enzymes related to 4-aminobutyrate metabolism in intestinal parasites. Comp. Biochem. Physiol. 1968, 25, 693–701. [Google Scholar] [CrossRef]
- Donahue, M.J.; Yacoub, N.J.; Harris, B.G. Correlation of muscle-activity with glycogen-metabolism in muscle of Ascaris suum. Am. J. Physiol. 1982, 242, R514–R521. [Google Scholar] [CrossRef]
- Barrett, J. The anaerobic end-products of helminths. Parasitology 1984, 88 Pt 1, 179–198. [Google Scholar] [CrossRef] [PubMed]
- Cavier, R.; Savel, J. Ammoniacal or ureic protein metabolism in Ascaris lumbricoides Linne, 1758. C R Hebd. Seances Acad. Sci. 1954, 238, 2448–2450. [Google Scholar]
- Barrett, J. Amino acid metabolism in helminths. Adv. Parasitol. 1991, 30, 39–105. [Google Scholar] [CrossRef]
- Arthur, S.; Sundaram, U. Protein kinase C-mediated phosphorylation of RKIP regulates inhibition of Na-alanine cotransport by leukotriene D(4) in intestinal epithelial cells. Am. J. Physiol. Cell Physiol. 2014, 307, C1010–C1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramanian, S.; Glitz, P.; Kipp, H.; Kinne, R.K.; Castaneda, F. Protein kinase-A affects sorting and conformation of the sodium-dependent glucose co-transporter SGLT1. J. Cell Biochem. 2009, 106, 444–452. [Google Scholar] [CrossRef]
- Arthur, S.; Coon, S.; Kekuda, R.; Sundaram, U. Regulation of sodium glucose co-transporter SGLT1 through altered glycosylation in the intestinal epithelial cells. BBA-Biomembranes 2014, 1838, 1208–1214. [Google Scholar] [CrossRef] [Green Version]
- Thwaites, D.T.; Ford, D.; Glanville, M.; Simmons, N.L. H+/solute-induced intracellular acidification leads to selective activation of apical Na+/H+ exchange in human intestinal epithelial cells. J. Clin. Investig. 1999, 104, 629–635. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, D.J.; Leibach, F.H.; Ganapathy, V.; Thwaites, D.T. Optimal absorptive transport of the dipeptide glycylsarcosine is dependent on functional Na+/H+ exchange activity. Pflug. Arch. Eur. J. Phys. 2002, 445, 139–146. [Google Scholar] [CrossRef]
- Becker, A.C.; Kraemer, A.; Epe, C.; Strube, C. Sensitivity and efficiency of selected coproscopical methods-sedimentation, combined zinc sulfate sedimentation-flotation, and McMaster method. Parasitol. Res. 2016, 115, 2581–2587. [Google Scholar] [CrossRef]
- Wossene, A.; Tsuji, N.; Kasuga-Aoki, H.; Miyoshi, T.; Isobe, T.; Arakawa, T.; Matsumoto, Y.; Yoshihara, S. Lung-stage protein profile and antigenic relationship between Ascaris lumbricoides and Ascaris suum. J. Parasitol. 2002, 88, 826–828. [Google Scholar] [CrossRef]
- Clarke, L.L. A guide to Ussing chamber studies of mouse intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G1151–G1166. [Google Scholar] [CrossRef] [Green Version]
- Atisook, K.; Carlson, S.; Madara, J.L. Effects of phlorizin and sodium on glucose-elicited alterations of cell junctions in intestinal epithelia. Am. J. Physiol. 1990, 258, C77–C85. [Google Scholar] [CrossRef] [PubMed]
- Schultz, S.G.; Zalusky, R. Ion Transport in Isolated Rabbit Ileum. I. Short-Circuit Current and Na Fluxes. J. Gen. Physiol. 1964, 47, 567–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winckler, C.; Breves, G.; Boll, M.; Daniel, H. Characteristics of dipeptide transport in pig jejunum in vitro. J. Comp. Physiol. B 1999, 169, 495–500. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analyte | Native Lyophilized RPMI (mmol/L) | After Cultivation of Untreated Worms (mmol/L) | % Difference | After Cultivation of Sealed Worms (mmol/L) | % Difference |
|---|---|---|---|---|---|
| Glucose | 325.82 | <0.01 | >99.9 | 28.66 | 91.2 |
| Alanine | <0.01 | 1.22 | >99.2 | 3.17 | >99.7 |
| γ-aminobutyric acid (GABA) | <0.01 | 0.18 | >94.4 | 1.42 | >99.3 |
| Arginine | 20.44 | 6.22 | 69.6 | 16.26 | 20.5 |
| Asparagine | 5.68 | <0.01 | >99.8 | 0.06 | 98.9 |
| Aspartic acid | 2.23 | <0.01 | >99.6 | 0.56 | 74.9 |
| Glutamic acid | 1.81 | 1.62 | 10.5 | 7.77 | 76.7 |
| Glutamine | 29.26 | 4.94 | 83.1 | 14.14 | 51.7 |
| Glycine | 1.92 | 0.32 | 83.3 | 0.79 | 58.9 |
| Histidine | 1.21 | 0.54 | 55.4 | 0.91 | 24.8 |
| Isoleucine | 5.44 | 2.42 | 55.5 | 4.10 | 24.6 |
| Leucine | 5.38 | 1.88 | 65.1 | 3.76 | 30.1 |
| Lysine | 2.96 | 0.15 | 94.9 | 0.93 | 68.6 |
| Methionine | 1.16 | 0.23 | 80.2 | 0.66 | 43.1 |
| Ornithine | <0.01 | 1.37 | >99.3 | 0.40 | >97.5 |
| Phenylalanine | 1.17 | 0.34 | 70.9 | 0.64 | 45.3 |
| Proline | 1.97 | 0.92 | 53.3 | 1.76 | 10.7 |
| Serine | 3.92 | <0.01 | >99.7 | 0.05 | 98.7 |
| Threonine | 2.31 | <0.01 | >99.6 | 0.39 | 83.1 |
| Tyrosine | 1.40 | 0.36 | 74.3 | 0.91 | 35.0 |
| Valine | 1.87 | 1.08 | 42.2 | 1.58 | 15.5 |
| Ammonia | 48.5 mg/L | 471.4 mg/L | 89.7 | 377.2 mg/L | 87.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koehler, S.; Springer, A.; Issel, N.; Klinger, S.; Wendt, M.; Breves, G.; Strube, C. Ascaris suum Nutrient Uptake and Metabolic Release, and Modulation of Host Intestinal Nutrient Transport by Excretory-Secretory and Cuticle Antigens In Vitro. Pathogens 2021, 10, 1419. https://doi.org/10.3390/pathogens10111419
Koehler S, Springer A, Issel N, Klinger S, Wendt M, Breves G, Strube C. Ascaris suum Nutrient Uptake and Metabolic Release, and Modulation of Host Intestinal Nutrient Transport by Excretory-Secretory and Cuticle Antigens In Vitro. Pathogens. 2021; 10(11):1419. https://doi.org/10.3390/pathogens10111419
Chicago/Turabian StyleKoehler, Sarina, Andrea Springer, Nicole Issel, Stefanie Klinger, Michael Wendt, Gerhard Breves, and Christina Strube. 2021. "Ascaris suum Nutrient Uptake and Metabolic Release, and Modulation of Host Intestinal Nutrient Transport by Excretory-Secretory and Cuticle Antigens In Vitro" Pathogens 10, no. 11: 1419. https://doi.org/10.3390/pathogens10111419
APA StyleKoehler, S., Springer, A., Issel, N., Klinger, S., Wendt, M., Breves, G., & Strube, C. (2021). Ascaris suum Nutrient Uptake and Metabolic Release, and Modulation of Host Intestinal Nutrient Transport by Excretory-Secretory and Cuticle Antigens In Vitro. Pathogens, 10(11), 1419. https://doi.org/10.3390/pathogens10111419