
Molecular Identification of Selected Tick-Borne Protozoan and Bacterial Pathogens in Thoroughbred Racehorses in Cavite, Philippines

 ,
, 

 ,
, 
Abstract
1. Introduction
2. Results
2.1. Sample Population and Demographics
2.2. Detected Tick-Borne Pathogens
2.3. Identifying Risk Factors Associated with TBP Detection
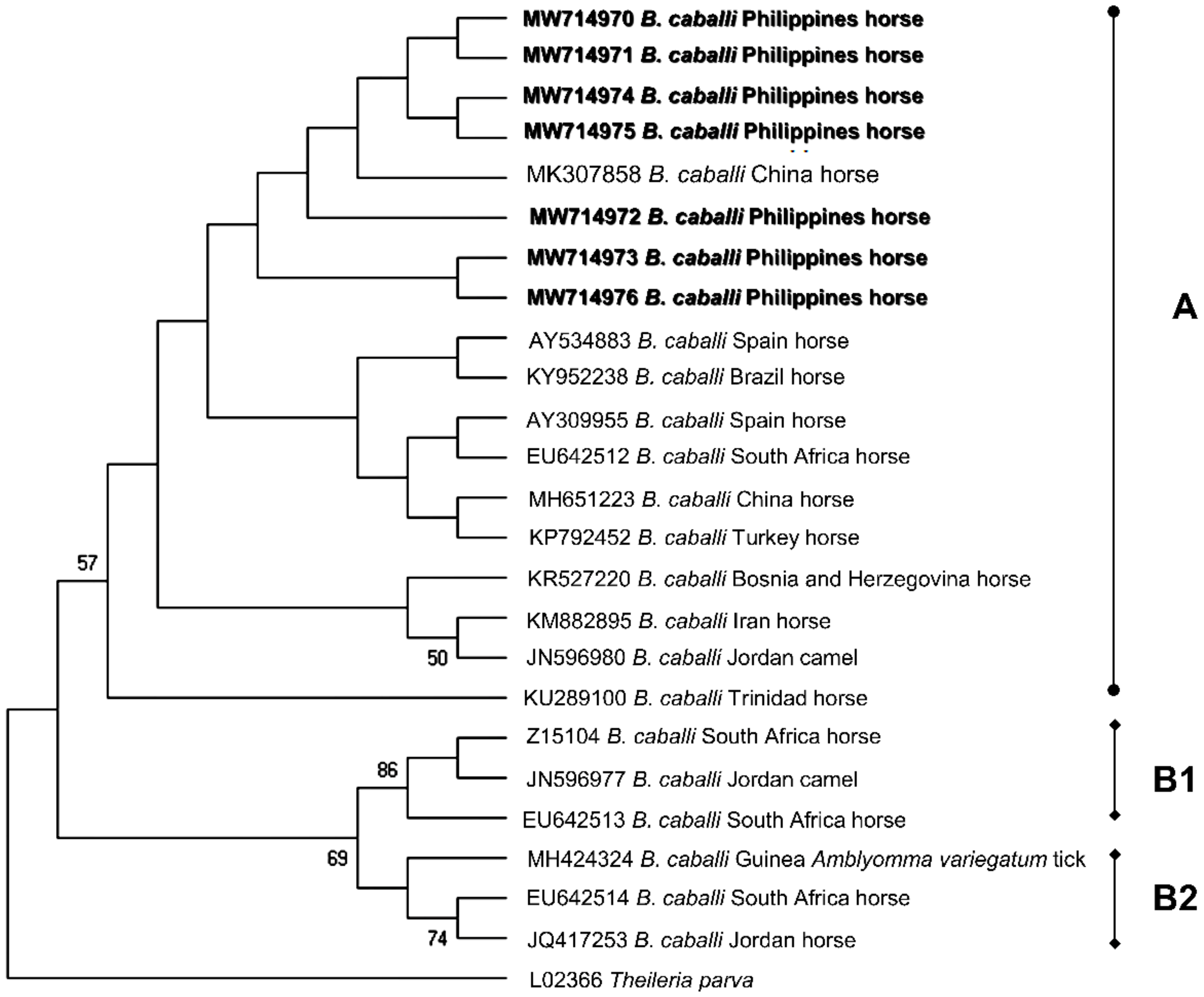
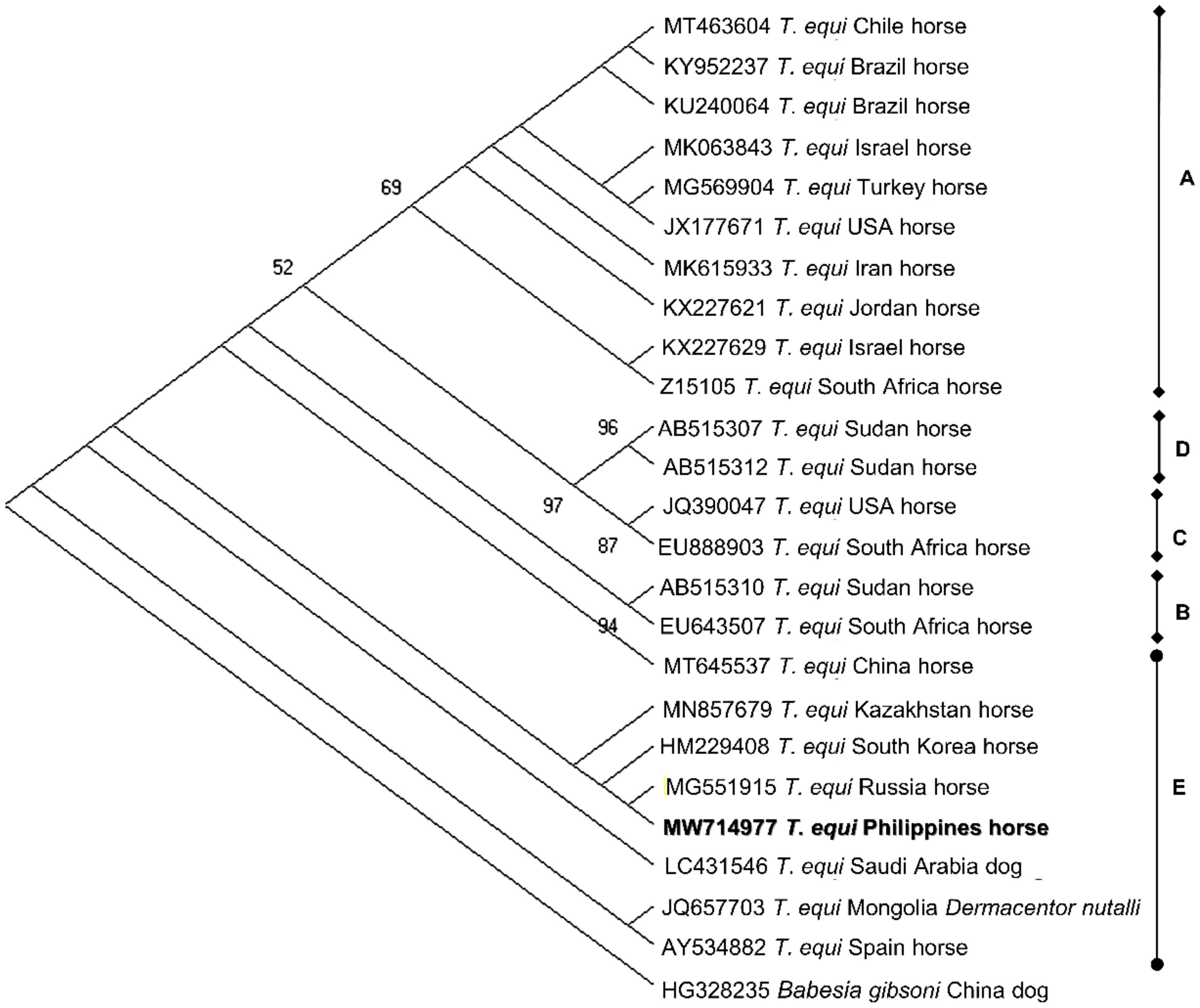
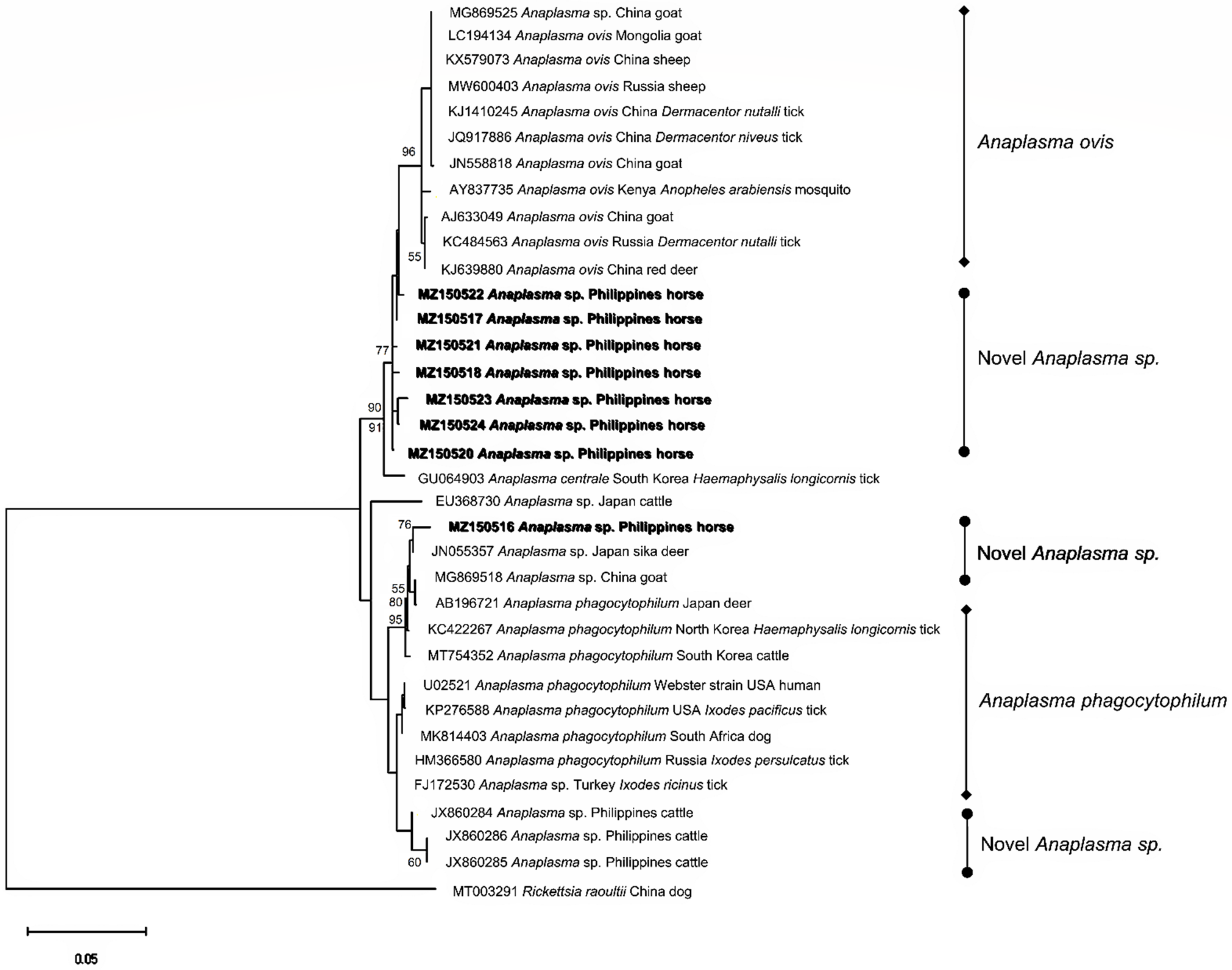
2.4. Sequence Identities and Phylogenetic Analysis
3. Discussion
4. Materials and Methods
4.1. Ethical Statements
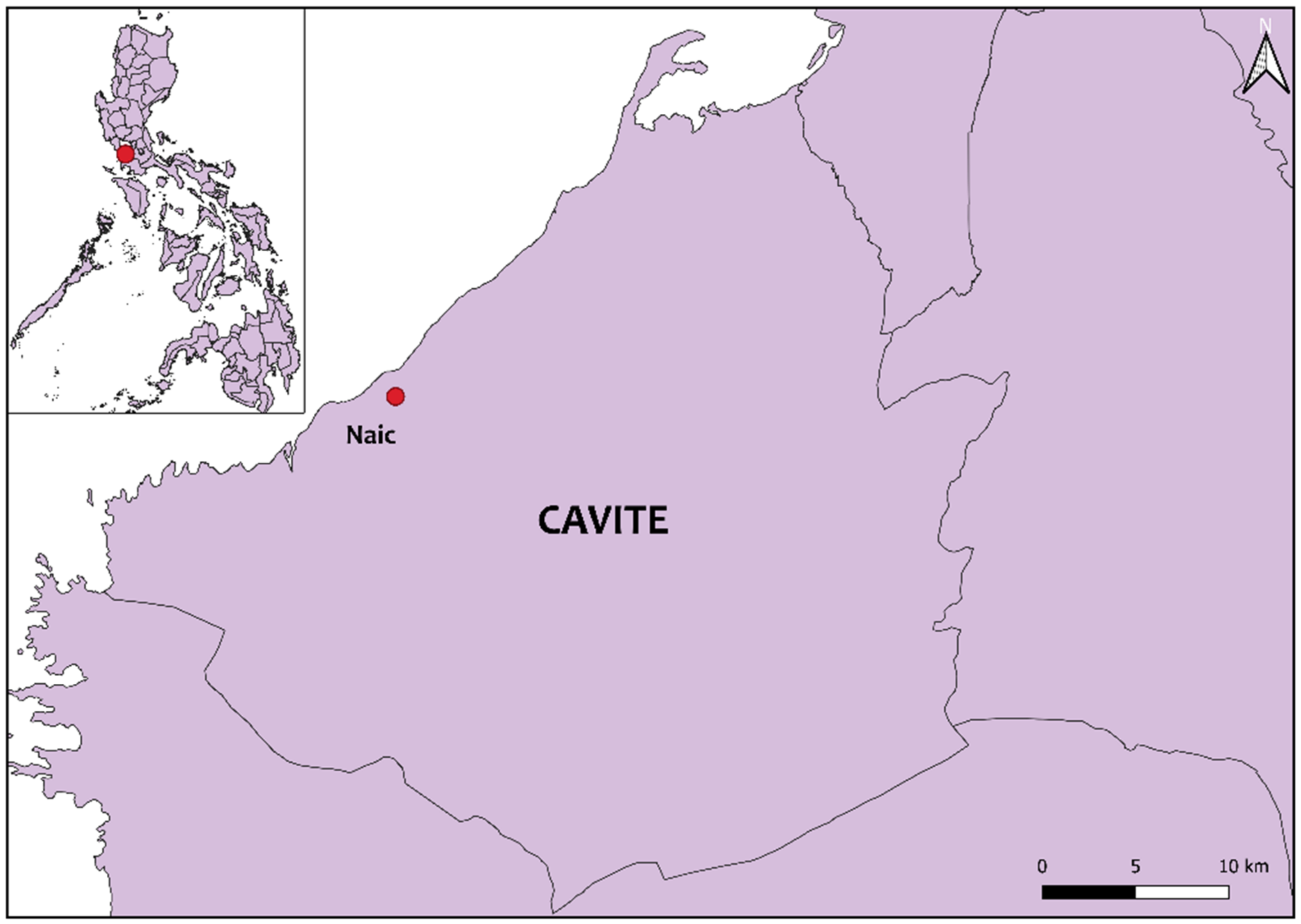
4.2. Location of Sampling and Animals
4.3. Extraction of DNA
4.4. PCR Assays for the Identification of Tick-Borne Pathogens
4.5. Sequencing and Phylogenetic Analyses
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Onyiche, T.E.; Suganuma, K.; Igarashi, I.; Yokoyama, N.; Xuan, X.; Thekisoe, O. A review on equine piroplasmosis: Epidemiology, vector ecology, risk factors, host immunity, diagnosis and control. Int. J. Environ. Res. Public. Health 2019, 16, 1736. [Google Scholar] [CrossRef]
- Stuen, S.; Granquist, E.G.; Silaghi, C. Anaplasma phagocytophilum—A widespread multi-host pathogen with highly adaptive strategies. Front. Cell. Infect. Microbiol. 2013, 3, 31. [Google Scholar] [CrossRef]
- Divers, T.J.; Gardner, R.B.; Madigan, J.E.; Witonsky, S.G.; Bertone, J.J.; Swinebroad, E.L.; Schutzer, S.E.; Johnson, A.L. Borrelia burgdorferi infection and Lyme disease in North American horses: A consensus statement: Lyme disease in horses. J. Vet. Intern. Med. 2018, 32, 617–632. [Google Scholar] [CrossRef]
- Freese, S.; Sheats, M.K. A suspected case of rocky mountain spotted fever in an adult horse in the southeastern United States. J. Equine Vet. Sci. 2019, 78, 134–137. [Google Scholar] [CrossRef] [PubMed]
- Marenzoni, M.L.; Stefanetti, V.; Papa, P.; Casagrande Proietti, P.; Bietta, A.; Coletti, M.; Passamonti, F.; Henning, K. Is the horse a reservoir or an indicator of Coxiella burnetii infection? Systematic review and biomolecular investigation. Vet. Microbiol. 2013, 167, 662–669. [Google Scholar] [CrossRef] [PubMed]
- Tirosh-Levy, S.; Gottlieb, Y.; Fry, L.M.; Knowles, D.P.; Steinman, A. Twenty years of equine piroplasmosis research: Global distribution, molecular diagnosis, and phylogeny. Pathogens 2020, 9, 926. [Google Scholar] [CrossRef]
- Scoles, G.A.; Ueti, M.W. Vector ecology of equine piroplasmosis. Annu. Rev. Entomol. 2015, 60, 561–580. [Google Scholar] [CrossRef]
- Wise, L.N.; Kappmeyer, L.S.; Mealey, R.H.; Knowles, D.P. Review of equine piroplasmosis. J. Vet. Intern. Med. 2013, 27, 1334–1346. [Google Scholar] [CrossRef]
- Rothschild, C.M. Equine piroplasmosis. J. Equine Vet. Sci. 2013, 33, 497–508. [Google Scholar] [CrossRef]
- Sears, K.P.; Kappmeyer, L.S.; Wise, L.N.; Silva, M.; Ueti, M.W.; White, S.; Reif, K.E.; Knowles, D.P. Infection dynamics of Theileria equi and Theileria haneyi, a newly discovered apicomplexan of the horse. Vet. Parasitol. 2019, 271, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Friedhoff, K.T.; Tenter, A.M.; Müller, I. Haemoparasites of equines: Impact on international trade of horses. Rev. Sci. Tech. Int. Off. Epizoot. 1990, 9, 1187–1194. [Google Scholar]
- Franzén, P.; Berg, A.-L.; Aspan, A.; Gunnarsson, A.; Pringle, J.D. Death of a horse infected experimentally with Anaplasma phagocytophilum. Vet. Rec. 2007, 160, 122–125. [Google Scholar] [CrossRef]
- Dzięgiel, B.; Adaszek, Ł.; Kalinowski, M.; Winiarczyk, S. Equine granulocytic anaplasmosis. Res. Vet. Sci. 2013, 95, 316–320. [Google Scholar] [CrossRef] [PubMed]
- Saleem, S.; Ijaz, M.; Farooqi, S.H.; Ghaffar, A.; Ali, A.; Iqbal, K.; Mehmood, K.; Zhang, H. Equine granulocytic anaplasmosis 28 years later. Microb. Pathog. 2018, 119, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Eisen, L. Vector competence studies with hard ticks and Borrelia burgdorferi sensu lato spirochetes: A review. Ticks Tick-Borne Dis. 2020, 11, 101359. [Google Scholar] [CrossRef] [PubMed]
- Basile, R.C.; Yoshinari, N.H.; Mantovani, E.; Bonoldi, V.N.; da Macoris, D.G.; de Queiroz-Neto, A. Brazilian borreliosis with special emphasis on humans and horses. Braz. J. Microbiol. 2017, 48, 167–172. [Google Scholar] [CrossRef]
- Kugeler, K.J.; Farley, G.M.; Forrester, J.D.; Mead, P.S. Geographic distribution and expansion of human Lyme disease, United States. Emerg. Infect. Dis. 2015, 21, 1455–1457. [Google Scholar] [CrossRef]
- Swinebroad, E.L. Borreliosis in sport horse practice. Vet. Clin. N. Am. Equine Pract. 2018, 34, 313–343. [Google Scholar] [CrossRef]
- Masuzawa, T. Terrestrial distribution of the Lyme borreliosis agent Borrelia burgdorferi sensu lato in East Asia. Jpn. J. Infect. Dis. 2004, 57, 229–235. [Google Scholar] [PubMed]
- Trinachartvanit, W.; Hirunkanokpun, S.; Sudsangiem, R.; Lijuan, W.; Boonkusol, D.; Baimai, V.; Ahantarig, A. Borrelia sp. phylogenetically different from Lyme disease- and relapsing fever-related Borrelia spp. in Amblyomma varanense from Python reticulatus. Parasit. Vectors 2016, 9, 359. [Google Scholar] [CrossRef] [PubMed]
- Takhampunya, R.; Thaloengsok, S.; Tippayachai, B.; Promsathaporn, S.; Leepitakrat, S.; Gross, K.; Davidson, S.A. Retrospective survey of Borrelia spp. From rodents and ticks in Thailand. J. Med. Entomol. 2021, 58, 1331–1344. [Google Scholar] [CrossRef]
- Lau, A.C.C.; Qiu, Y.; Moustafa, M.A.M.; Nakao, R.; Shimozuru, M.; Onuma, M.; Mohd-Azlan, J.; Tsubota, T. Detection of Borrelia burgdorferi sensu lato and relapsing fever Borrelia in feeding Ixodes ticks and rodents in Sarawak, Malaysia: New geographical records of Borrelia yangtzensis and Borrelia miyamotoi. Pathogens 2020, 9, 846. [Google Scholar] [CrossRef]
- Khoo, J.J.; Ishak, S.N.; Lim, F.S.; Mohd-Taib, F.S.; Khor, C.S.; Loong, S.K.; AbuBakar, S. Detection of a Borrelia sp. from Ixodes granulatus ticks collected from rodents in Malaysia. J. Med. Entomol. 2018, 55, 1642–1647. [Google Scholar] [CrossRef]
- Tyrrell, J.D.; Qurollo, B.A.; Tornquist, S.J.; Schlaich, K.G.; Kelsey, J.; Chandrashekar, R.; Breitschwerdt, E.B. Molecular identification of vector-borne organisms in Ehrlichia seropositive Nicaraguan horses and first report of Rickettsia felis infection in the horse. Acta Trop. 2019, 200, 105170. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, Y.; Adjou Moumouni, P.F.; Lee, S.-H.; Galon, E.M.; Tumwebaze, M.A.; Yang, H.; Huercha; Liu, M.; Guo, H.; et al. First description of Coxiella burnetii and Rickettsia spp. infection and molecular detection of piroplasma co-infecting horses in Xinjiang Uygur Autonomous Region, China. Parasitol. Int. 2020, 76, 102028. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.-C.; Hwang, S.; Kim, E.-M.; Park, Y.-J.; Shin, S.-U.; Jang, D.-H.; Chae, J.-S.; Choi, K.-S. Prevalence and molecular characterization of Coxiella burnetii in cattle, goats, and horses in the Republic of Korea. Vector Borne Zoonotic Dis. 2021, 21, 502–508. [Google Scholar] [CrossRef] [PubMed]
- Khademi, P.; Ownagh, A.; Ataei, B.; Kazemnia, A.; Eydi, J.; Khalili, M.; Mahzounieh, M.; Mardani, K. Molecular detection of Coxiella burnetii in horse sera in Iran. Comp. Immunol. Microbiol. Infect. Dis. 2020, 72, 101521. [Google Scholar] [CrossRef] [PubMed]
- Philippine Statistics Authority. Number of Livestock Holdings/Farms: Philippines. OpenSTAT Database. Available online: https://openstat.psa.gov.ph/Metadata/Agriculture-Forestry-Fisheries/Livestock-and-Poultry (accessed on 10 July 2021).
- The Manila Times. Philippine Horse-Racing Returns on July. The Manila Times: Metro Manila, Philippines, 12 June 2020. Available online: https://www.manilatimes.net/2020/06/12/sports/philippine-horse-racing-returns-in-july/731149 (accessed on 1 June 2021).
- Cruz-Flores, M.J.; Bata, M.; Co, B.; Claveria, F.G.; Verdida, R.; Xuan, X.; Igarashi, I. Immunochromatographic assay of Babesia caballi and Babesia equi Laveran 1901 (Theileria equi Mehlhorn and Schein, 1998) (Phylum Apicomplexa) infection in Philippine horses correlated with parasite detection in blood smears. Vet. Arkhiv 2010, 80, 715–722. [Google Scholar]
- Ybañez, A.P.; Ybañez, R.H.D.; Talle, M.G.; Arreglo, R.M.T.; Geens, M.J.C.; Villas, J.G.I.; Villar, S.R.; Laruga, C.L.; Cao, S.; Adjou Moumouni, F.P.; et al. Serological and molecular detection of Theileria equi and Babesia caballi in Philippine horses. Ticks Tick Borne Dis. 2018, 9, 1125–1128. [Google Scholar] [CrossRef]
- Del Pino, L.E.B.; Roberto, N.; Vincenzo, V.; Francesca, I.; Antonella, C.; Luca, A.G.; Francesco, B.; Teresa, S.M. Babesia caballi and Theileria equi infections in horses in central-southern Italy: Sero-molecular survey and associated risk factors. Ticks Tick Borne Dis. 2016, 7, 462–469. [Google Scholar] [CrossRef]
- Zhao, S.; Wang, H.; Zhang, S.; Xie, S.; Li, H.; Zhang, X.; Jia, L. First report of genetic diversity and risk factor analysis of equine piroplasm infection in equids in Jilin, China. Parasit. Vectors 2020, 13, 459. [Google Scholar] [CrossRef]
- Moretti, A.; Mangili, V.; Salvatori, R.; Maresca, C.; Scoccia, E.; Torina, A.; Moretta, I.; Gabrielli, S.; Tampieri, M.P.; Pietrobelli, M. Prevalence and diagnosis of Babesia and Theileria infections in horses in Italy: A preliminary study. Vet. J. 2010, 184, 346–350. [Google Scholar] [CrossRef]
- Salim, B.O.M.; Hassan, S.M.; Bakheit, M.A.; Alhassan, A.; Igarashi, I.; Karanis, P.; Abdelrahman, M.B. Diagnosis of Babesia caballi and Theileria equi infections in horses in Sudan using ELISA and PCR. Parasitol. Res. 2008, 103, 1145. [Google Scholar] [CrossRef]
- Ueti, M.W.; Palmer, G.H.; Scoles, G.A.; Kappmeyer, L.S.; Knowles, D.P. Persistently infected horses are reservoirs for intrastadial tick-borne transmission of the apicomplexan parasite Babesia equi. Infect. Immun. 2008, 76, 3525–3529. [Google Scholar] [CrossRef]
- Hailat, N.Q.; Lafi, S.Q.; Al-Darraji, A.M.; Al-Ani, F.K. Equine babesiosis associated with strenuous exercise: Clinical and pathological studies in jordan. Vet. Parasitol. 1997, 69, 1–8. [Google Scholar] [CrossRef]
- Qablan, M.A.; Oborník, M.; Petrželková, K.J.; Sloboda, M.; Shudiefat, M.F.; Hořín, P.; Lukeš, J.; Modrý, D. Infections by Babesia caballi and Theileria equi in Jordanian equids: Epidemiology and genetic diversity. Parasitology 2013, 140, 1096–1103. [Google Scholar] [CrossRef] [PubMed]
- Grandi, G.; Molinari, G.; Tittarelli, M.; Sassera, D.; Kramer, L.H. Prevalence of Theileria equi and Babesia caballi infection in horses from Northern Italy. Vector-Borne Zoonotic Dis. 2011, 11, 955–956. [Google Scholar] [CrossRef] [PubMed]
- Rüegg, S.R.; Torgerson, P.; Deplazes, P.; Mathis, A. Age-dependent dynamics of Theileria equi and Babesia caballi infections in southwest Mongolia based on IFAT and/or PCR prevalence data from domestic horses and ticks. Parasitology 2007, 134, 939–947. [Google Scholar] [CrossRef] [PubMed]
- Kouam, M.K.; Kantzoura, V.; Masuoka, P.M.; Gajadhar, A.A.; Theodoropoulos, G. Genetic diversity of equine piroplasms in Greece with a note on speciation within Theileria genotypes (T. equi and T. equi-like). Infect. Genet. Evol. 2010, 10, 963–968. [Google Scholar] [CrossRef] [PubMed]
- Hao, L.; Yuan, D.; Li, S.; Jia, T.; Guo, L.; Hou, W.; Lu, Z.; Mo, X.; Yin, J.; Yang, A.; et al. Detection of Theileria spp. in ticks, sheep keds (Melophagus ovinus), and livestock in the eastern Tibetan Plateau, China. Parasitol. Res. 2020, 119, 2641–2648. [Google Scholar] [CrossRef]
- Seo, M.-G.; Yun, S.-H.; Choi, S.-K.; Cho, G.-J.; Park, Y.-S.; Cho, K.-H.; Kwon, O.-D.; Kwak, D. Molecular and phylogenetic analysis of equine piroplasms in the Republic of Korea. Res. Vet. Sci. 2013, 94, 579–583. [Google Scholar] [CrossRef]
- Ybañez, A.P.; Mingala, C.N.; Ybañez, R.H.D. Historical review and insights on the livestock tick-borne disease research of a developing country: The Philippine scenario. Parasitol. Int. 2018, 67, 262–266. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Oh, H.-S.; Park, S.-C.; Chun, J. Towards a taxonomic coherence between average nucleotide identity and 16S rRNA gene sequence similarity for species demarcation of prokaryotes. Int. J. Syst. Evol. Microbiol. 2014, 64, 346–351. [Google Scholar] [CrossRef] [PubMed]
- Ybañez, A.P.; Ybañez, R.H.D.; Claveria, F.G.; Cruz-Flores, M.J.; Xuenan, X.; Yokoyama, N.; Inokuma, H. High genetic diversity of Anaplasma marginale detected from Philippine cattle. J. Vet. Med. Sci. 2014, 76, 1009–1014. [Google Scholar] [CrossRef] [PubMed]
- Ybañez, A.P.; Sivakumar, T.; Ybañez, R.H.D.; Vincoy, M.R.B.; Tingson, J.A.; Perez, Z.O.; Gabotero, S.R.; Buchorno, L.P.; Inoue, N.; Matsumoto, K.; et al. Molecular survey of bovine vector-borne pathogens in Cebu, Philippines. Vet. Parasitol. 2013, 196, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Atif, F.A. Anaplasma marginale and Anaplasma phagocytophilum: Rickettsiales pathogens of veterinary and public health significance. Parasitol. Res. 2015, 114, 3941–3957. [Google Scholar] [CrossRef]
- Woldehiwet, Z. The natural history of Anaplasma phagocytophilum. Vet. Parasitol. 2010, 167, 108–122. [Google Scholar] [CrossRef] [PubMed]
- Zeman, P.; Jahn, P. An entropy-optimized multilocus approach for characterizing the strains of Anaplasma phagocytophilum infecting horses in the Czech Republic. J. Med. Microbiol. 2009, 58, 423–429. [Google Scholar] [CrossRef][Green Version]
- Bae, J.-M. Horses as a potential reservoir of Lyme borreliosis in Jeju-Do, Korea. J. Prev. Med. Pub. Health 2018, 51, 213–214. [Google Scholar] [CrossRef]
- Marcelis, L.; De Marneffe, P.; Chaidron, E.; Bigaignon, G.; Kageruka, P.; Goubau, P. Horse reservoir for Borrelia burgdorferi? Lancet 1987, 329, 977. [Google Scholar] [CrossRef]
- Galay, R.L.; Talactac, M.R.; Ambita-Salem, B.V.; Chu, D.M.M.; dela Costa, L.M.O.; Salangsang, C.M.A.; Caracas, D.K.B.; Generoso, F.H.; Babelonia, J.A.; Vergano, J.L.; et al. Molecular detection of Rickettsia spp. and Coxiella burnetii in cattle, water buffalo, and Rhipicephalus (Boophilus) microplus ticks in Luzon island of the Philippines. Trop. Med. Infect. Dis. 2020, 5, 54. [Google Scholar] [CrossRef]
- Cardona, F.C. Serological detection of Q–Fever in the sera of humans and ruminants in selected areas in the Philippines. IAMURE Int. J. Ecol. Conserv. 2016, 18, 108–117. [Google Scholar] [CrossRef]
- Galay, R.L.; Manalo, A.A.L.; Dolores, S.L.D.; Aguilar, I.P.M.; Sandalo, K.A.C.; Cruz, K.B.; Divina, B.P.; Andoh, M.; Masatani, T.; Tanaka, T. Molecular detection of tick-borne pathogens in canine population and Rhipicephalus sanguineus (sensu lato) ticks from southern Metro Manila and Laguna, Philippines. Parasit. Vectors 2018, 11, 643. [Google Scholar] [CrossRef]
- Renvoisé, A.; Rolain, J.-M.; Socolovschi, C.; Raoult, D. Widespread use of real-time PCR for rickettsial diagnosis. FEMS Immunol. Med. Microbiol. 2012, 64, 126–129. [Google Scholar] [CrossRef]
- Alota, S.L.; Edquiban, T.R.J.; Galay, R.L.; Bernardo, J.M.G.; Sandalo, K.A.C.; Divina, B.P.; Tanaka, T. Determination of resistance status to amitraz in the cattle tick Rhipicephalus (Boophilus) microplus from Luzon, Philippines through bioassay and molecular analysis. Exp. Appl. Acarol. 2021, 83, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Goolsby, J.A.; Guerrero, F.D.; Gaskin, J.; Bendele, K.G.; Azhahianambi, P.; Amalin, D.; Cruz-Flores, M.J.; Kashefi, J.; Smith, L.; Racelis, A.; et al. Molecular comparison of cattle fever ticks from native and introduced ranges, with insights into optimal search areas for classical biological control agents. Southwest. Entomol. 2016, 41, 595–604. [Google Scholar] [CrossRef]
- Dugat, T.; Lagrée, A.-C.; Maillard, R.; Boulouis, H.-J.; Haddad, N. Opening the black box of Anaplasma phagocytophilum diversity: Current situation and future perspectives. Front. Cell. Infect. Microbiol. 2015, 5, 61. [Google Scholar] [CrossRef] [PubMed]
- Parrish, D. The Occurrence, Geographical Distribution and Wild Vertebrate Host Relationships of Ticks (Ixodoidea) on Luzon Island, Philippines, with Descriptions of Three New Species. Ph.D. Thesis, Oklahoma State University, Stillwater, OK, USA, 1971. [Google Scholar]
- Philippine Atmospheric, Geophysical and Astronomical Services Administration (PAGASA). Climate of the Philippines. Available online: https://www.pagasa.dost.gov.ph/information/climate-philippines (accessed on 28 September 2021).
- Alhassan, A.; Pumidonming, W.; Okamura, M.; Hirata, H.; Battsetseg, B.; Fujisaki, K.; Yokoyama, N.; Igarashi, I. Development of a single-round and multiplex PCR method for the simultaneous detection of Babesia caballi and Babesia equi in horse blood. Vet. Parasitol. 2005, 129, 43–49. [Google Scholar] [CrossRef]
- Barlough, J.E.; Madigan, J.E.; DeRock, E.; Bigornia, L. Nested polymerase chain reaction for detection of Ehrlichia equi genomic DNA in horses and ticks (Ixodes pacificus). Vet. Parasitol. 1996, 63, 319–329. [Google Scholar] [CrossRef]
- Simser, J.A.; Palmer, A.T.; Fingerle, V.; Wilske, B.; Kurtti, T.J.; Munderloh, U.G. Rickettsia monacensis sp. nov., a spotted fever group rickettsia, from ticks (Ixodes ricinus) collected in a European city park. Appl. Environ. Microbiol. 2002, 68, 4559–4566. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Liu, M.; Adjou Moumouni, P.F.; Liu, X.; Efstratiou, A.; Liu, Z.; Liu, Y.; Tao, H.; Guo, H.; Wang, G.; et al. First molecular detection of tick-borne pathogens in dogs from Jiangxi, China. J. Vet. Med. Sci. 2017, 79, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Postic, D.; Assous, M.V.; Grimont, P.A.D.; Baranton, G. Diversity of Borrelia burgdorfeii sensu lato evidenced by restriction fragment length polymorphism of rrf (5S)-rrl (23S) intergenic spacer amplicons. Int. J. Syst. Bacteriol. 1994, 44, 743–752. [Google Scholar] [CrossRef] [PubMed]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: http://www.R-project.org/ (accessed on 1 June 2021).
- Aragon, T. Epitools: Epidemiology Tools; R Package Version 0.5-10.1. 2020. Available online: https://CRAN.R-project.org/package=epitools (accessed on 1 June 2021).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | n | Babesia caballi | Anaplasma phagocytophilum | Borrelia burgdorferi Sensu Lato | Theileria equi (%) | Anaplasma marginale (%) | Coxiella burnetii (%) | Rickettsia spp. (%) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No. of Positives (%) | OR | 95% CI | p Value | No. of Positives (%) | OR | 95% CI | p Value | No. of Positives (%) | OR | 95% CI | p Value | ||||||
| Age Group | |||||||||||||||||
| Yearling (≤2 years) | 16 | 4 (25.00) | Ref. | 0.17 # | 3 (18.75) | Ref. | 0.44 | 6 (37.50) | Ref. | 0.96 | n.d. | n.d. | n.d. | n.d. | |||
| Colt/Filly (<4 years) | 22 | 3 (13.64) | 0.48 | 0.06–3.41 | 1 (4.55) | 0.22 | 0.0038–3.00 | 9 (40.91) | 1.15 | 0.26–5.37 | n.d. | n.d. | 1 (4.55) | n.d. | |||
| Stallion/Mare (≥4 years) | 86 | 8 (9.30) | 0.31 | 0.07–1.64 | 9 (10.47) | 0.51 | 0.11–3.31 | 33 (38.37) | 1.04 | 0.31–3.82 | 1 (1.16) | 1 (1.16) | n.d. | n.d. | |||
| Gender | n.d. | ||||||||||||||||
| Male | 54 | 2 (3.70) | Ref. | 0.013 #,* | 5 (9.26) | Ref. | 0.77 | 19 (35.19) | Ref. | 0.58 | 1 (1.85) | 1 (1.85) | 1 (1.85) | n.d. | |||
| Female | 70 | 13 (18.57) | 5.86 | 1.24–55.90 | 8 (11.43) | 1.26 | 0.34–5.23 | 29 (41.43) | 1.30 | 0.59–2.91 | n.d. | n.d. | n.d. | n.d. | |||
| Total | 124 | 15 (12.10) | 13 (10.48) | 48 (38.71) | 1 (0.81) | 1 (0.81) | 1 (0.81) | n.d. | |||||||||
| Pathogen | Number of Positives | % |
|---|---|---|
| Single infection | 42 | 33.87 |
| Babesia caballi | 5 | 4.03 |
| Theileria equi | 1 | 0.81 |
| Anaplasma phagocytophilum | 4 | 3.23 |
| Anaplasma marginale | 1 | 0.81 |
| Borreliaburgdorferi sensu lato (Bbsl) | 31 | 25 |
| Multiple infections | 18 | 14.52 |
| Bbsl and B. caballi | 9 | 7.26 |
| Bbsl and A. phagocytophilum | 7 | 5.65 |
| B. caballi and A. phagocytophilum | 1 | 0.81 |
| Bbsl, A. phagocytophilum, and C. burnetii | 1 | 0.81 |
| Total number of of positive samples | 60 | 48.39 |
| Coinfecting Pathogens | No. of Positives (%) | OR | 95% CI | p Value |
|---|---|---|---|---|
| Bbsl and B. caballi | 9 (7.26) | 2.67 | 0.78–9.84 | 0.09 # |
| Bbsl and A. phagocytophilum | 8 (6.45) | 2.81 | 0.75–11.71 | 0.13 # |
| B. caballi and A. phagocytophilum | 1 (0.81) | 0.58 | 0.013–4.53 | 0.61 |
| Pathogen | Variable | Category | β | SE | p Value | OR | 95% CI | Final Model AIC |
|---|---|---|---|---|---|---|---|---|
| B. caballi | Gender | Male | Ref. | 87.54 | ||||
| Female | 1.75 | 0.79 | 0.026 * | 5.77 | 1.23–27.03 | |||
| Bbsl positivity | Negative | Ref. | ||||||
| Positive | 0.95 | 0.58 | 0.10 | 2.58 | 0.83–8.03 | |||
| A. phagocytophilum | Bbsl positivity | Negative | Ref. | 84.13 | ||||
| Positive | 1.04 | 0.6 | 0.08 | 2.84 | 0.87–9.27 | |||
| Borreliaburgdorferi sensu lato (Bbsl) | B. caballi positivity | Negative | Ref. | 164.88 | ||||
| Positive | 1.06 | 0.57 | 0.063 | 2.89 | 0.95–8.82 | |||
| A. phagocytophilum positivity | Negative | Ref. | ||||||
| Positive | 1.12 | 0.61 | 0.067 | 3.06 | 0.93–10.12 | |||
| Pathogen | Target Gene | PCR Assay Type | Primer Sequence (5′–3′) | Ta (°C) | Amplicon Size (bp) | Detection Threshold | Positive Control | Reference |
|---|---|---|---|---|---|---|---|---|
| Equine piroplasma | 18S rRNA | multiplex-nested | GTTGATCCTGCCAGTAGTCA | 54 | 913/867 | B. caballi- and T. equi-positive horse gDNA | [62] | |
| CGGTATCTGATCGTCTTCGA | ||||||||
| TCGAAGACGATCAGATACCGTCG | 54 | |||||||
| Babesia caballi | CTCGTTCATGATTTAGAATTGCT | 540 | 0.18 parasite cells | |||||
| Theileria equi/Theileria haneyi | TGCCTTAAACTTCCTTGCGAT | 392 | 0.018 parasite cells | |||||
| Anaplasma phagocytophilum | 16S rRNA | nested | TCCTGGCTCAGAACGAACGCTGGCGGC | 50 | 1433 | 3 infected neutrophils | A. phagocytophilum-positive cattle gDNA | [63] |
| AGTCACTGACCCAACCTTAAATGGCTG | ||||||||
| GTCGAACGGATTATTCTTTATAGCTTGC | 50 | 928 | ||||||
| CCCTTCCGTTAAGAAGGATCTAATCTCC | ||||||||
| Anaplasma marginale | groEL | nested-touchdown | TCCTGGCTCAGAACGAACGCTGGCGGC | 74–65 | 866 | 2 gene copies | A. marginale-positive cattle gDNA | [46] |
| AGTCACTGACCCAACCTTAAATGGCTG | ||||||||
| GTCGAACGGATTATTCTTTATAGCTTGC | 74–68 | 618–768 | ||||||
| CCCTTCCGTTAAGAAGGATCTAATCTCC | ||||||||
| Rickettsia spp. | 16S rRNA | single | AACGTCATTATCTTCCTTGC | 59 | 426 | n.a. | R. slovaca-positive horse gDNA | [64] |
| AGAGTTTGATCCTGGCTCAG | ||||||||
| Coxiella burnetii | 16S rRNA | touchdown | ATTGAAGAGTTTGATTCTGG | 58–48 | ~1450 | n.a. | C. burnetii-positive horse gDNA | [65] |
| CGGCTTCCCGAAGGTTAG | ||||||||
| Borrelia burgdorferi sensu lato | rrf-rrl (5–23S rRNA intergenic spacer) | nested | CGACCTTCTTCGCCTTAAAGC | 57.6 | 412 | n.a. | B. garinii-positive tick gDNA | [66] |
| TAAGCTGACTAATACTAATTACCC | ||||||||
| CTGCGAGTTCGCGGGAGA | 55 | 226–266 | ||||||
| TCCTAGGCATTCACCATA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galon, E.M.; Macalanda, A.M.; Garcia, M.M.; Ibasco, C.J.; Garvida, A.; Ji, S.; Zafar, I.; Hasegawa, Y.; Liu, M.; Ybañez, R.H.; et al. Molecular Identification of Selected Tick-Borne Protozoan and Bacterial Pathogens in Thoroughbred Racehorses in Cavite, Philippines. Pathogens 2021, 10, 1318. https://doi.org/10.3390/pathogens10101318
Galon EM, Macalanda AM, Garcia MM, Ibasco CJ, Garvida A, Ji S, Zafar I, Hasegawa Y, Liu M, Ybañez RH, et al. Molecular Identification of Selected Tick-Borne Protozoan and Bacterial Pathogens in Thoroughbred Racehorses in Cavite, Philippines. Pathogens. 2021; 10(10):1318. https://doi.org/10.3390/pathogens10101318
Chicago/Turabian StyleGalon, Eloiza May, Adrian Miki Macalanda, Mary Margarett Garcia, Chrysler James Ibasco, Anatolio Garvida, Shengwei Ji, Iqra Zafar, Yae Hasegawa, Mingming Liu, Rochelle Haidee Ybañez, and et al. 2021. "Molecular Identification of Selected Tick-Borne Protozoan and Bacterial Pathogens in Thoroughbred Racehorses in Cavite, Philippines" Pathogens 10, no. 10: 1318. https://doi.org/10.3390/pathogens10101318
APA StyleGalon, E. M., Macalanda, A. M., Garcia, M. M., Ibasco, C. J., Garvida, A., Ji, S., Zafar, I., Hasegawa, Y., Liu, M., Ybañez, R. H., Umemiya-Shirafuji, R., Ybañez, A., Claveria, F., & Xuan, X. (2021). Molecular Identification of Selected Tick-Borne Protozoan and Bacterial Pathogens in Thoroughbred Racehorses in Cavite, Philippines. Pathogens, 10(10), 1318. https://doi.org/10.3390/pathogens10101318