Distribution and Genetic Diversity of Aedes aegypti Subspecies across the Sahelian Belt in Sudan

 ,
,  ,
,  , ,
, , 
Abstract
1. Introduction
2. Results
2.1. Morphological and Molecular Identification of Ae. aegypti at Eight Sites of Sudan
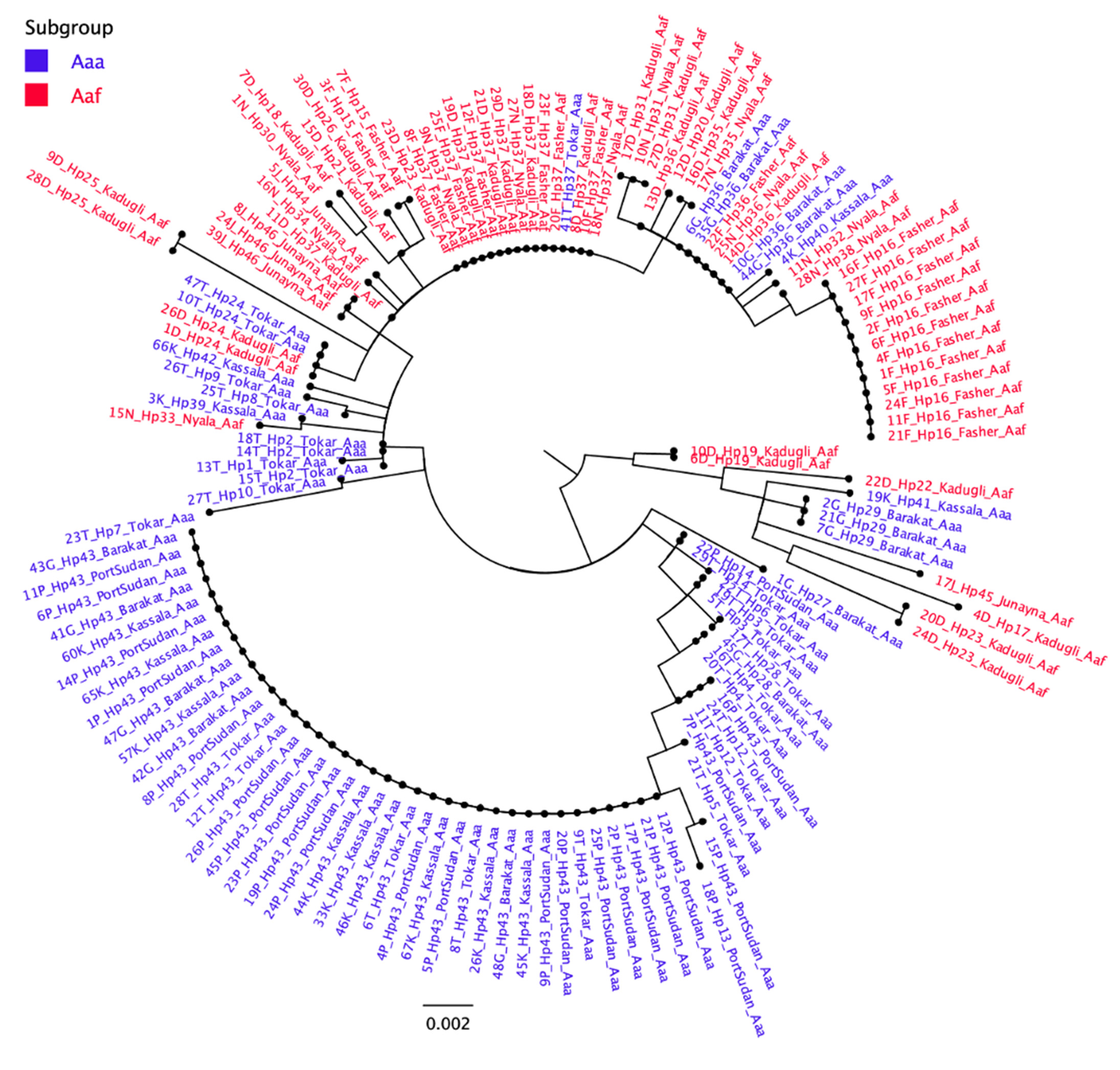
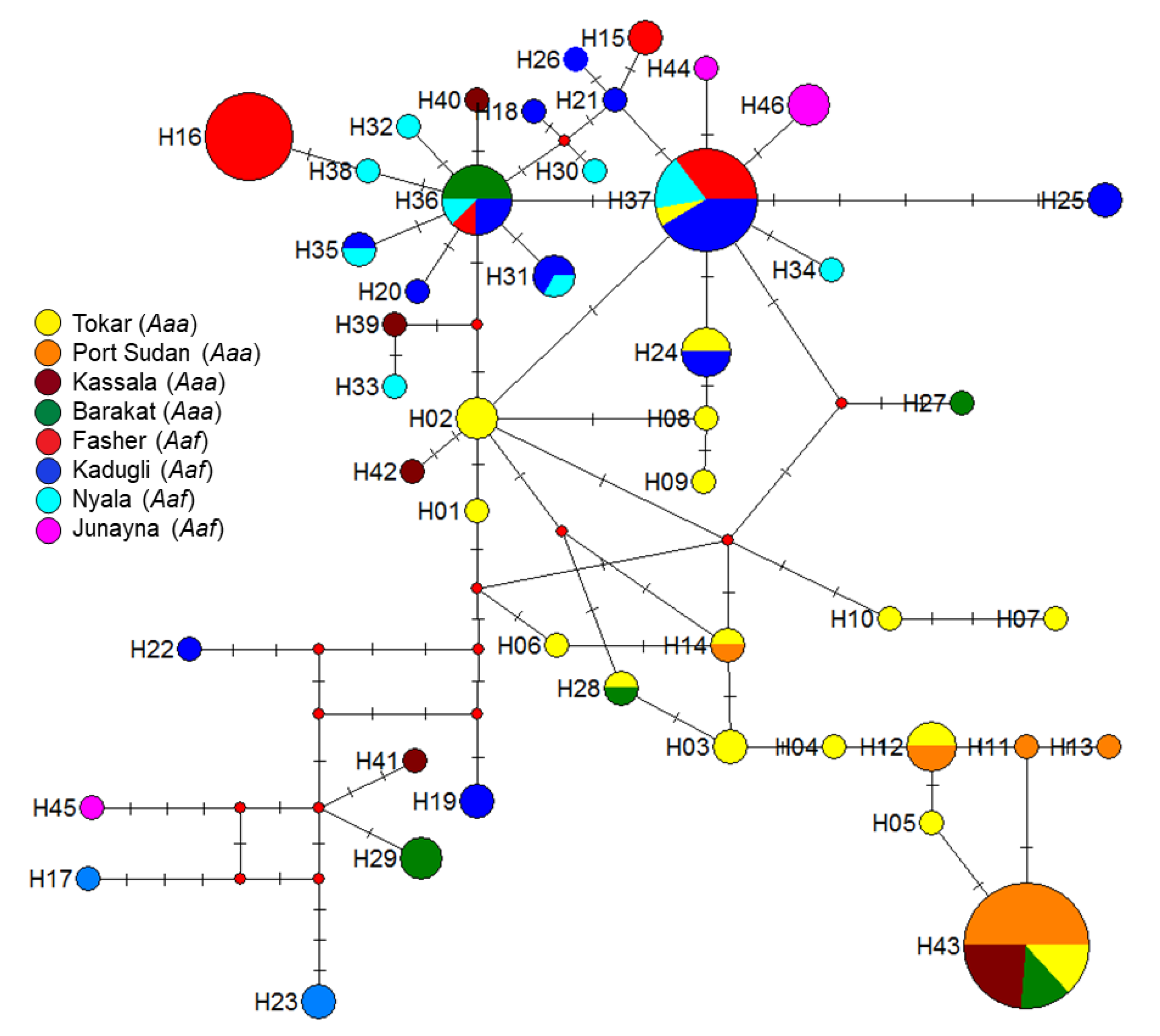
2.2. Mitochondrial Haplotype Analysis
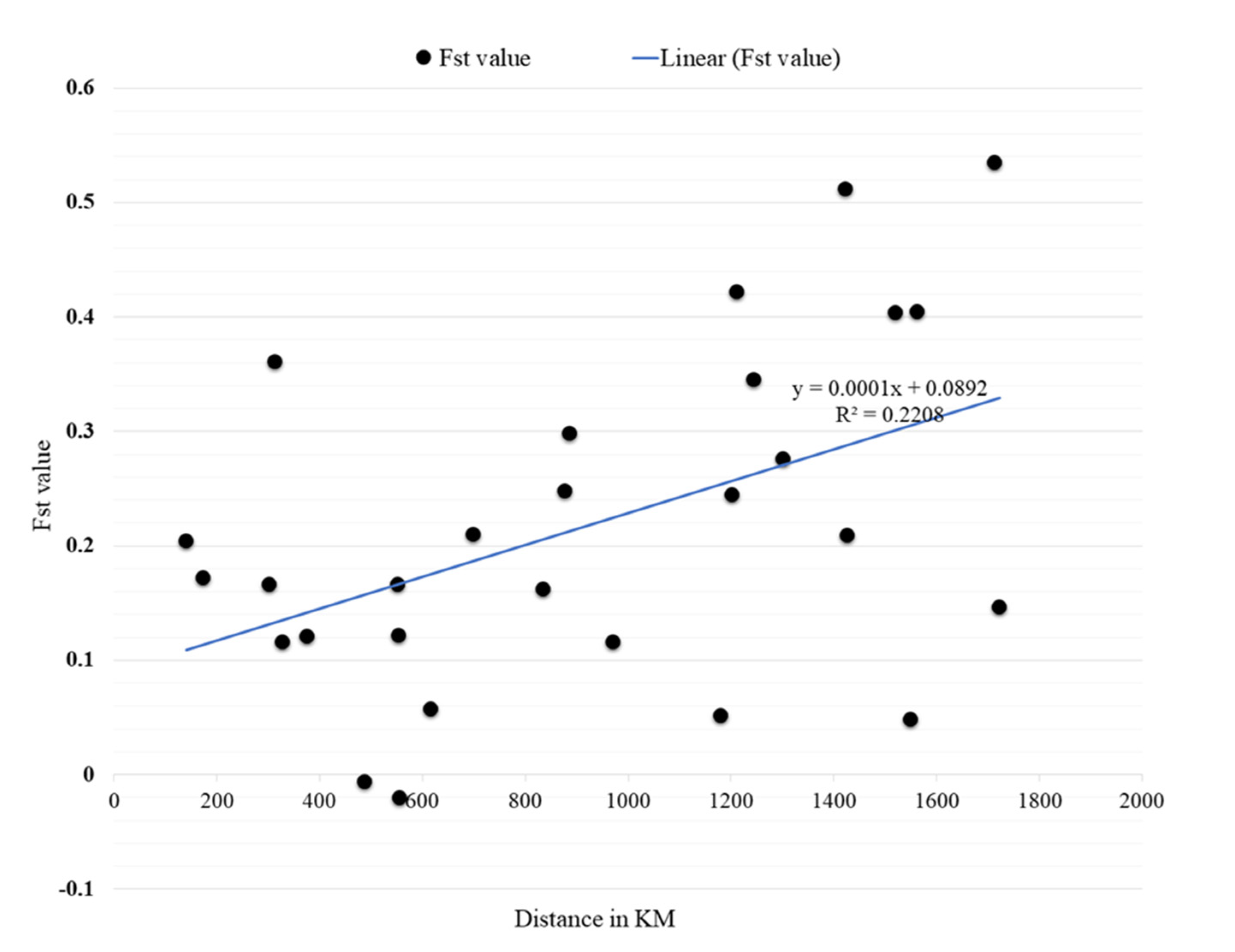
2.3. Genetic Variance (Pairwise FST) and Mantel Test Results
2.4. Genetic Structure of Aedes aegypti in Different Study Sites
2.5. Test of Neutrality and Natural Selection in Different Populations of Ae. aegypti
3. Discussion
4. Methods
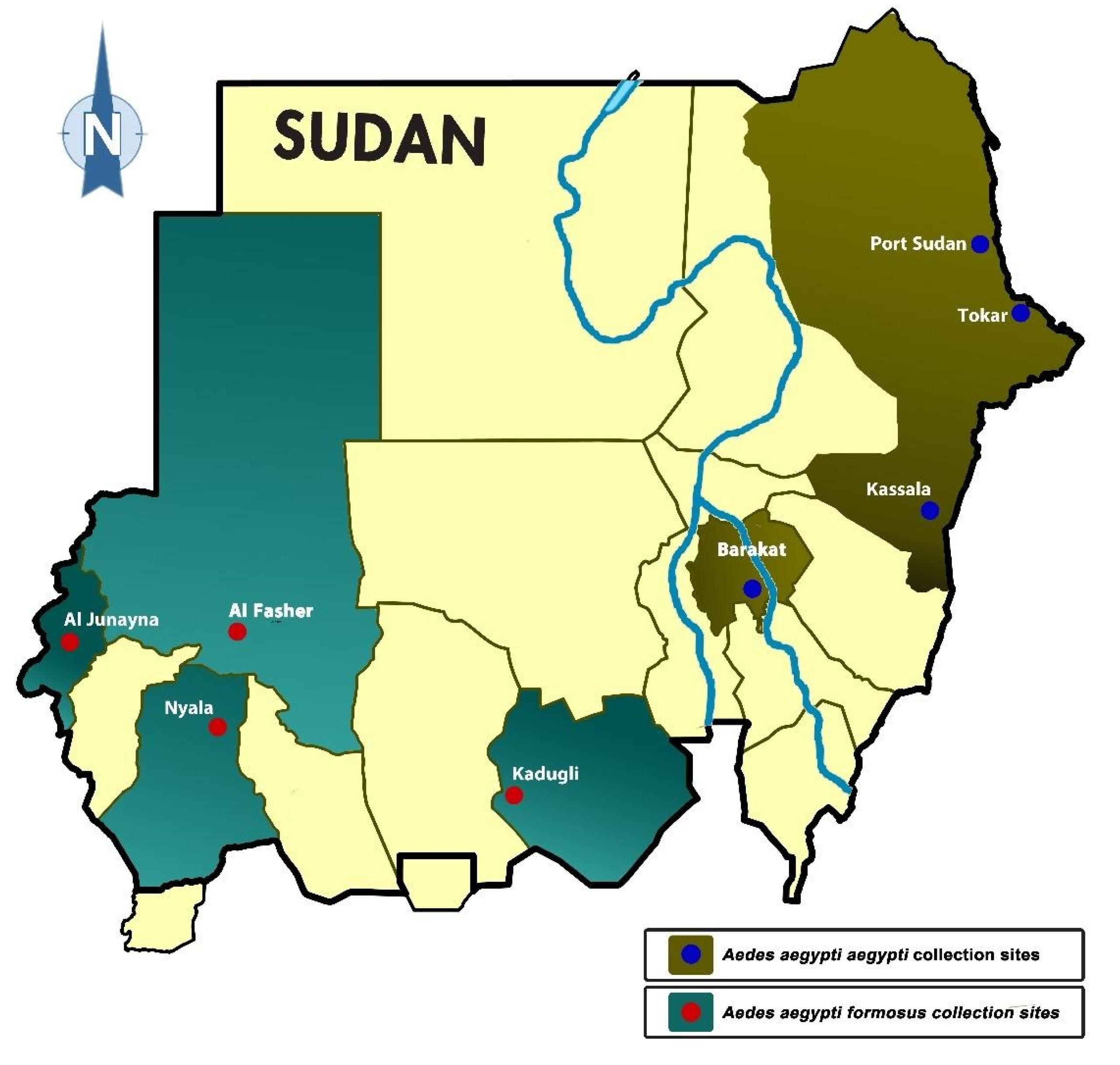
4.1. Study Sites
4.2. Mosquito Collection, Rearing, and Morphological Identification
4.3. DNA Extraction and Molecular Identification
4.4. PCR Amplification for Mitochondrial Marker
4.5. Mitochondrial Sequences Analyses
4.6. Phylogenetic Analyses/Trees
4.7. Genetic Diversity and Genetic Structure of Aedes aegypti Subspecies
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kraemer, M.U.; E Sinka, M.; A Duda, K.; Mylne, A.Q.N.; Shearer, F.M.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; E Coelho, G.; Van Bortel, W.; et al. The global distribution of the arbovirus vectors Aedes aegypti and Ae. albopictus. eLife 2015, 4, e08347. [Google Scholar] [CrossRef] [PubMed]
- Souza-Neto, J.A.; Powell, J.R.; Bonizzoni, M. Aedes aegypti vector competence studies: A review. Infect Genet Evol. 2019, 67, 191–209. [Google Scholar] [CrossRef] [PubMed]
- Weetman, D.; Kamgang, B.; Badolo, A.; Moyes, C.L.; Shearer, F.M.; Coulibaly, M.B.; Pinto, J.; Lambrechts, L.; McCall, P. Aedes Mosquitoes and Aedes-Borne Arboviruses in Africa: Current and Future Threats. Int. J. Environ. Res. Public Health 2018, 15, 220. [Google Scholar] [CrossRef] [PubMed]
- Kamal, M.; Kenawy, M.A.; Rady, M.H.; Khaled, A.S.; Samy, A.M. Mapping the global potential distributions of two arboviral vectors Aedes aegypti and Ae. albopictus under changing climate. PLoS ONE 2018, 13, e0210122. [Google Scholar] [CrossRef] [PubMed]
- Kweka, E.J.; Baraka, V.; Mathias, L.; Mwang’onde, B.; Baraka, G.; Lyaruu, L.; Mahande, A.M. Ecology of Aedes Mosquitoes, the Major Vectors of Arboviruses in Human Population. In Dengue Fever—A Resilient Threat in the Face of Innovation; Abelardo Falcón-Lezama, J., Betancourt-Cravioto, M., Tapia-Conyer, R., Eds.; IntechOpen: London, UK, 2019; Available online: https://www.intechopen.com/books/dengue-fever-a-resilient-threat-in-the-face-of-innovation/ecology-of-aedes-mosquitoes-the-major-vectors-of-arboviruses-in-human-population (accessed on 9 November 2020).
- Morrison, A.C.; Zielinski-Gutierrez, E.; Scott, T.W.; Rosenberg, R. Defining Challenges and Proposing Solutions for Control of the Virus Vector Aedes aegypti. PLoS Med. 2008, 5, e68. [Google Scholar] [CrossRef]
- Scott, T.W.; Takken, W. Feeding strategies of anthropophilic mosquitoes result in increased risk of pathogen transmission. Trends Parasitol. 2012, 28, 114–121. [Google Scholar] [CrossRef]
- Mattingly, P.F. Genetical Aspects of the Aëdes Aegypti Problem: I.—Taxonomy and Bionomics. Ann. Trop. Med. Parasitol. 1957, 51, 392–408. [Google Scholar] [CrossRef]
- McBride, C.S.; Baier, F.; Omondi, A.B.; Spitzer, S.A.; Lutomiah, J.; Sang, R.; Ignell, R.; Vosshall, L.B. Evolution of mosquito preference for humans linked to an odorant receptor. Nat. Cell Biol. 2014, 515, 222–227. [Google Scholar] [CrossRef]
- Lewis, D.J. The Aëdes Mosquitoes of the Sudan. Ann. Trop. Med. Parasitol. 1955, 49, 164–173. [Google Scholar] [CrossRef]
- Lewis, D.J. Observations on the distribution and taxonomy of culicidae (diptera) in the sudan. Trans. R. Entomol. Soc. Lond. 2009, 95, 1–24. [Google Scholar] [CrossRef]
- Paupy, C.; Brengues, C.; Ndiath, O.; Toty, C.; Hervé, J.-P.; Simard, F. Morphological and genetic variability within Aedes aegypti in Niakhar, Senegal. Infect. Genet. Evol. 2010, 10, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.E.; McBride, C.S.; Johnson, P.; Ritchie, S.; Paupy, C.; Bossin, H.; Lutomiah, J.; Fernandez-Salas, I.; Ponlawat, A.; Cornel, A.J.; et al. Worldwide patterns of genetic differentiation imply multiple ‘domestications’ of Aedes aegypti, a major vector of human diseases. Proc. R. Soc. B Boil. Sci. 2011, 278, 2446–2454. [Google Scholar] [CrossRef] [PubMed]
- Gloria-Soria, A.; Ayala, D.; Bheecarry, A.; Calderon-Arguedas, O.; Chadee, D.D.; Chiappero, M.; Coetzee, M.; Bin Elahee, K.; Fernandez-Salas, I.; Kamal, H.A.; et al. Global genetic diversity ofAedes aegypti. Mol. Ecol. 2016, 25, 5377–5395. [Google Scholar] [CrossRef] [PubMed]
- Captain-Esoah, M.; Baidoo, P.K.; Frempong, K.K.; Adabie-Gomez, D.; Chabi, J.; Obuobi, D.; Amlalo, G.K.; Veriegh, F.B.; Donkor, M.; Asoala, V.; et al. Biting Behavior and Molecular Identification of Aedes aegypti (Diptera: Culicidae) Subspecies in Some Selected Recent Yellow Fever Outbreak Communities in Northern Ghana. J. Med. Entomol. 2020, 57, 1239–1245. [Google Scholar] [CrossRef]
- Black, W.C.; Bennett, K.E.; Gorrochótegui-Escalante, N.; Barillas-Mury, C.V.; Fernández-Salas, I.; de Lourdes Muñoz, M.; Farfán-Alé, J.A.; Olson, K.E.; Beaty, B.J. Flavivirus Susceptibility in Aedes aegypti. Arch. Med. Res. 2002, 33, 379–388. [Google Scholar] [CrossRef]
- Dickson, L.B.; Sanchez-Vargas, I.; Sylla, M.; Fleming, K.; Black, W.C. Vector Competence in West African Aedes aegypti Is Flavivirus Species and Genotype Dependent. PLoS Negl. Trop. Dis. 2014, 8, e3153. [Google Scholar] [CrossRef]
- Powell, J.R.; Tabachnick, W.J. History of domestication and spread of Aedes aegypti—A Review. Memórias Inst. Oswaldo Cruz 2013, 108 (Suppl. 1), 11–17. [Google Scholar] [CrossRef]
- Tabachnick, W.J.; Powell, J.R. A world-wide survey of genetic variation in the yellow fever mosquito, Aedes aegypti. Genet. Res. 1979, 34, 215–229. [Google Scholar] [CrossRef]
- Rašić, G.; Schama, R.; Powell, R.; Maciel-de Freitas, R.; Endersby-Harshman, N.M.; Filipović, I.; Sylvestre, G.; Máspero, R.C.; Hoffmann, A.A. Contrasting genetic structure between mitochondrial and nuclear markers in the dengue fever mosquito from Rio de Janeiro: Implications for vector control. Evol. Appl. 2015, 8, 901–915. [Google Scholar] [CrossRef]
- Sousa, A.A.D.; Fraga, E.; Sampaio, I.; Schneider, H.; Barros, M.C. Genetic differentiation in populations of Aedes aegypti (Diptera, Culicidae) dengue vector from the Brazilian state of Maranhão. Rev. Bras. Entomol. 2017, 61, 51–59. [Google Scholar] [CrossRef][Green Version]
- Crawford, J.E.; Alves, J.M.; Palmer, W.J.; Day, J.P.; Sylla, M.; Ramasamy, R.; Surendran, S.N.; Iv, W.C.B.; Pain, A.; Jiggins, F.M. Population genomics reveals that an anthropophilic population of Aedes aegypti mosquitoes in West Africa recently gave rise to American and Asian populations of this major disease vector. BMC Biol. 2017, 15, 16. [Google Scholar] [CrossRef] [PubMed]
- Joyce, A.L.; Torres, M.M.; Torres, R.; Moreno, M. Genetic variability of the Aedes aegypti (Diptera: Culicidae) mosquito in El Salvador, vector of dengue, yellow fever, chikungunya and Zika. Parasites Vectors 2018, 11, 637. [Google Scholar] [CrossRef]
- Naim, D.M.; Kamal, N.Z.M.; Mahboob, S. Population structure and genetic diversity of Aedes aegypti and Aedes albopictus in Penang as revealed by mitochondrial DNA cytochrome oxidase I. Saudi J. Biol. Sci. 2020, 27, 953–967. [Google Scholar] [CrossRef] [PubMed]
- Lewis, D.J. TheStegomyiaMosquitoes of the Anglo-Egyptian Sudan. Ann. Trop. Med. Parasitol. 1953, 47, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Lewis, D.J. Mosquitoes in Relation to Yellow Fever in the Nuba Mountains, Anglo-Egyptian Sudan. Ann. Trop. Med. Parasitol. 1943, 37, 65–76. [Google Scholar] [CrossRef]
- Ahmed, A.; Elduma, A.H.; Magboul, B.; Higazi, T.B.; Ali, Y. The First Outbreak of Dengue Fever in Greater Darfur, Western Sudan. Trop. Med. Infect. Dis. 2019, 4, 43. [Google Scholar] [CrossRef]
- Ahmed, R.M.H. Breeding and Resting Behaviour of Aedes aegypti in Indoor and Outdoor Environment in Kassala City, Sudan 2014/2015. Health Sci. J. 2019, 13, 10. [Google Scholar]
- Elnour, M.B.; Moustafa, M.A.M.; Khogali, R.; Azrag, R.S.; Alanazi, A.D.; Kheir, A.; Nakao, R.; De Meeûs, T.; Salim, B. Distinct haplotypes and free movement of Aedes aegypti in Port Sudan, Sudan. J. Appl. Entomol. 2020, 144, 817–823. [Google Scholar] [CrossRef]
- Ahmed, A.; Dietrich, I.; LaBeaud, A.D.; Lindsay, S.W.; Musa, A.M.; Weaver, S.C. Risks and Challenges of Arboviral Diseases in Sudan: The Urgent Need for Actions. Viruses 2020, 12, 81. [Google Scholar] [CrossRef]
- Himatt, S.; Osman, K.E.; Okoued, S.I.; Seidahmed, O.E.; Beatty, M.E.; Soghaier, M.A.; Elmusharaf, K. Sero-prevalence of dengue infections in the Kassala state in the eastern part of the Sudan in 2011. J. Infect. Public Health 2015, 8, 487–492. [Google Scholar] [CrossRef]
- Rm, A.; Sm, H.; Ah, E. Climatic Factors Affecting Density of Aedes aegypti (Diptera: Culicidae) in Kassala City, Sudan 2014/2015. Asploro J. Biomed. Clin. Case Rep. 2019, 2, 58–68. [Google Scholar] [CrossRef]
- Soghaier, M.A.; Hagar, A.; Abbas, M.A.; Elmangory, M.M.; Eltahir, K.M.; Sall, A.A. Yellow Fever outbreak in Darfur, Sudan in October 2012; the initial outbreak investigation report. J. Infect. Public Health 2013, 6, 370–376. [Google Scholar] [CrossRef] [PubMed]
- Seidahmed, O.M.E.; Siam, H.A.M.; Soghaier, M.A.; Abubakr, M.; Osman, H.A.; Elrhman, L.S.A.; Elmagbol, B.; Velayudhan, R. Dengue vector control and surveillance during a major outbreak in a coastal Red Sea area in Sudan. East Mediterr. Health J. 2012, 18, 8. [Google Scholar] [CrossRef]
- Soghaier, M.A.; Abdelgadir, D.M.; Abdelkhalig, S.M.; Kafi, H.; Zarroug, I.M.A.; A Sall, A.; Eldegai, M.H.; Elageb, R.M.; Osman, M.M.; Khogali, H. Evidence of pre-existing active Zika virus circulation in Sudan prior to 2012. BMC Res. Notes 2018, 11, 906. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A. Urgent call for a global enforcement of the public sharing of health emergencies data: Lesson learned from serious arboviral disease epidemics in Sudan. Int. Health 2020, 12, 238–240. [Google Scholar] [CrossRef]
- Gould, L.H.; Osman, M.S.; Farnon, E.C.; Griffith, K.S.; Godsey, M.S.; Karch, S.; Mulenda, B.; El Kholy, A.; Grandesso, F.; De Radiguès, X.; et al. An outbreak of yellow fever with concurrent chikungunya virus transmission in South Kordofan, Sudan, 2005. Trans. R. Soc. Trop. Med. Hyg. 2008, 102, 1247–1254. [Google Scholar] [CrossRef]
- Adam, A.; Seidahmed, O.M.; Weber, C.; Schnierle, B.; Schmidt-Chanasit, J.; Reiche, S.; Jassoy, C. Low Seroprevalence Indicates Vulnerability of Eastern and Central Sudan to Infection with Chikungunya Virus. Vector-Borne Zoonotic Dis. 2016, 16, 290–291. [Google Scholar] [CrossRef]
- Bushra, H.E.E.; Habtewold, B.W.; Gasseer, N.A.; Mohamed, R.E.; Mohamednour, S.A.; Abshar, M.; Al Magboul, B. Outbreak of Chikungunya Fever in Sudan, 2018–2019. JOJ Pub Health. 2019, 4, 555644. [Google Scholar]
- Bracco, J.E.; Capurro, M.L.; Lourenço-De-Oliveira, R.; Sallum, M.A.M. Genetic variability of Aedes aegypti in the Americas using a mitochondrial gene: Evidence of multiple introductions. Memórias Inst. Oswaldo Cruz 2007, 102, 573–580. [Google Scholar] [CrossRef]
- Ahmed, A.; Ali, Y.; Elmagboul, B.; Mohamed, O.; Elduma, A.; Bashab, H.; Mahamoud, A.; Khogali, H.; Elaagip, A.; Higazi, T. Dengue Fever in the Darfur Area, Western Sudan. Emerg. Infect Dis. 2019, 25, 2126. [Google Scholar] [CrossRef]
- Futami, K.; Iwashita, H.; Higa, Y.; A Lutiali, P.; O Sonye, G.; Mwatele, C.; Njenga, S.M.; Minakawa, N. Geographical Distribution of Aedes aegypti aegypti and Aedes aegypti formosus (Diptera: Culicidae) in Kenya and Environmental Factors Related to Their Relative Abundance. J. Med. Entomol. 2019, 57, 772–779. [Google Scholar] [CrossRef] [PubMed]
- Thomson, M.C.; Elnaiern, D.A.; Ashford, R.W.; Connor, S.J. Towards a kala azar risk map for Sudan: Mapping the potential distribution of Phlebotomus orientalis using digital data of environmental variables. Trop. Med. Int. Health 1999, 4, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Elnaiem, D.A.; Hassan, H.K.; Ward, R.D. Associations of Phlebotomus orientalis and other sandflies with vegetation types in the eastern Sudan focus of kala-azar. Med. Vet. Entomol. 1999, 13, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Powell, J.R.; Gloria-Soria, A.; Kotsakiozi, P. Recent History of Aedes aegypti: Vector Genomics and Epidemiology Records. BioScience 2018, 68, 854–860. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, G.H.E. Mosquito of the Ethiopian Region, 2nd ed.; British Museum (NATURAL HISTORY), CROMWELL ROAD, S.W.7; 1952; Available online: http://mosquito-taxonomic-inventory.info/sites/mosquito-taxonomic-inventory.info/files/Hopkins%201952.pdf (accessed on 24 February 2011).
- Huang, Y.-M. The subgenus Stegomyia of Aedes in the Afrotropical Region with keys to the species (Diptera: Culicidae). Zootaxa 2004, 700, 1. [Google Scholar] [CrossRef]
- Beebe, N.W.; Whelan, P.I.; Corcoran, S.; Cooper, R.D. A Polymerase Chain Reaction-Based Diagnostic to Identify Larvae and Eggs of Container Mosquito Species from the Australian Region. J. Med. Entomol. 2007, 44, 5. [Google Scholar] [CrossRef]
- Paupy, C.; Le Goff, G.; Brengues, C.; Guerra, M.; Revollo, J.; Simon, Z.B.; Hervé, J.-P.; Fontenille, D. Genetic structure and phylogeography of Aedes aegypti, the dengue and yellow-fever mosquito vector in Bolivia. Infect Genet Evol. 2012, 12, 1260–1269. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree. 2012. Available online: https://github.com/rambaut/figtree/releases (accessed on 26 November 2018).
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Bandelt, H.J.; Forster, P.; Rohl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Dupanloup, I.; Schneider, S.; Excoffier, L. A simulated annealing approach to define the genetic structure of populations: Defining the genetic structure of populations. Mol. Ecol. 2002, 11, 2571–2581. [Google Scholar] [CrossRef] [PubMed]
- Byron, V. Weckworth NJD Mark Hebblewhite, Marco Musiani. Genetic Analysis of Caribou Connectivity and the Spatial Effects of Wolf Predation on Caribou Survival in Banff and Jasper National Parks. Parks Canada (Banff and Jasper National Parks) Banff National Park. 2014. Available online: https://www.researchgate.net/publication/236651958. (accessed on 26 November 2018).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Site | Region | Subspecies | Coordinates | Collection Year | Larval Habitats | NO |
|---|---|---|---|---|---|---|---|
| P | Port Sudan | Coastal/Eastern | Aaa | 19.617°37′0″ N, 37.217°13′0″ E | 2014 | Clay pots, cement water reservoir, plastic barrels and jerrycans | 26 |
| T | Tokar | Eastern | Aaa | 18.425°25′31″N, 37.729°43′45″E | 2016 | Clay pots and jerrycans. | 22 |
| K | Kassala | Eastern | Aaa | 15.45°27′0″N, 36.4°24′0″E | 2014 | Flowerpots, cement reservoir, and clay pots. | 13 |
| G | Barakat/Gezira | Central | Aaa | 14.314°18′50.84″N, 33.534°32′3.74″E | 2014 | Plastic barrels, tyres, and bathtubs | 20 |
| D | Kadugli | South | Aaf | 11.017°1′0″N, 29.717°43′0″E | 2015 | Tyres, clay pots, and plastic barrels | 30 |
| N | Nyala | West | Aaf | 12.036°2′11″N, 24.878°52′37″E | 2016–2017 | Clay pots and jerrycans | 16 |
| F | Al Fasher | Northern West | Aaf | 13.631°37′50″N, 25.35°21′0″E | 2017 | Clay pots and jerrycans | 22 |
| J | Al Junaynah | West | Aaf | 13.45°27′0″N, 22.45°27′0″E | 2014 | Clay pots, cement barrels, and plastic containers | 8 |
| Site | Subspecies/Form | N | S | H | Hd | π | Tajima’s D | Fu’s FS |
|---|---|---|---|---|---|---|---|---|
| Port Sudan | Aaa/domestic | 24 | 6 | 5 | 0.377 | 0.002 | −1.319 | −1.142 |
| Tokar | Aaa/domestic | 25 | 14 | 16 | 0.947 | 0.007 | 0.592 | −6.792 |
| Kassala | Aaa/domestic | 13 | 17 | 5 | 0.539 | 0.008 | −0.606 | 2.254 |
| Barakat/Gezira | Aaa/domestic | 14 | 14 | 5 | 0.791 | 0.010 | 1.500 | 3.419 |
| Kadugli | Aaf/wild | 27 | 26 | 14 | 0.920 | 0.009 | −0.892 | −2.899 |
| Nyala | Aaf/wild | 10 | 10 | 8 | 0.933 | 0.004 | −1.507 | −4.469 |
| Al Fasher | Aaf/wild | 21 | 5 | 4 | 0.610 | 0.003 | 1.076 | 1.690 |
| Al Junaynah | Aaf/wild | 5 | 11 | 3 | 0.700 | 0.007 | −1.200 | 2.054 |
| Site (form) | P (Aaa) | T (Aaa) | K (Aaa) | G (Aaa) | D (Aaf) | N (Aaf) | F (Aaf) |
|---|---|---|---|---|---|---|---|
| P (Aaa) | |||||||
| T (Aaa) | 0.425 | ||||||
| K (Aaa) | 0.125 | 0.158 | |||||
| G (Aaa) | 0.389 | 0.097 | 0.045 | ||||
| D (Aaf) | 0.652 | 0.248 | 0.396 | 0.186 | |||
| F (Aaf) | 0.848 | 0.355 | 0.545 | 0.309 | 0.015 | ||
| N (Aaf) | 0.837 | 0.421 | 0.603 | 0.384 | 0.160 | 0.191 | |
| J (Aaf) | 0.782 | 0.252 | 0.352 | 0.124 | 0.037 | 0.205 | 0.366 |
| Site (form) | P (Aaa) | T (Aaa) | K (Aaa) | G (Aaa) | D (Aaf) | N (Aaf) | F (Aaf) | J (Aaf) |
|---|---|---|---|---|---|---|---|---|
| P (Aaa) | ||||||||
| T (Aaa) | 0.000 | |||||||
| K (Aaa) | 0.021 | 0.014 | ||||||
| G (Aaa) | 0.000 | 0.030 | 0.172 | |||||
| D (Aaf) | 0.000 | 0.000 | 0.000 | 0.001 | ||||
| N (Aaf) | 0.000 | 0.000 | 0.000 | 0.002 | 0.267 | |||
| F (Aaf) | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.014 | ||
| J (Aaf) | 0.000 | 0.008 | 0.007 | 0.087 | 0.156 | 0.004 | 0.000 |
| Source of Variation | df | Sum of Squares | Variance Components | Percentage of Variation |
|---|---|---|---|---|
| Among Groups | 2 | 131.012 | 1.310 | 39.22 |
| Among Individuals within Groups | 5 | 27.672 | 0.243 | 7.26 |
| Within Populations | 131 | 234.244 | 1.788 | 53.53 |
| Total | 138 | 392.928 | 3.341 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abuelmaali, S.A.; Jamaluddin, J.A.F.; Noaman, K.; Allam, M.; Abushama, H.M.; Elnaiem, D.E.; Ishak, I.H.; Wajidi, M.F.F.; Jaal, Z.; Abu Kassim, N.F. Distribution and Genetic Diversity of Aedes aegypti Subspecies across the Sahelian Belt in Sudan. Pathogens 2021, 10, 78. https://doi.org/10.3390/pathogens10010078
Abuelmaali SA, Jamaluddin JAF, Noaman K, Allam M, Abushama HM, Elnaiem DE, Ishak IH, Wajidi MFF, Jaal Z, Abu Kassim NF. Distribution and Genetic Diversity of Aedes aegypti Subspecies across the Sahelian Belt in Sudan. Pathogens. 2021; 10(1):78. https://doi.org/10.3390/pathogens10010078
Chicago/Turabian StyleAbuelmaali, Sara Abdelrahman, Jamsari Amirul Firdaus Jamaluddin, Kheder Noaman, Mushal Allam, Hind Mohammad Abushama, Dia Eldin Elnaiem, Intan Haslina Ishak, Mustafa Fadzil Farid Wajidi, Zairi Jaal, and Nur Faeza Abu Kassim. 2021. "Distribution and Genetic Diversity of Aedes aegypti Subspecies across the Sahelian Belt in Sudan" Pathogens 10, no. 1: 78. https://doi.org/10.3390/pathogens10010078
APA StyleAbuelmaali, S. A., Jamaluddin, J. A. F., Noaman, K., Allam, M., Abushama, H. M., Elnaiem, D. E., Ishak, I. H., Wajidi, M. F. F., Jaal, Z., & Abu Kassim, N. F. (2021). Distribution and Genetic Diversity of Aedes aegypti Subspecies across the Sahelian Belt in Sudan. Pathogens, 10(1), 78. https://doi.org/10.3390/pathogens10010078