Abstract
This study investigated the Yongning Fortress ruins in Taiyuan through a comprehensive analytical approach employing scanning electron microscopy (SEM), energy-dispersive X-ray spectroscopy (EDS), laser particle size analysis, X-ray diffraction (XRD), X-ray fluorescence spectroscopy (XRF), and ion chromatography (IC). The research focused on elucidating the disturbance mechanisms and environmental impacts induced by the root systems of five representative herbaceous species on rammed earth structures. The results demonstrated distinct, species-specific disturbance patterns. Melica roots created three-dimensional network damage, Artemisia capillaris primarily caused deep root penetration, Fallopia aubertii exhibited coupled physical–chemical effects, Convolvulus arvensis induced shallow horizontal expansion damage, while Cirsium formed a heterogeneous structure characterized by dense taproots and loose lateral roots. Environmental conditions, particularly moisture content, significantly influenced disturbance intensity. All root activities led to common deterioration processes, including particle rounding, gradation degradation, and formation of organic–mineral composites. Notably, vegetation markedly altered soluble salt distribution patterns, with Cirsium increasing total salt content to 3.7 times that of undisturbed rammed earth (0.48%), while sulfate ion concentration (1.16 × 10−3) approached hazardous thresholds. The study established a theoretical framework linking plant traits, disturbance mechanisms, and environmental response, and proposed risk-based zoning strategies for preservation. These outcomes provide significant theoretical foundations and practical guidance for the scientific conservation of rammed earth heritage sites.
1. Introduction
The deterioration of earthen archeological sites is an internationally recognized research issue driven by environmental and material factors. Common deterioration patterns include surface exfoliation, crack propagation, salt crystallization, volume changes induced by wet–dry cycles, wind and rain erosion, as well as biological erosion. These processes often interact synergistically, accelerating the loss of mechanical properties and material degradation of the structures, with significant variations in mechanisms and rates observed across arid, semi-arid, and humid regions.
Internationally, research on the impact of vegetation on earthen sites widely acknowledges its dual role: root systems can mechanically reinforce the soil and regulate the micro-environment yet simultaneously exacerbate deterioration through physical wedging and biogeochemical processes. Current scholarship strives to differentiate the effects under various vegetation types, root architectures, and environmental contexts to develop scientifically informed vegetation management strategies for heritage conservation.
To date, studies on the influence of plants on the properties of earthen sites have established a multi-scale international framework of understanding. In terms of mechanical properties, living roots enhance site stability by increasing soil shear and tensile strength. For instance, Rezaei et al. [1] demonstrated that root-reinforced soil in Iranian earthen sites exhibits strengthened cohesion, correlating positively with root density and distribution uniformity. However, thick root growth can induce tensile cracks, particularly in soils with high moisture fluctuations, where root wedging significantly compromises structural integrity [2]. Regarding hydrological properties, plant transpiration can reduce soil water content and mitigate shrinkage, yet roots also form preferential water pathways that facilitate infiltration and local saturation, aggravating freeze–thaw cycles and salt transport [3]. In chemical weathering, root exudates and organic acids from litter decomposition significantly alter soil pH, promoting clay mineral dissolution and cementation loss. Notably, vegetation cover in high-evaporation zones may even intensify surface salt enrichment [4]. Furthermore, plant–climate interactions play a critical role: mosses and lichens in arid environments can reduce wind and water erosion through biocrust formation [5], whereas in humid settings, similar communities accelerate material decay via moisture retention and bio-drilling [6]. Recent studies also emphasize the indirect role of plants as ecosystem engineers, altering microhabitats and promoting animal burrowing or microbial colonization, thereby triggering chain-reaction degradation [7]. Nevertheless, research on herbaceous root dynamics, comparative plant–soil responses across moisture gradients, and multi-factor coupling mechanisms—such as plant-salt–climate interactions—remains underdeveloped, constraining the precision of vegetation management strategies in heritage conservation.
Notwithstanding these advances, critical research gaps persist. Current literature exhibits a strong bias toward woody species, with herbaceous impacts remaining underexplored; comparative analyses across moisture regimes are scarce; and investigations into coupled bio-climatic–geo-chemical degradation mechanisms are still in their infancy.
It is within this context that a deeper engagement with the international literature on the decay and conservation of earthen architecture becomes not only relevant but essential. Examining both historic and contemporary earthen structures across diverse climatic and cultural settings allows researchers to identify recurring material vulnerabilities—such as erosion, capillary rise, and salt crystallization—while also discerning degradation patterns unique to specific environmental and socio-cultural conditions. Seminal works in the field, including that of Ashurst and Ashurst, offer foundational guidance on practical conservation by systematically addressing the intrinsic properties of earth materials and the physical and chemical processes underlying their decay [8]. Complementing these broad overviews, region-specific and technology-focused studies provide nuanced insights into how local factors influence deterioration. For instance, Hallock’s analysis of pisé construction in Virginia [9] reveals how humidity fluctuations and freeze–thaw cycles affect rammed earth walls in temperate regions. Similarly, Macdonald and Morgan’s investigation of African-inspired earthen architecture in Louisiana [10] demonstrates how cultural transmission and adaptation to subtropical climates inform material performance and durability. Meanwhile, Maher and Madrigal’s research on rural Egyptian earth buildings [11] underscores the role of traditional maintenance practices and socio-economic factors in shaping degradation pathways. Further deepening the discourse, detailed structural and conservation assessments—such as Sebastián and Cultrone’s study on Andalusian tapial traditions [12] and Ruiz et al.’s work on the seismic rehabilitation of earthen churches in the Andean region [13]—shed light on the dynamic interplay between inherited building knowledge, contemporary engineering interventions, and persistent environmental pressures. These studies illustrate how traditional construction systems respond to stressors such as earthquakes and moisture and how modern conservation strategies can be informed by historical resilience. Moreover, longitudinal field analyses like Spennemann’s [14,15] diachronic documentation of pisé decay in Australia provide invaluable perspectives on the temporal dimension of decay. By tracking changes in material condition over extended periods, such research captures the progressive nature of environmental degradation, offering critical data on long-term weathering behaviors and the efficacy of various conservation treatments over time.
Together, this body of international scholarship not only elucidates shared challenges in earthen heritage conservation but also underscores the necessity of context-responsive strategies. In particular, it reveals that plant impacts must be situated within broader material–environment interactions, where vegetation acts as one mediator within a complex system of decay drivers [16,17]. Case studies confirm that vegetation often simultaneously plays protective and destructive roles, presenting significant challenges for management decisions regarding plant retention or removal [18,19,20,21]. Positively, root systems enhance soil stability through reinforcement and anchoring, while canopy coverage mitigates direct erosion from various natural forces—such as rainfall, weathering, solar radiation, thermal shock, evaporation, particulate pollution, and freeze–thaw cycles—and helps stabilize the microclimate [22,23,24,25,26]. However, adverse effects are equally notable: root growth causes wedging that promotes crack propagation and even local collapse; secreted organic acids, H+, and HCO3− accelerate chemical weathering; and decomposing dead roots further facilitate structural disintegration and pedogenesis [27,28,29]. These plant-induced deterioration phenomena are collectively termed “plant-induced deterioration,” with vascular plants being particularly destructive in the short term due to their vigorous physiological activity [30,31].
To systematically assess plant impacts, scholars have developed a multidimensional research framework. Fang et al. [32] proposed a three-level classification system based on vegetation type, invasiveness, and root characteristics, establishing a methodological basis for preliminary evaluation. Case studies reveal that herbaceous plants at the Xi’an Han Shenming Tai site accelerated crack development by forming water transport channels [33]; at the Ming Dynasty Datong Zhen rammed earth wall, woody plant roots caused significant wedging damage, while herbaceous plants induced gradual soilization through long-term water and nutrient absorption [34]. Notably, plant impacts exhibit significant spatial heterogeneity: moss communities in Qinghai’s Guide Ancient City demonstrated restorative functions through rhizoid networks that reinforced surface soil and reduced powdering rates via salt absorption, significantly enhancing resistance to rain erosion and disintegration [35,36,37,38]. Similar protective effects were verified at arid and semi-arid sites such as the Qinghai Ming Great Wall and Lanzhou Xiaguanying Fortress, manifested as increased surface penetration resistance, micro-crack repair, and reduced salt accumulation. In humid environments, however, mosses may negatively affect cultural layers or accelerate bioweathering [39,40], highlighting the critical regulatory role of environmental context in plant–site interactions [41,42,43].
Mechanistic research exhibits interdisciplinary characteristics. Zhao et al. [44] quantified the enhancement of soil tensile strength through root–soil composite shear tests, identifying root morphology and root content as key factors. Zhang et al. [45] established early-warning thresholds for root diameter and length parameters in artificial rockery cracks. Regarding dynamic processes, scholars note a coupling relationship between plant succession (e.g., lichen → moss → herbaceous → woody) and site degradation, with climate type indirectly influencing preservation status via vegetation diversity. For instance, humus layers at the Jinyang Ancient City site [46] accelerated soil decomposition, while the Alhagi sparsifolia community at Guazhou Suoyang City [35] induced alkali return and cracking. Particularly noteworthy is the role of plants in altering micro-environments and triggering secondary disasters: root-damaged areas in shrubs at the Qinghai Ming Great Wall [39] became habitats for animals, initiating chain-reaction biological damage.
Despite these advances, critical knowledge gaps persist. The current research landscape exhibits the following: (1) disproportionate focus on woody species, leaving herbaceous impacts poorly understood; (2) insufficient comparative analysis across moisture regimes; and (3) limited exploration of synergistic effects between biological, climatic, and geochemical factors. The Yongning Fortress in Taiyuan, China, as a representative arid region rammed earth heritage site, provides an ideal case study to address these gaps. This investigation employs multiscale analytical methods to elucidate early-stage herbaceous root impacts on rammed earth structures, with dual objectives: advancing fundamental understanding of plant-rammed earth interactions in heritage contexts and developing temporally sensitive management strategies that balance conservation needs with ecological preservation. The findings promise to inform more nuanced, evidence-based approaches to vegetation management in rammed earth heritage conservation worldwide.
2. Investigation of Herbaceous Plants at Yongning Fortress
2.1. Survey Sections and Methodology
This study conducted systematic surveys of herbaceous vegetation on the remaining wall structures of Yongning Fortress, specifically targeting the eastern section of the northern wall, the eastern wall, and the southern wall remnants as shown in Figure 1, with particular focus on distribution patterns along the exterior facades. The field survey was executed in late April 2024 to coincide with the early growth stage of various herbaceous species, optimizing documentation of plant community composition. Using the standard transect survey method, the ramparts were divided into 10 m segments as independent survey units. Species identification was performed through on-site morphological examination supplemented by authoritative references such as Flora of China. Detailed records were made of the dominant species and their distribution patterns within each segment.
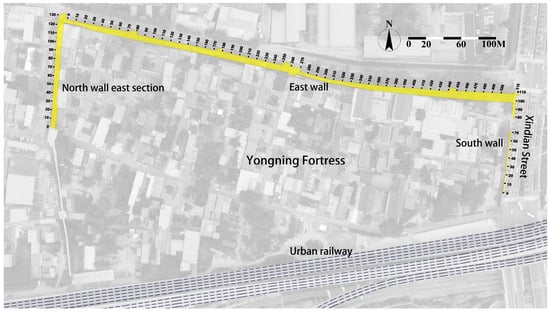
Figure 1.
Section division for herbaceous plant survey at Yongning Fortress Ruins.
2.2. Herbaceous Plant Composition and Distribution Patterns
The survey revealed distinct spatial distribution patterns of herbaceous vegetation across Yongning’s earthen walls. Vegetation density varied significantly by orientation; the northern and eastern walls exhibited particularly dense colonization, whereas the southern wall remained sparsely vegetated due to intense solar exposure and consequent substrate aridity. Ten dominant herbaceous species were identified, including Melica scabrosa Trin (1836), Artemisia capillaris Thunb (1784), et al., exhibiting clear vertical stratification:
The base zone (0–1.8 m height), characterized by accumulated collapse deposits and weathered, loosened rammed earth, served as the primary vegetation establishment area, with root systems typically penetrating 10–20 cm depth. The middle-collapsed zone (reaching 3.5 m in height) showed severe structural damage coupled with high vegetation coverage. The upper wall zone, where surface loosening occurred from rainwater erosion, supported herbaceous growth.
Notably, dominant species composition varied significantly by orientation. The northern wall was primarily affected by Melica scabrosa Trin and Fallopia aubertii (L. Henry) Holub (1971), while the eastern wall displayed alternating dominance between Melica scabrosa Trin and Cymbaria mongolica Maxim (1881). On the southern wall, shrub predominance substantially limited herbaceous development, creating markedly different vegetation patterns compared to other orientations.
2.3. Ecological Characteristics of Dominant Herbaceous Species at Yongning Fortress
The herbaceous vegetation at Yongning Fortress comprises ten species belonging to five plant families, as shown in Figure 2, exhibiting three key ecological adaptations. (1) Environmental resilience: Most species display exceptional drought tolerance and thrive in nutrient-poor substrates. Particularly noteworthy are the Asteraceae family members that exhibit specialized salt–alkali tolerance, enabling colonization of mineral-rich earthen walls. (2) Divergent growth strategies: Species evolved distinct root architectures to overcome water limitations. The fibrous root system of Melica scabrosa Trin forms dense, shallow networks to maximize surface water capture, while deep-rooted species like Fallopia aubertii Holub develops extensive taproots penetrating over 20 cm to access deeper moisture reserves. (3) Pronounced seasonal dynamics: Clear phenological patterns govern growth cycles, exemplified by Artemisia annua L observed in a senescent state during the survey period.

Figure 2.
Dominant herbaceous species at Yongning Fortress. (a) Melica scabrosa Trin; (b) Taraxacum mongolicum (1907); (c) Artemisia annua L. (1753); (d) Artemisia capillaris; (e) Convolvulus arvensis (1753); (f) Cirsium arvense (1772); (g) Cymbaria mongolica Maxim (1881); (h) Bothriospermum (1848); (i) Sonchus oleraceus L. (1753); (j) Fallopia aubertii.
2.4. Root System Characteristics of Herbaceous Plants
The herbaceous vegetation colonizing Yongning earthen ruins exhibits distinct root system morphologies and adaptive strategies, primarily categorized into taproot systems (e.g., Artemisia capillaris, Fallopia aubertii) and fibrous root systems (e.g., Convolvulus arvensis). Taproot-dominant species develop robust primary roots (0.5–0.7 cm in diameter) with pronounced vertical penetration. Fibrous-root species, in contrast, form dense lateral root networks (spanning up to 18 cm horizontally).
3. Sampling of Test Specimens
3.1. Sampling Methodology
Adhering strictly to minimally invasive principles, all samples were collected from concealed or collapsed sections using vibration-damped tools to ensure structural integrity (<0.1 mm displacement). Sampling followed a stratified 3D coordinate-based system, with triplicate samples taken at 0, 10, and 20 cm depths within both vegetated zones (focusing on five target species’ root systems) and undisturbed control areas (eastern wall, 260 m mark, 5 m elevation). Rigorous quality control measures included on-site physicochemical screening and a complete traceability chain. Post-sampling restoration followed strict traceability-based management protocols.
3.2. Sampling of Plant–Root Rammed Earth
Based on prior vegetation surveys, five representative herbaceous species were selected, as listed in Table 1. Based on the preliminary survey of herbaceous plants at the Yongning Fort ruins site, this study selected five representative dominant plant species and collected rammed earth samples from both vertical and horizontal ranges of their root systems. Plant selection followed these principles: (1) belonging to dominant species; (2) widespread distribution across different sections and areas of the site; (3) preference for representatives from distinct families and genera; and (4) consideration of diverse root system types.

Table 1.
Selection of herbaceous plants for testing.
Following these criteria, five herbaceous plants were ultimately chosen: Melica scabrosa, Artemisia capillaris, Fallopia aubertii, Convolvulus arvensis, and Cirsium arvense, as listed in Table 1. Melica scabrosa was widely distributed along the rammed earth walls. Sampling sites included the following areas: The north wall, near the wall base, approximately 5 m west of the sampling point. The east wall, near the northern end about 300 m along the wall, where severe collapse had reduced the height to less than half of its original. Samples were taken from the remnant top surface. The south wall, about 6 m west along the inner side of the wall, at a height of roughly 1 m above ground. Artemisia capillaris was primarily found along the base of the north wall, with samples collected approximately 51 m west along this section. Fallopia aubertii mainly grew on the middle-lower sections of the north and east walls. Due to significant structural damage near its growth points on the east wall, which posed stability risks, sampling was conducted on a plant located about 83 m west along the north wall. Convolvulus arvensis was predominantly distributed along the base of the east wall, with samples taken approximately 164 m north along this section. Cirsium arvense primarily grew along the base of the north wall, sampled about 66 m west along this segment. Following field sampling, all disturbed areas were restored in accordance with site conservation requirements.
3.3. Spatial Sampling Design
The study implemented a rigorous three-dimensional “Root Influence Zone” sampling model to systematically investigate spatial patterns of root-rammed earth interactions, as depicted in Figure 3. The sampling design incorporated concentric horizontal gradients radiating outward from the center collar axis (point 0; 0) to 10 cm (0–10) and 20 cm (0–20) radial distances, complemented by vertical stratification at 0 cm (surface), 10 cm (intermediate), and 20 cm (deep) depth levels.
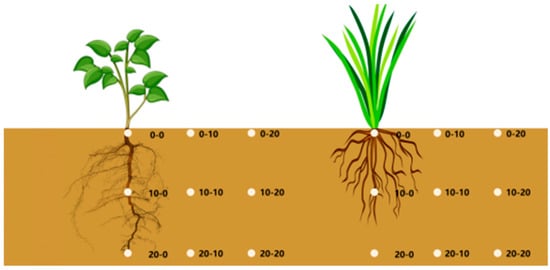
Figure 3.
Rammed earth measurement points for herbaceous plant growth locations.
4. Experimental Methods
To systematically evaluate the potential impacts of herbaceous plant roots on rammed earth heritage structures, a suite of microstructural, physical, and chemical analytical techniques was employed. The selection of each method was guided by its specific capacity to elucidate key deterioration mechanisms relevant to conservation practice.
4.1. Microstructure Observation
A combination of on-site video microscopy (60× magnification) and scanning electron microscopy (SEM) was used to conduct a comparative analysis of the microstructural characteristics of undisturbed rammed earth and root-affected zones. The primary objective was to visualize root–soil interaction interfaces, changes in pore structure, and the morphology of weathering products (e.g., salt crystals). Small representative fragments (0.5–2 cm3) are selected and mounted for observation.
4.2. Particle Size Distribution Test
Laser particle size analysis was performed to determine if root activity induces particle aggregation or dispersion, thereby altering key engineering properties such as shear strength, erodibility, and hydraulic conductivity. A representative sub-sample of homogenized, oven-dried, and disaggregated (but not ground) material (10–50 g) is needed.
4.3. Mineral Composition and Element Distribution Test
X-ray diffraction (XRD) and energy dispersive spectroscopy (EDS) were combined to qualitatively and quantitatively assess mineralogical composition and to detect evidence of element migration and enrichment at root–soil interfaces, providing insights into the chemical weathering of mineral phases by root exudates. A small amount of finely powdered, homogenized sample (1–5 g) is packed into a holder.
4.4. Oxide Composition and Content Test
X-ray fluorescence (XRF) spectrometry was utilized to measure bulk chemical changes and to identify the formation of secondary minerals, particularly salts, which are critical for assessing long-term durability. Samples consisted of finely powdered, homogenized samples (2–10 g) pressed into a pellet or fused into a glass bead.
4.5. Soluble Salt Ion Types and Content Test
Ion chromatography (IC) was employed to quantify the types and concentrations of soluble salts, with a central aim of evaluating how plant growth drives salt migration and accumulation, thus assessing the potential risk of salt crystallization damage. A larger representative sample (20–50 g) is needed to prepare rammed earth.
4.6. Loss on Ignition (LOI) Test
Following the standard for soil test methods, this test was crucial for assessing the input of organic matter from root activity (exudates, debris decomposition) and its implications for changing the soil’s chemical environment, microbial activity, and mineral stability. The standard typically specifies using 10–30 g of oven-dried, pulverized sample. The organic matter content was measured using the ignition method, calculated as follows:
where m1 = mass of crucible after ignition (g); m2 = mass of rammed earth sample (g); m3 = total mass of rammed earth sample and crucible after ignition (g).
Changes in LOI were analyzed to assess the influence of root activity on organic matter content and to explore the effects of biochemical processes on rammed earth stability.
4.7. Colorimetric Test
A colorimeter was used for on-site measurement of rammed earth color changes. Measurements are taken directly on the surface of a representative fragment or a smoothed, packed surface of a small amount of material (5–10 g) in a holder. The color values of undisturbed rammed earth (L0, a0, b0) served as the baseline, while root-affected rammed earth (L1, a1, b1) were compared using the color difference according to Equation
This quantitative assessment of color changes evaluates the visual impact of plant growth on heritage surfaces, integrating data on mineralogy, organic matter, and soluble salts for a comprehensive analysis of plant-induced rammed earth deterioration mechanisms.
5. Results and Discussion
5.1. Analysis of Microstructural Morphology
5.1.1. Undisturbed Rammed Earth Samples
The microscopic morphology of undisturbed rammed earth at different depths was observed using video microscopy and SEM, as shown in Figure 4 and Figure 5. It reveals that there are distinct depth-dependent weathering gradients in the undisturbed rammed earth unaffected by plant roots. The surface layer (0 cm) exhibits a structurally loose fabric attributed to prolonged exposure, the intermediate layer (10 cm depth) displays transitional pore development, and the deep layer (20 cm depth) maintains a dense structure, with localized grayish-white areas indicating calcareous deposits.

Figure 4.
Morphology of undisturbed rammed earth sampling points (×60). (a) 0 cm; (b) −10 cm; (c) −20 cm.

Figure 5.
SEM microscopic morphology of rammed earth particles in undisturbed rammed earth. (a) 100×; (b) 500×.
Under SEM at 100× magnification, the rammed earth particles appear randomly distributed with significant size heterogeneity. These particles were physically interlocked with surrounding debris-like materials, reflecting a relatively dense and intact microstructure. At 500× magnification, coarse particles exhibit elongated, angular, or irregularly rounded shapes, interlocking with each other. Concurrently, fine-grained components exhibited irregular, non-oriented arrangements characteristic of silt–clay fractions. At the micrometer scale, the crystalline structure remains relatively intact, indicating stable and high-strength rammed earth properties.
5.1.2. Rammed Earth at MST Growth Sites
The introduction of Melica scabrosa (MST) root systems significantly altered rammed earth microstructure and stability, exhibiting pronounced environment-dependent disturbance characteristics. Comparative observations of the north, east, and south walls found that the root zone of MST on the east wall exhibited the strongest disturbance effect. Its well-developed root network caused substantial damage in both vertical and horizontal directions. At 20 cm depth (measuring point 20-0), distinct root-induced fissures were still observable, as shown in Figure 6. These channels not only directly damaged the rammed earth structure but also provided preferential pathways for water infiltration, further accelerating the deterioration process of the rammed earth. Lateral roots of MST could still be detected at horizontal extensions of 10 cm and 20 cm (points 10-20 and 20-20), indicating its extensive root system with a wide disturbance range. The growth and development of roots disrupted the originally dense rammed earth structure at the growth sites, increasing its porosity and reducing its structural integrity and cohesion.

Figure 6.
Morphology of MST-root-affected rammed earth on the east wall. (a) 20-0; (b) 20-10; (c) 20-20.
This intense disturbance is associated with the unique micro-environment of the east wall. The collapsed condition at the top increased the rainwater-receiving area, and, combined with human activities, created favorable conditions for plant growth that significantly promoted root biomass accumulation. In contrast, the disturbance caused by MST on the south wall was weakest. Continuous sunlight exposure on the sun-facing slope maintained dry conditions that not only restricted root development but also enhanced the cementation of clay minerals, forming a natural anti-disturbance barrier. In this environment, MST could only develop sparse, fine lateral roots locally, causing relatively limited damage to the rammed earth structure.
The north wall presented an intermediate condition, displaying a unique alternating structure of “vertical looseness and horizontal compactness”. This pattern reflects the adaptive strategy of MST roots under moderate growth conditions, which is prioritizing main root development in the vertical direction where moisture conditions were relatively favorable, forming loose zones at measuring points 0-0, 10-0, and 20-0. While maintaining relatively dense original structures in areas with restricted horizontal expansion, this differentiated growth pattern resulted in an overall disturbance level on the north wall that was significantly lower than the east wall but higher than the south wall.
SEM observations of rammed earth samples from MST growth sites at 100× and 500× magnifications are shown in Figure 7. Compared with undisturbed rammed earth samples, the rammed earth particles at MST growth sites appeared looser (Figure 7a), with shapes transitioning from relatively elongated and angular to more rounded irregular particles (Figure 7b). Plant root hairs were interspersed within the rammed earth (Figure 7c), and the particle sizes at MST growth sites were generally smaller than those in undisturbed rammed earth, indicating that rammed earth particle structure had been disrupted by root activity.

Figure 7.
SEM microscopic morphology of MST-root-affected rammed earth on the east wall. (a) 100×; (b) 500×; (c) root hairs in rammed earth 300×.
5.1.3. Rammed Earth at ACT Growth Sites
The impact of Artemisia capillaris (ACT) roots on rammed earth structure demonstrates significantly different characteristics compared to MST. As shown in Figure 8, the rammed earth morphology around ACT growth sites exhibits typical deep-rooted damage patterns, where its taproot creates vertical cavities at 10 cm depth (point 10-0). These cavities directly compromise rammed earth continuity while establishing preferential flow paths for water infiltration that accelerate subsurface weathering.

Figure 8.
Morphology of ACT-root-affected rammed earth. (a) 10-0; (b) 10-10; (c) 10-20.
Notably distinct from MST, ACT shows relatively limited development of lateral roots, only fine lateral roots observed at point 10-10, while exhibiting exceptional vertical penetration capacity. This growth strategy concentrates mechanical disruption in deeper rammed earth strata, indicating greater destructive potential to foundational layers.
Comparative SEM analysis (Figure 9) confirms rammed earth degradation at ACT colonization sites; mineral particles display significant size reduction and morphological transformation from angular to rounded irregular shapes. These alterations directly reflect root-induced structural disaggregation and strength reduction, demonstrating that ACT’s deep-reaching taproot causes particle-scale weathering distinct from MST’s near-surface biomodification.

Figure 9.
SEM microscopic morphology of ACT-root-affected rammed earth. (a) 100×; (b) 500×.
5.1.4. Rammed Earth at FAH Growth Sites
The structural transformation of rammed earth by Fallopia aubertii (FAH) exhibits distinctive spatial differentiation patterns, where root activities reconstruct the rammed earth’s microarchitecture through a combination of multiple mechanisms. As illustrated in Figure 10, vertical profiles appear to show progressive structural loosening from the surface layer (0-0) to 20 cm depth (20-0), demonstrating comprehensive disturbance that directly corresponds with the vertical distribution of the root system. This transformation displays clear depth-dependent variations. At 10 cm depth (10-0), well-developed lateral roots create a characteristic radial fissure network, while at 20 cm depth (20-0), the effects of root exudates become predominant, resulting in noticeable particle aggregation phenomena. Horizontally, disturbance gradients emerge with dense fine rootlets within 10 cm of the plants (10-10 and 20-10), forming complex three-dimensional structures that induce directional realignment of rammed earth particles. Even at 20 cm distance from the plant (10-20 and 20-20), where disturbance intensity decreases, the diffusion effects of root metabolites remain detectable.

Figure 10.
Morphology of FAH-root-affected rammed earth; (a) 0-0; (b) 0-10; (c) 0-20; (d) 10-0; (e) 10-10; (f) 10-20; (g) 20-0; (h) 20-10; (i) 20-20.
SEM analysis (Figure 11) reveals a dual-phase modification regime. The one phase is macropore development at root–RE interfaces through physical displacement, as shown in Figure 11a. The other phase is localized consolidation via exudate-mediated particle aggregation, as shown in Figure 11b. This apparent contradiction arises from spatiotemporally segregated root activities: initial mechanical penetration creates void networks, while subsequent sustained exudation drives mineral transformation through moisture regulation and biogeochemical cementation.

Figure 11.
FAH microscopic morphology of MST-root-affected rammed earth. (a) 100×; (b) 500×.
5.1.5. Rammed Earth at CAL Growth Sites
Convolvulus arvensis (CAL) exhibits characteristic shallow horizontal oriented disturbance of rammed earth structures, as shown in Figure 12. Vertical profiles demonstrate relatively dense conditions in the surface 0–20 cm layer (0-0 to 0-20), while horizontal sections reveal distinct gradient variations. Within 10 cm of the plants (10-0 to 10-20), rhizome expansion generates a well-defined structural degradation zone corresponding to CAL’s lateral growth strategy.

Figure 12.
Morphology of CAL-root-affected rammed earth. (a) 10-0; (b) 10-10; (c) 10-20.
Under 100× SEM magnification (Figure 13a), the samples show minimal difference from undisturbed rammed earth. However, at 500× magnification (Figure 13b), clear modifications become apparent in the mineral particles at CAL. growth sites: they are smaller than those in undisturbed samples and have transformed into more rounded, irregular shapes. These alterations likely result from mechanical abrasion caused by persistent particle friction under horizontal rhizome pressure and accelerated surface erosion from frequent wet-dry cycling in the shallow root activity zone. Together, these processes have measurably impacted the structural stability and strength of the rammed earth.

Figure 13.
SEM microscopic morphology of CAL-root-affected rammed earth. (a) 100×; (b) 500×.
5.1.6. Rammed Earth at CAI Growth Sites
The morphological characteristics of rammed earth within Cirsium arvense (CAI) growth zones exhibit distinct spatial heterogeneity, as demonstrated in Figure 14. Adjacent to the primary growth area (0-0 to 0-20), the rammed earth surface appears dry and grayish-white with a characteristically dense structure. However, at a 10 cm longitudinal extension (10-0), the rammed earth fabric transitions to a notably looser consistency containing embedded lateral roots, revealing how secondary root development creates localized structural disturbance while maintaining limited spatial influence that diminishes with increasing distance from the plant center.

Figure 14.
Morphology of CAI-root-affected rammed earth. (a) 0-0; (b) 0-10; (c) 0-20; (d) 10-0; (e) 10-10; (f) 10-20.
SEM provides mechanistic insights into these root-rammed earth interactions (Figure 15). Under 100× magnification, the development of enlarged pores and intricate networks of fine root hairs illustrates how roots actively reshape the rammed earth’s pore architecture, establishing new preferential flow pathways. More remarkably, 500× magnification reveals two concurrent transformation processes; that is, mineral particles undergo size reduction while forming irregular aggregates that intimately associate with root structures, suggesting biochemical mediation through either root exudates or stimulated microbial activity that enhances particle cohesion and localized rammed earth consolidation.

Figure 15.
SEM microscopic morphology of CAI-root-affected rammed earth. (a) 100×; (b) 500×.
These microstructural observations reveal the dualistic nature of CAI’s impact on rammed earth. The taproot system creates zones of desiccated compaction through combined water uptake and physical compression, while lateral root expansion generates biologically disturbed, porous regions. This spatial dichotomy reflects the plant’s capacity to simultaneously create new void structures while reorganizing mineral particles through biogeochemical processes, ultimately forming distinctive root-rammed earth composites.
Comprehensive analysis of the root-induced rammed earth disturbance mechanisms exhibit species-specific patterns governed by microenvironmental factors, categorized into five characteristic typologies: (1) Three-dimensional network type (MST): the developed lateral and taproots create comprehensive spatial damage, with root-induced fissures enhancing pore connectivity and causing particle rounding. (2) Vertical taproot type (ACT): dominated by deep-penetrating taproots that form vertical root channels, with limited lateral root influence and noticeable particle size reduction. (3) Full-profile coupling type (FAH): combines physical fissuring with biochemical cementation, initially forming macropores and subsequently promoting mineral aggregation. (4) Shallow horizontal expansion type (CAL): rhizome spreading creates horizontal loosening zones within a 10 cm range, increasing particle roundness. (5) Localized heterogeneity type (CAI): features compacted zones from taproot water uptake alongside biologically disturbed areas from lateral roots, forming distinctive root-rammed earth composites.
The intensity of plant-induced modification of the rammed earth was strongly correlated with micro-environmental conditions, based on observed spatial patterns of disturbance. The primary correlating factor was moisture availability, with disturbance intensity greatest on the east wall (a collapse area with high water infiltration), intermediate on the north wall, and weakest on the south wall (where drought limited root development). Secondary factors included light and temperature: sustained sunlight on the south wall appeared to promote clay cementation, thereby forming more resistant barriers against root disturbance. Furthermore, human-induced structural damage (e.g., the roof collapse on the east wall) created conditions that enhanced both water infiltration and root biomass accumulation, correlating with significantly stronger deterioration. While these observed relationships are compelling and supported by known mechanisms, further controlled studies are needed to definitively establish causality.
Despite varying disturbance patterns, all affected rammed earths demonstrate particle rounding through combined mechanical abrasion and chemical weathering. Short-term effects are dominated by root-created tubular pores (e.g., MST, ACT), while long-term evolution leads to exudate-induced microaggregates (e.g., FAH, CAI). The cyclic wetting–drying of clay minerals (particularly in CAI taproot zones) and formation of organo–mineral complexes (through root exudate cementation) represent universal processes.
Consequently, conservation strategies should prioritize controlling MST (most destructive 3D networks), using the south wall’s dry conditions as a preservation reference. Selected species like CAI (rammed earth stabilization + infiltration enhancement) and FAH (pore-aggregate synergy) could be utilized for targeted rammed earth improvement applications, demonstrating how understanding species-specific root architectures enables development of differentiated conservation approaches that balance structural protection with ecological function.
5.2. Particle Size Distribution Analysis
The influence of plant root systems on the particle composition and structural stability of rammed earth is evident when comparing the particle size distribution characteristics between undisturbed rammed earth and root-affected rammed earth zones, as shown in Figure 16. The undisturbed rammed earth is dominated by silt particles (74.43–77.41%), supplemented by clay (11.22–13.77%) and sand (9.27–11.8%), forming a well-graded silty structure. Its uniform particle distribution, along with ideal coefficients of uniformity (Cu) and curvature (Cc), collectively maintains the dense state of the rammed earth. However, root-affected zones show systematic changes in particle composition; sand content increases to 13.29–17.08%, while silt and clay decrease to 73.57–76.05% and 9.35–11.24%, respectively. The reduction in both Cu and Cc indicates that root activity transforms fine particles into coarser aggregates.
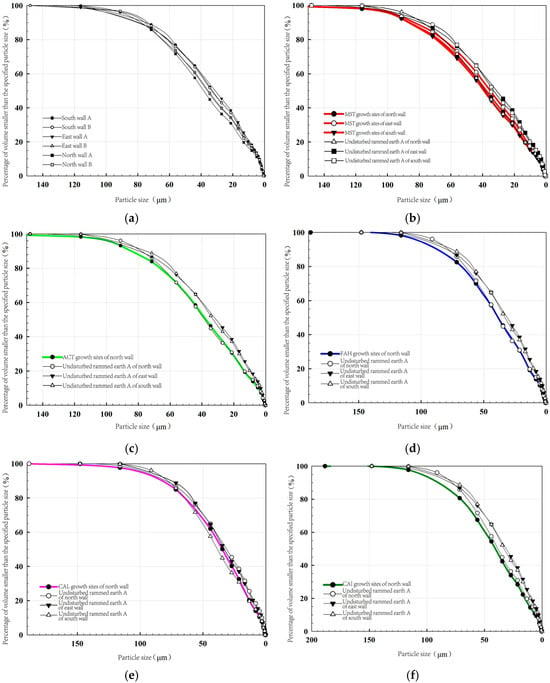
Figure 16.
Particle size distribution curve of different rammed earth samples. (a) Undisturbed rammed earth; (b) MST; (c) ACT; (d) FAH; (e) CAL; (f) CAI.
This particle redistribution arises from a dual mechanism of root action. The first is root-secreted mucilaginous polysaccharides and other organic substances that bind fine particles into larger aggregates through cohesive action. The second is root growth exerts mechanical pressure, while fibrous root networks occupy pore spaces, collectively reshaping the rammed earth’s microstructure.
The intensity of root influence varies significantly among plant species (CAI > MST > FAH > ACT > CAL), correlating with root morphology. For instance, CAI and MST, with their high root area ratio and dense fibrous roots, exhibit more pronounced fine-particle aggregation, whereas deep-rooted species like FAH and ACT, despite their robust taproots, have weaker micro-aggregation effects.
Although current herbaceous plants are in their early growth stages with underdeveloped root systems, their long-term cumulative effects on rammed earth structures remain a critical concern. Persistent particle aggregation progressively degrades the rammed earth gradation, replacing the original dense structure with a looser, more porous one. This microscale deterioration significantly reduces the mechanical strength of rammed earth. Coupled with external forces like wind and rain erosion, it accelerates surface weathering, crack propagation, and other forms of degradation, ultimately threatening the long-term preservation of rammed earth heritage sites. Heritage conservation practices must therefore prioritize controlling deep-rooted and fibrous herbaceous plants to disrupt this chain of progressive structural damage.
5.3. Mineral Composition and Elemental Distribution Analysis
Comparative mineral composition and elemental distribution of rammed earth samples provide profound insights into how plant roots modify rammed earth structures. Mineralogical examination (Figure 17) demonstrates that undisturbed rammed earth and root-affected rammed earth contain seven primary minerals: quartz, calcite, albite, dolomite, chamosite, muscovite, and microcline. Notably, no microcline was found at CAI growth sites, indicating that its root activity may exert a distinct influence on the stability and formation processes of this mineral.
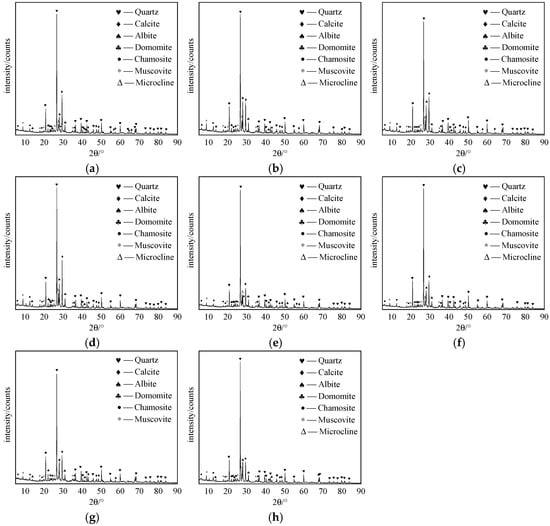
Figure 17.
Mineral composition of undisturbed and root-affected rammed earth samples. (a) MST on the north wall; (b) MST on the east wall; (c) MST on the south wall; (d) ACT; (e) FAH; (f) CAL; (g) CAI; (h) Undisturbed.
Elemental distribution patterns show significant variations. Undisturbed rammed earth exhibits sparse carbon (C) distribution, reflecting its low organic content, while detectable nitrogen (N) and phosphorus (P) provide potential nutrients for plant growth (Figure 18). At different plant growth sites, the carbon accumulation gradient exhibited the following pattern: MST > CAI > CAL ≈ ACT ≈ FAH ≈ undisturbed rammed earth (Figure 19). This gradient pattern was directly linked to root system characteristics: species with fibrous root systems (MST and CAI) enhanced organic matter accumulation through high-density root hairs, while deep-rooted species (FAH and ACT) contributed less significantly due to lower root surface area ratio.

Figure 18.
Distribution of C, N, and P elements in undisturbed rammed earth. (a) C; (b) N; (c) P.

Figure 19.
Distribution of carbon in rammed earth at growth sites of five herbaceous plants. (a) MST; (b) ACT; (c) FAH; (d) CAL; (e) CAI.
Microscale mineralogical changes further illustrate root-mediated alterations. At ACT sites, calcite particles exhibit reduced size and dispersed distribution, suggesting root activity may promote dissolution–recrystallization processes. The detection of secondary minerals like chamosite and muscovite at MST sites implies potential root-induced mineral transformation.
Collectively, herbaceous plants not only redistribute organic matter but may also influence mineral stability and transformation, including CAI’s potential decomposition of microcline and plant-specific calcite morphology variations, offering new scientific understanding of long-term weathering mechanisms in rammed earth heritage sites. While currently subtle, these micro-scale changes could substantially modify rammed earth’s physicochemical properties through cumulative effects, warranting special attention in heritage conservation practices.
5.4. Oxide Composition and Content Analysis
Chemical analysis of oxide composition and content was conducted on undisturbed and root-affected rammed earth samples, as presented in Table 2. Yongning Fort’s rammed earth, derived primarily from loess, features high SiO2 content (corresponding to quartz minerals) and significant CaO levels, indicating artificial lime addition, forming the fundamental material framework of the structural walls. While the major oxide components (SiO2, CaO, Fe2O3, etc.) remain relatively stable across all five herbaceous plant growth sites compared to undisturbed rammed earth.
Table 2.
Oxide composition and content of each rammed earth sample.
The fluctuations in Na2O content (notably at ACT and CAI sites) and heterogeneous distribution of trace components (BaO, Y2O3, Cl) suggest plant-mediated selective element absorption or accumulation. This differentiation correlates with species-specific root physiology; for instance, Cl detection at ACT and CAI sites may relate to their halophytic adaptations.
A systematic enrichment of P2O5 and SO3 across all vegetated sites directly evidences plant-induced biogeochemical cycling. Phosphorus enrichment stems from plant residue decomposition and root exudates, while SO3 increase may exert dual effects through sulfate hydration, providing sulfur nutrition short-term yet potentially accelerating salt crystallization damage long-term. Critically, these chemical modifications, while not altering major oxide proportions, may progressively impact durability by destabilizing cementitious materials (e.g., sulfate attack on lime-based binders), modifying pore solution ionic composition, and generating expansive salts through SO3–water coupling effects. Such chemical weathering, combined with previously observed physical–biological processes (particle aggregation, organic matter accumulation), completes the chain of plant-rammed earth interactions.
5.5. Types and Contents of Soluble Salt Ions Analysis
Soluble salt composition of undisturbed and root-affected rammed earth samples listed in Table 3. The ionic profile shows four cations (Na+, K+, Ca2+, Mg2+) and four anions (F−, Cl−, NO3−, SO42−), with Ca2+ content (998.0 μg/g) being markedly higher than other cations, which is consistent with the traditional lime addition construction technique, as evidenced by whitish regions observed under SEM. K+ (56.2 μg/g) and Mg2+ (49.2 μg/g) follow in concentration, while Na+ registers the lowest level (7.4 μg/g). Notably, nitrogen exists exclusively as NO3− (34.7 μg/g) with no detection of NH4+ or NO2−, indicating fixed nitrate as the dominant inorganic nitrogen form. The absence of PO43− suggests phosphorus likely exists as organic phosphorus compounds or insoluble minerals (e.g., apatite) within the rammed earth.
Table 3.
Types and contents of soluble salt ions in rammed earth samples.
It is found that root systems significantly alter soluble salt distribution patterns, with overall impact intensity ranking as CAI (4764.0 μg/g) > MST (1658.9–1981.3 μg/g) > ACT (2375.9 μg/g) > CAL (2773.9 μg/g) > FAH (1957.7 μg/g). This differentiation stems from three root-mediated mechanisms: (1) Biochemical processes: Root exudates (citric/oxalic acids) dissolving minerals (dolomite, chamosite) to release Ca2+/Mg2+; rhizosphere microbes driving organic matter decomposition and nutrient conversion (e.g., nitrification); selective nutrient uptake (K+/NO3− preference) altering ionic balance. (2) Physical transport: Enhanced capillary action from transpiration concentrating Cl− (425.5 μg/g at CAI sites) and SO42− (1163.9 μg/g); root channels creating preferential flow paths for water–salt migration; rhizosphere dehydration causing salt concentration. (3) Environmental inputs: Atmospheric deposition contributing exogenous NO3− (188.3 μg/g at CAL sites) and SO42−; groundwater (containing Cl−/SO42−/Na+) replenishing salts via capillary rise.
Evaluation using Yang Shanlong’s salt damage thresholds (ω(SO42−) ≤ 1.3 × 10−3, ω(Cl−) ≤ 6.5 × 10−4, total salt ≤ 0.20%) provides a critical risk assessment framework. The analysis reveals that CAI sites present a severe salinization risk, with a total salt content (0.48%) 3.7 times that of undisturbed rammed earth (0.13%) and well above the conservative threshold. This massive exceedance indicates a high potential for accelerated salt weathering cycles. CAL (0.28%) and ACT (0.24%) sites indicate a moderate risk, while MST (0.17–0.20%) and FAH (0.20%) approach critical levels. Regarding specific ions, CAI’s SO42− content (1.16 × 10−3) nears its damaging threshold, suggesting a pronounced risk of sulfate-related crystallization damage. Furthermore, Cl− levels in ACT and CAI zones (4.34 × 10−4 and 4.26 × 10−4, respectively) are approximately 15 times higher than in undisturbed rammed earth (0.27 × 10−4), heightening the risk of hygroscopic moisture uptake and cyclic dissolution–crystallization. While these thresholds are proxies for material vulnerability rather than precise predictors of failure timing, their consistent exceedance underlines a significantly elevated risk profile. These conditions are widely documented to initiate and exacerbate erosion, powdering, and loss of cohesion, thereby threatening the structural integrity of the rammed earth over time. Therefore, these results prioritize CAI and other high-salinity species for immediate management to mitigate long-term deterioration.
Salt-induced damage mechanisms include crystallization pressure from phase transitions Na2SO4 volume change up to 314%; hydration pressure from salt hydration (Na2SO4·10H2O generating ≤200 MPa); thermal expansion mismatch between salts and rammed earth matrix; salt-mediated particle aggregation altering gradation; pore clogging by salt crystals; and weakened rammed earth-salt interfacial bonds.
These findings demonstrate that plant roots fundamentally modify rammed earth salt regimes through complex biogeochemical processes, directly impacting heritage preservation. Priority measures should include controlling high salt-accumulating plants like Cirsium, optimizing drainage to reduce capillary rise, implementing sand–gravel barriers to limit salt migration, and developing species-specific vegetation management protocols based on salt accumulation potential.
5.6. LOI Analysis
The LOI analysis (Table 4) reveals significant organic matter redistribution in root-affected rammed earth. The undisturbed rammed earth exhibits a baseline LOI value of 10.67%, while root-affected rammed earth areas show elevated levels ranging from 10.85% to 14.98%. Notably, the CAL site displays the most substantial increase at 14.98%, representing a 40.4% rise compared to the undisturbed rammed earth. This organic accumulation exhibits distinct species-specific patterns. CAL and FAH sites exhibit the highest LOI values, reflecting their different root system characteristics. CAL’s fibrous roots contribute more labile organic materials, while FAH’s deep root system introduces lignin-rich compounds that undergo greater mass loss during combustion. Interestingly, although the MST site showed higher carbon content in elemental analysis, its relatively modest LOI increase (2.6–7.3%) suggests its organic matter exists in more stable forms, potentially as humified complexes or mineral-associated organics.
Table 4.
LOI of each rammed earth sample.
These organic matter changes exert dual effects on rammed earth conservation. The increased organic content enhances microbial activity that may accelerate mineral weathering (particularly calcite dissolution) and modifies pore structure, reducing bulk density by approximately 0.05 g/cm3 for every 1% LOI increase.
5.7. Colorimetric Analysis
Comparative chromaticity analysis reveals that plant growth induces significant and visually detectable color variations (ΔE > 3) in rammed earth at heritage sites. FAH demonstrates the most pronounced impact (maximum = 37.54), likely attributable to its deep-rooting characteristics that facilitate subsurface mineral transformation and organic matter input. Spatial patterns of chromatic alteration exhibit distinct gradients. Horizontally, the 10–20 cm zone surrounding growth points shows greater color variation than direct rhizosphere areas, suggesting a diffusion gradient effect of root exudates. Vertically, species-specific influence depths emerge, CAL exerts strongest chromatic effects at 10 cm depth ( = 15.95), while MST induces maximal color change at 20 cm depth.
These chromatic modifications were caused by the following reasons: (1) Organic matter accumulation alters light absorption properties, evidenced by significant correlation between elevated LOI values (particularly CAL’s 14.98%) and decreased b* values (yellowness–blueness axis). (2) Microtopographic changes from wind/water erosion increase surface roughness, causing amplified a* (redness–greenness) value fluctuations. Notably, MST growth points (north/south walls) show < 3, adjacent areas exhibit abnormal values (e.g., = 23.80 at south wall 10-10 measurement point), implying root-mediated hydrological modification may indirectly affect color distribution.
For conservation practice, it is recommended to establish a multi-parameter warning system integrating (>10 threshold), LOI increase (>15%), and microbial activity, and implement vegetation control when thresholds are exceeded.
5.8. Integrated Findings
- (1)
- The differential disturbance patterns of five typical herbaceous plants on rammed earth structures were classified. MST develops extensive 3D fibrous root networks, creating interconnected pore channels with horizontal–vertical damage synergy. ACT exhibits deep taproot penetration (>20 cm), threatening structural stability at depth. FAH displays the most complex disturbance, combining physical cracking with biochemical cementation. CAL primarily causes shallow (0–10 cm) horizontal expansion damage. CAI forms a unique “dense taproot–loose lateral root” heterogeneous structure.
- (2)
- Microenvironmental conditions significantly influence root disturbance intensity. East wall collapse zones enhance MST biomass and damage through increased water availability, whereas south wall sunlight exposure restricts root growth while promoting protective clay–mineral cementation that forms natural anti-disturbance barriers. North wall moisture gradients produce transitional “vertically loose-horizontally dense” structures.
- (3)
- All disturbed rammed earth exhibits the following common changes: increased particle roundness, reduced angular grains, a shift from silt (74.43–77.41%) to sand (13.29–17.08%) content, progressive “physical pore formation (early stage) to biochemical aggregation (later stage)” evolution, leading to a decline in structural strength.
- (4)
- Root systems significantly alter soluble salt distribution. CAI increases total salt content to 3.7 times that of the undisturbed rammed earth. SO42− in CAI zones (1.16 × 10−3) approaches the damage threshold (1.3 × 10−3). Cl− enrichment exceeds 15× in ACT and CAI zones. Salt transport is driven by transpiration and root channels.
- (5)
- The undisturbed rammed earth exhibits an LOI value of 10.67%, while plant-rooted areas show elevated levels ranging from 10.85% to 14.98%. The color difference () generally exceeds the recognizable threshold (>3), with FAH having the greatest impact (37.54).
- (6)
- Priority should be given to controlling high-risk species such as CAI and MST. An integrated engineering system combining drainage and salt barrier measures should be established. A multi-parameter early warning system monitoring color difference ( > 10 threshold), organic matter content (LOI increase > 15%), and microbial activity should be developed, with intervention protocols activated when threshold values are exceeded. The dry microenvironment conditions observed in the south wall section are recommended as a reference model for conservation practice.
5.9. Discussion
5.9.1. Theoretical and Practical Implications for Rammed Earth Heritage Biodeterioration
This study significantly refines the mechanistic understanding of plant-induced biodeterioration in rammed earth heritage. It demonstrates that root impacts constitute a coupled biogeophysical and biogeochemical reorganization process, rather than mere physical disruption or isolated salt accumulation. Critically, we established a direct link between species-specific root architectures (exemplified by CAI’s dense taproot with loose lateral roots), site-specific microenvironmental gradients (e.g., enhanced clay–mineral cementation on the sun-exposed south wall), and resultant multi-phase material alteration (as seen in FAH’s combined physical and biochemical disturbance). This evidence resolves the longstanding debate in the literature [16,19,21] by conclusively showing that plant damage is a synergistic process wherein physical and chemical mechanisms are interdependent and co-evolve.
These findings bridge a critical gap between general soil science research on root–soil interactions and the specific need for heritage conservation models. While prior studies have broadly acknowledged vegetation’s dual role [5,7,8], our results provide a granular, mechanistic framework that is directly translatable to conservation practice. It moves the field beyond qualitative assessments towards a predictive understanding of decay pathways.
5.9.2. Novelty and Relevance to Conservation Science
Within this framework, this study offers three key advances that address specific limitations highlighted in the introductory review:
- (1)
- A risk prioritization framework: The study provides the first quantitative, evidence-based hierarchy for species-specific removal priority (CAI > MST > ACT). By quantifying disturbance intensity—such as CAI’s localized salt enrichment to 3.7× baseline levels or FAH’s significant color change (ΔE = 37.54)—it offers conservation managers a defensible scientific rationale for allocating often limited resources to the most threatening species, a practical challenge frequently encountered in field conservation [22,23].
- (2)
- Microenvironment as a decay mediator: The results robustly demonstrate that wall orientation (e.g., south vs. east) fundamentally alters root behavior and resultant decay morphology through differential moisture and light gradients. This finding directly addresses the pronounced research bias towards generalized models that overlook microenvironmental heterogeneity [34,35]. It provides a compelling explanation for the frequent failure of standardized plant removal protocols and underscores the necessity of developing context-specific conservation strategies that account for microclimatic extremes, as seen in other regional case studies [24,35].
- (3)
- Proactive early-warning diagnostics: The proposed quantitative thresholds (e.g., ΔE > 10 for significant chemical alteration, LOI increase > 15% for organic enrichment) offer field-applicable tools for identifying incipient decay before severe structural damage occurs. This shifts the conservation paradigm from reactive interventions to proactive monitoring, a transition called for by recent research but lacking in practical methodologies [47,48]. These parameters serve as tangible indicators for the “plant-disease” conceptual models previously described [16,18].
5.9.3. Reconciling with Broader Biodeterioration Paradigms
- (1)
- Salt-driven deterioration: The mechanism of deterioration driven by sulfate salts (e.g., CAI’s SO42− approximating known damage thresholds) mirrors processes observed in porous stone heritage. However, this study demonstrates that the risk is severely exacerbated in earthen matrices by transpiration-induced capillary rise, a process less dominant in stone. This confirms the need to adapt salt weathering models from lithic to earthen contexts, rather than directly applying them.
- (2)
- The pore system evolution: The observed evolution from physical root penetration to biochemical weathering within the pore network parallels bioweathering mechanisms in sandstone. A novel finding for earthen materials is that the dominance of clay minerals can enable a protective response through recrystallization and cementation (as on the arid south wall), acting as a previously unrecognized buffering mechanism against complete structural collapse. This suggests that earthen materials may possess a degree of resilience that is not fully accounted for in current decay models.
- (3)
- These contrasts underscore a central conclusion: earthen architecture requires decay models distinct from those developed for stone or wood. The unique interplay between biogeochemistry (organic acids, salt cycling) and the physicochemical nature of clay-rich matrices necessitates the development of material-specific conservation strategies. This study provides a critical step towards building that dedicated theoretical framework, effectively answering the call for a more nuanced understanding of plant–site interactions raised in the literature review [6,33,35].
5.9.4. Implications for Holistic Conservation Management
While this study provides a mechanistic and quantitative assessment of the biodeterioration risks associated with specific herbaceous plants, it is imperative to situate these findings within a broader conservation ethic. Vegetation at heritage sites is not merely a pathological agent; it can constitute an integral part of the site’s ecosystem, contributing to biodiversity, preventing wind erosion of uncolonized areas, and potentially holding cultural or esthetic value as part of the site’s recent history and perceived ‘natural’ setting. A management strategy based solely on our material risk assessment would be reductive and potentially counterproductive.
Therefore, the prioritization of species for control (e.g., CAI, MST) should not be interpreted as a prescription for wholesale removal but as a critical input for a value-based decision-making process. For instance, a plant species identified as high-risk might be tolerated in a non-critical area where its ecological stabilizing function outweighs its slow deterioration potential, while its removal would be prioritized in areas of critical structural vulnerability.
6. Conclusions
This study systematically elucidates the species- and context-dependent mechanisms by which herbaceous plant roots contribute to the deterioration of rammed earth heritage structures. The key conclusion is that degradation is driven by a complex feedback loop involving simultaneous alterations to microstructure, chemistry, and mechanical properties. Consequently, effective conservation must extend beyond simple physical removal of plants. Instead, it requires integrated strategies that combine holistic microenvironment management—such as reducing moisture infiltration to emulate the protective, arid conditions observed on the south wall—with real-time diagnostic monitoring of critical decay indicators, including salt content, color change, and organic matter accumulation.
The mechanistic and predictive framework presented here provides a novel and essential foundation for developing material-specific conservation protocols, which are increasingly urgent in the context of climate change. Accordingly, management efforts should focus on controlling high-risk species, such as CAI and MST, particularly in areas where their presence directly jeopardizes structural stability. Nonetheless, decision-making must be guided by a comprehensive evaluation that also incorporates the ecological benefits and potential cultural significance of existing vegetation. For example, some species, including CAI (which may enhance stabilization and infiltration) and FAH (with its pore–aggregate synergy), warrant further investigation for potential targeted use in non-critical zones. This evidence-based, species-specific understanding enables the design of differentiated conservation strategies that aim to balance structural preservation with ecological integrity and ethical considerations.
Future research should establish an integrated framework synthesizing ecological succession dynamics, soil–structure interaction mechanisms, and biogeochemical cycling principles to quantify degradation feedback loops. It is essential to incorporate climate change variables—such as extreme precipitation and freeze–thaw cycles—alongside anthropogenic stressors (e.g., visitor traffic and pollution) and microclimate gradients through controlled experiments and geospatial monitoring. These critical mediating factors are currently underrepresented in root–soil interaction models. To move beyond static assessments, autonomous sensor networks should be deployed to track seasonal variations in salt migration, root exudation patterns, and microstructural changes. Non-destructive techniques such as X-ray μCT (for 3D pore network visualization) and ground-penetrating radar (for subsurface root mapping) can help correlate root architecture with crack propagation while preserving structural integrity.
To further enhance interpretive depth and interdisciplinary relevance, this mechanistic modeling should be contextualized within ecological functionality and digital conservation paradigms. For instance, computational approaches used in the study of traditional built environments—such as Liu et al.’s data-mining analysis of the ecologicality and functionality of kiln architecture in China—offer a methodological precedent for linking material decay with broader ecological and cultural dimensions [40]. Although focused on kiln structures, their analytical framework provides a valuable comparative perspective for interpreting plant-induced disturbances within the larger ecological–architectural systems of heritage sites.
Similarly, integrating insights from digital heritage conservation can significantly extend the implications of this research. Recent advancements summarized by Li et al. [47,48,49], including 3D modeling, IoT-based monitoring, and AI-driven diagnostics, demonstrate how digital tools are being used to address complex, multidimensional conservation challenges. Embedding empirical root–soil models within such digital frameworks would not only improve predictive accuracy but also facilitate the transition from observational correlations to culturally nuanced and preemptive preservation strategies, particularly under accelerating climate pressures.
Author Contributions
Methodology, X.J.; Writing—original draft, Investigation, X.C.; Investigation, Supervision, Review & editing W.H. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Shanxi Provincial Foundation Research Program (Grant No. 202203021211171), Shanxi Provincial Philosophy and Social Science Planning Project (Grant No.2023YJ025) and the High-Level Talent Recruitment Research Startup Fund of Shanxi Vocational University of Engineering Science and Technology (Grant No. RCK202307).
Data Availability Statement
Data will be available upon request.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Rezaei, M.; Azarnivand, H.; Ghorbani, A.; Tavili, A.; Abtahi, S.A. The effect of root reinforcement on the stability of earth slopes. J. Ecol. Eng. 2019, 20, 205–214. [Google Scholar]
- Ghestem, M.; Veylon, G.; Bernard, A.; Vanel, Q.; Stokes, A. Influence of plant root system morphology and architectural traits on soil shear resistance. Plant Soil 2014, 377, 43–61. [Google Scholar] [CrossRef]
- Viles, H.A. Plant-Environment Interactions in the Stone Degradation: A Review; Oxford University Press: Oxford, UK, 2022. [Google Scholar]
- Liu, T.; Zhao, Y.; Wu, X.; Song, C.; Li, J. Mechanisms of salt enrichment in surface soil under vegetation cover in arid regions: A review. Earth-Sci. Rev. 2020, 210, 103340. [Google Scholar]
- Wang, F.; Zhang, C.; Wang, X.; Liu, B. Biocrusts enhance the resistance of soil to erosion by water and wind in a semi-arid environment. Catena 2017, 157, 335–344. [Google Scholar]
- Carter, N.E.A.; Viles, H.A. Bioprotection explored: The story of a little known earth surface process. Geomorphology 2005, 67, 273–281. [Google Scholar] [CrossRef]
- Dulias, R. The impact of vegetation on the weathering of historical monuments. Environ. Socio-Econ. Stud. 2016, 4, 1–8. [Google Scholar]
- Ashurst, J.; Ashurst, N. Brick, Terracotta and Earth. In Practical Building Conservation: English Heritage Technical Handbook; Gower Technical Press: Aldershot, UK, 1988; Volume 2. [Google Scholar]
- Hallock, G. Pisé construction in early nineteenth-century Virginia. Perspect. Vernac. Archit. 2004, 11, 40–53. [Google Scholar]
- Macdonald, K.C.; Morgan, D.W. African earthen structures in colonial Louisiana: Architecture from the Coincoin plantation (1787–1816). Antiquity 2012, 86, 161–177. [Google Scholar] [CrossRef][Green Version]
- Maher, J.; Madrigal, J. Earth Architecture in Rural Egypt: Challenges of the context and the material. A+ Arch Des. Int. J. Archit. Des. 2021, 7, 99–112. [Google Scholar][Green Version]
- Sebastián, E.; Cultrone, G. Technology of Rammed-Earth Constructions (“Tapial”) in Andalusia (Spain): Their Restoration and Conservation. In Materials, Technologies and Practice in Historic Heritage Structures; Bostenaru Dan, M., Pøikryl, R., Török, A., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 11–28. [Google Scholar][Green Version]
- Ruiz, D.; López, C.; Unigarro, S.; Domínguez, M. Seismic rehabilitation of sixteenth-and seventeenth-century rammed earth–built churches in the Andean highlands: Field and laboratory study. J. Perform. Constr. Facil. 2015, 29, 04014144. [Google Scholar] [CrossRef]
- Spennemann, D.H.R. Diachronic Observations of the Decay of a Pisé Building at Jugiong (NSW); Institute for Land, Water and Society Report; Charles Sturt University: Albury, Australia, 2015. [Google Scholar]
- Spennemann, D.H.R. Earth to Earth: Patterns of environmental decay affecting historic and modern pisé walls. Buildings 2022, 12, 748. [Google Scholar] [CrossRef]
- Chen, X.; Bai, F.; Huang, J.; Lu, Y.; Wu, Y.; Yu, J.; Bai, S. The organisms on rock cultural heritages: Growth and weathering. Geoheritage 2021, 13, 56. [Google Scholar] [CrossRef]
- Liu, Y.; Jia, R.; Wang, W.; Wan, Y.; Gao, Y.; Zhan, H.; Zhu, J. Vascular plant communities and biocrusts act as controlling factors in mitigating soil erosion on the Great Wall in a semi-humid area of Northwestern China. Sci. Total Environ. 2024, 918, 170515. [Google Scholar] [CrossRef]
- Liu, T.; Zhao, X.W.; Liu, J.J.; Wang, K. Plant-induced diseases at an earthen site, using the Epang Palace Site as an example. Sci. Conserv. Archaeol. 2019, 31, 105–110. [Google Scholar]
- Yang, X.J.; Wu, F.; Jia, R.L.; Guo, Q.L.; Wang, Y.W.; Zhao, S.; Zhao, X.Y. Advances in research on the effects of plants on rock-soil relics and their conservation technology. Sci. Conserv. Archaeol. 2023, 35, 150–164. [Google Scholar]
- Cao, Y.; Bowker, M.A.; Delgado-Baquerizo, M.; Xiao, B. Biocrusts protect the Great Wall of China from erosion. Sci. Adv. 2023, 9, 5892. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Belnap, J. Growth responses of five desert plants as influenced by biological soil crusts from a temperate desert, China. Ecol. Res. 2015, 30, 1037–1045. [Google Scholar] [CrossRef]
- Zegeye, A.D.; Langendoen, E.J.; Tilahun, S.A. Root reinforcement to soils provided by common Ethiopian highland plants for gully erosion control. Ecohydrology 2018, 11, e1940. [Google Scholar] [CrossRef]
- Fabbri, K.; Canuti, G.; Ugolini, A. A methodology to evaluate outdoor microclimate of the archaeological site and vegetation role: A case study of the Roman Villa in Russi (Italy). Sustain. Cities Soc. 2017, 35, 107–133. [Google Scholar] [CrossRef]
- Hanssen, S.V.; Viles, H.A. Can plants keep ruins dry? A quantitative assessment of the effect of soft capping on rainwater flows over ruined walls. Ecol. Eng. 2014, 71, 173–179. [Google Scholar] [CrossRef]
- Li, Y.; Xia, C.; Wu, R. Role of the urban plant environment in the sustainable protection of an ancient city wall. Build. Environ. 2021, 187, 107405. [Google Scholar] [CrossRef]
- Coombes, M.A.; Viles, H.A.; Zhang, H. Thermal blanketing by ivy (Hedera helix L.) can protect building stone from damaging frosts. Sci. Rep. 2018, 8, 9834. [Google Scholar] [CrossRef]
- Sun, M.L.; Li, Z.X.; Wang, X.D. Classification of deteriorations associated with many rammed earth heritage sites in arid areas of northwest China. J. Eng. Geol. 2007, 6, 772–778. [Google Scholar]
- Viles, H.A.; Cutler, N.A. Global environmental change and the biology of heritage structures. Glob. Change Biol. 2012, 18, 2406–2418. [Google Scholar] [CrossRef]
- Lu, W.; Liu, C.; Liu, Q.; Pang, J.; Li, D. Influence of fiber content and length on the mechanical behavior of the grout slurry mixtures used for earthen sites conservation: A study of coir fibers and basalt fibers. npj Herit. Sci. 2025, 13, 85. [Google Scholar] [CrossRef]
- Xiao, B.; Bowker, M.A.; Zhao, Y.; Chamizo, S.; Issa, O.M. Biocrusts: Engineers and architects of surface soil properties, functions, and processes in dryland ecosystems. Geoderma 2022, 424, 116015. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Maestre, F.T.; Reich, P.B.; Jeffries, T.C.; Gaitan, J.J.; Encinar, D. Microbial diversity drives multifunctionality in terrestrial ecosystems. Nat. Commun. 2016, 7, 10541. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.Y.; Guo, K.; Wang, G.H.; Tang, Z.Y.; Xie, Z.Q.; Shen, Z.H.; Wang, R.Q.; Qiang, S.; Liang, C.Z.; Da, L.J.; et al. Vegetation classification system and classification of vegetation types used for the compilation of vegetation of China. Chin. J. Plant Ecol. 2020, 44, 96–110. [Google Scholar] [CrossRef]
- Yang, L.; Sun, M.L.; Huang, J.H.; Zhu, M.M.; Niu, P.; Bian, D. Analysis of major diseases and protection strategies of the Shenmingtai Site of the Han dynasty in Xi’an. Dunhuang Res. 2007, 5, 83–86. [Google Scholar]
- Shen, Y.; Xiang, R.; Chang, J.F. A study on construction technologies and preservation of the rammed-earth Great Wall of Ming Dynasty: A case study of the Datong Garrison. Archit. J. 2018, 2, 14–21. [Google Scholar]
- Du, Y.M.; Chen, W.W.; Cui, K.; Gong, S.Y.; Pu, T.B.; Fu, X.L. A model characterizing deterioration at earthen sites of the Ming Great Wall in Qinghai Province, China. Soil Mech. Found. Eng. 2017, 53, 426–432. [Google Scholar]
- Chen, W.W.; Yang, G.; Zhang, Y.M.; Liu, J.; Ren, X.F.; Gong, S.Y. Effect of moss soil crust on the scaling off of earthen sites in Qinghai. Adv. Eng. Sci. 2017, 49, 22–27. [Google Scholar]
- Zhao, H.Y.; Guo, R.H.; Duan, Y.T.; Zhang, H.T.; Chen, W.W. Experimental study on slope protection and soil reinforcement for moss. J. Lanzhou Univ. Nat. Sci. 2018, 54, 364–368. [Google Scholar]
- Li, Y.; Dong, Y.Z.; Li, W.Z.; Bai, Q.; Xu, J. Diversity research of the Bryophyta on the wall of Tongwan Castle Site. Chin. Wild Plant Resour. 2017, 36, 61–65. [Google Scholar]
- Pozo-Antonio, J.S.; Barreiro, P.G.; Paz-Bermúdez, G.; González, P.; Fernandes, A.B. Effectiveness and durability of chemical- and laser-based cleanings of lichen mosaics on schists at archaeological sites. Int. Biodeterior. Biodegrad. 2021, 163, 105276. [Google Scholar] [CrossRef]
- Zheng, Y.; Liu, S.B.; Zhang, Y.F.; Du, A.L.; Li, M.; Luo, H.J.; Zhu, J.F. Effect of in situ polymerization of super absorbent polymers on the protection of rammed earth heritage sites in semi-arid regions. Materials 2024, 19, 4839. [Google Scholar] [CrossRef]
- Ndidzulafhi, I.S. Sacred natural sites as other effective area-based conservation measures (OECMS) for biodiversity conservation in South Africa: Key opportunities and challenges for policy and practice. J. Nat. Conserv. 2025, 86, 126935. [Google Scholar] [CrossRef]
- Ferrari, P.; Rossi, P.L.; Bettuzzi, M.; Mariotti, F.; Morigi, M.P.; Riccardizi, C.; Canevari, C.; Modesti, P. X-ray imaging of a museum artefact on site: Monte Carlo simulations for radiation protection evaluations. J. Radiol. Prot. 2025, 45, 022501. [Google Scholar] [CrossRef]
- Wu, F.; Wang, W.F.; He, D.P.; Xu, R.H.; Su, B.M. Analysis of algae growing on the surface of the North City Wall of Liangzhu Site. Dunhuang Res. 2014, 4, 114–120. [Google Scholar]
- Zhao, Y.J.; Li, H.T.; Liu, C.Y. Shear tests of plant root-soil complex and present status and prospect of its mechanical model. Soil Water Conserv. China 2015, 10, 51–55. [Google Scholar]
- Zhang, Q.P.; Chen, F.Y. The influence of plant root splitting on garden rockeries in Jiangnan: Taking the Zhanyuan Garden as an example. J. Nanjing For. Univ. Nat. Sci. 2024, 48, 271–278. [Google Scholar]
- Ding, L.; Yang, Q.; Shi, D.S.; Wang, Q.Q.; Pei, J.R.; Hu, G. Chemical investigation of herbal wine from Jin Yang ancient city site during the late Western Han period in China. Archaeometry 2023, 66, 204–218. [Google Scholar] [CrossRef]
- Li, W.C.; Xie, Q.; Ao, J.H.; Lin, H.P.; Ji, S.S.; Yang, M.S.; Sun, J.H. Systematic review: A scientometric analysis of the status, trends and challenges in the application of digital technology to cultural heritage conservation (2019–2024). npj Herit. Sci. 2025, 13, 90. [Google Scholar] [CrossRef]
- Liu, R.Z.; Wang, X.; Yuan, L.; Gao, Y.T.; Zhang, Y.; Yang, M.S.; Li, W.C. Analyzing the ecologicality and functionality of kiln architecture in China through KH coder data mining algorithm and hierarchical event coding. Sci. Rep. 2025, 15, 16189. [Google Scholar] [CrossRef]
- Du, Y.M.; Chen, W.W.; Cui, K.; Zhang, K. Study on damage assessment of earthen sites of the Ming Great Wall in Qinghai Province based on Fuzzy-AHP and AHP-TOPSIS. Int. J. Archit. Herit. 2019, 14, 903–916. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).