
Integrating Molecular Docking and Electrophysiology Reveals Sesquiterpenes as Candidate Attractants for Ceratitis capitata Wiedemann (Diptera: Tephritidae)




Simple Summary
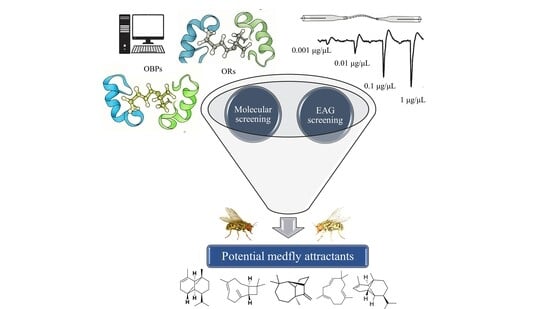
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Protein Molecular Models
2.2. Semiochemicals and Attractants
2.3. Docking Studies
2.4. Electroantennogram
2.5. Statistical Analysis
3. Results
3.1. Odorant-Binding Proteins
3.2. Odorant Receptors
3.3. Electroantennography
4. Discussion
5. Conclusions and Future Directions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| OBP | Odorant Binding Protein |
| OR | Odorant Receptor |
| EAG | Electroantennography |
| IPM | Integrated Pest Management |
References
- Lull, C.; Gil-Ortiz, R.; Cantín, Á. A Chemical Approach to Obtaining α-Copaene from Clove Oil and Its Application in the Control of the Medfly. Appl. Sci. 2023, 13, 5622. [Google Scholar] [CrossRef]
- Hafsi, A.; Abbes, K.; Harbi, A.; Chermiti, B. Field Efficacy of Commercial Food Attractants for Ceratitis capitata (Diptera: Tephritidae) Mass Trapping and Their Impacts on Non-Target Organisms in Peach Orchards. Crop Prot. 2020, 128, 104989. [Google Scholar] [CrossRef]
- Gómez-Escobar, E.; Alavez-Rosas, D.; Castellanos, D.; Quintero-Fong, L.; Liedo, P.; Malo, E.A. Effect of Aging on Three Lures Used for Monitoring Ceratitis capitata (Diptera: Tephritidae): Release Rate, Volatile Composition, and Fly Recaptures. J. Econ. Entomol. 2022, 115, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Jofré Barud, F.; López, S.; Tapia, A.; Feresin, G.E.; López, M.L. Attractant, Sexual Competitiveness Enhancing and Toxic Activities of the Essential Oils from Baccharis spartioides and Schinus polygama on Ceratitis capitata Wiedemann. Ind. Crops Prod. 2014, 62, 299–304. [Google Scholar] [CrossRef]
- Cossé, A.; Todd, J.; Millar, J.; Martinez, L.; Baker, T. Electroantennographic and Coupled Gas Chromatographic-Electroantennographic Responses of the Mediterranean Fruit Fly, Ceratitis capitata, to Male-Produced Volatiles and Mango Odor. J. Chem. Ecol. 1995, 21, 1823–1836. [Google Scholar] [CrossRef]
- Niogret, J.; Epsky, N.D. Attraction of Ceratitis capitata (Diptera: Tephritidae) Sterile Males to Essential Oils: The Importance of Linalool. Environ. Entomol. 2018, 47, 1287–1292. [Google Scholar] [CrossRef]
- Luu-Dam, N.; Tabanca, N.; Estep, A.; Nguyen, D.; Kendra, P. Insecticidal and Attractant Activities of Magnolia citrata Leaf Essential Oil Against Two Major Pests from Diptera: Aedes aegypti (Culicidae) and Ceratitis capitata (Tephritidae). Molecules 2021, 26, 2311. [Google Scholar] [CrossRef]
- Kharrat, S.; Gonzalez, F.; Rodriguez, C.; Calvo, C.; Gonzalez-Puschendorf, E.; Shelly, T.; Oehlschlager, C. Field Evaluation of α-Copaene Enriched Natural Oil Lure for Detection of Male Ceratitis capitata (Diptera: Tephritidae) in Area-Wide Monitoring Programs: Results from Tunisia, Costa Rica and Hawaii. Fla. Entomol. 2025, 108, 20240082. [Google Scholar] [CrossRef]
- Falchetto, M.; Ciossani, G.; Scolari, F.; Di Cosimo, A.; Nenci, S.; Field, L.M.; Mattevi, A.; Zhou, J.J.; Gasperi, G.; Forneris, F. Structural and Biochemical Evaluation of Ceratitis capitata Odorant-Binding Protein 22 Affinity for Odorants Involved in Intersex Communication. Insect Mol. Biol. 2019, 28, 431–443. [Google Scholar] [CrossRef]
- Siciliano, P.; He, X.; Woodcock, C.; Pickett, J.; Field, L.; Birkett, M.; Kalinova, B.; Gomulski, L.; Scolari, F.; Gasperi, G.; et al. Identification of Pheromone Components and Their Binding Affinity to the Odorant Binding Protein CcapOBP83a-2 of the Mediterranean Fruit Fly, Ceratitis capitata. Insect Biochem. Mol. Biol. 2014, 48, 51–62. [Google Scholar] [CrossRef]
- Vaníčková, L.; Do Nascimento, R.R.; Hoskovec, M.; Ježková, Z.; Břízová, R.; Tomčala, A.; Kalinová, B. Are the Wild and Laboratory Insect Populations Different in Semiochemical Emission? The Case of the Medfly Sex Pheromone. J. Agric. Food Chem. 2012, 60, 7168–7176. [Google Scholar] [CrossRef] [PubMed]
- Sollai, G.; Solari, P.; Crnjar, R. Olfactory Sensitivity to Major, Intermediate and Trace Components of Sex Pheromone in Ceratitis capitata Is Related to Mating and Circadian Rhythm. J. Insect Physiol. 2018, 110, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Kouloussis, N.; Mavraganis, V.; Damos, P.; Ioannou, C.; Bempelou, E.; Koveos, D.; Papadopoulos, N. Trapping of Ceratitis capitata Using the Low-Cost and Non-Toxic Attractant Biodelear. Agronomy 2022, 12, 525. [Google Scholar] [CrossRef]
- Guerrero, A.; Reddy, G.V.P. Chemical Communication in Insects: New Advances in Integrated Pest Management Strategies. Insects 2023, 14, 799. [Google Scholar] [CrossRef]
- Abendroth, J.; Moural, T.; Wei, H.; Zhu, F. Roles of Insect Odorant Binding Proteins in Communication and Xenobiotic Adaptation. Front. Insect Sci. 2023, 3, 1274197. [Google Scholar] [CrossRef]
- Sims, C.; Birkett, M.; Withall, D. Enantiomeric Discrimination in Insects: The Role of OBPs and ORs. Insects 2022, 13, 368. [Google Scholar] [CrossRef]
- Casaña-Giner, V.; Navarro-Llopis, V.; Levi, V.; Jang, E. Implication of SAR of Male Medfly Attractants in Insect Olfaction. SAR QSAR Environ. Res. 2002, 13, 629–640. [Google Scholar] [CrossRef]
- Scolari, F.; Valerio, F.; Benelli, G.; Papadopoulos, N.; Vaníčková, L. Tephritid Fruit Fly Semiochemicals: Current Knowledge and Future Perspectives. Insects 2021, 12, 408. [Google Scholar] [CrossRef]
- Zhang, J.; Luo, D.; Wu, P.; Li, H.; Zhang, H.; Zheng, W. Identification and Expression Profiles of Novel Odorant Binding Proteins and Functional Analysis of OBP99a in Bactrocera dorsalis. Arch. Insect Biochem. Physiol. 2018, 98, e21452. [Google Scholar] [CrossRef]
- Spanos, L.; Koutroumbas, G.; Kotsyfakis, M.; Louis, C. The Mitochondrial Genome of the Mediterranean Fruit Fly, Ceratitis capitata. Insect Mol. Biol. 2000, 9, 139–144. [Google Scholar] [CrossRef]
- Siciliano, P.; Scolari, F.; Gomulski, L.; Falchetto, M.; Manni, M.; Gabrieli, P.; Field, L.; Zhou, J.J.; Gasperi, G.; Malacrida, A. Sniffing out Chemosensory Genes from the Mediterranean Fruit Fly, Ceratitis capitata. PLoS ONE 2014, 9, e85523. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Jones, W.; Nguyen, T.; Kloss, B.; Lee, K.; Vosshall, L. Functional Conservation of an Insect Odorant Receptor Gene Across 250 Million Years of Evolution. Curr. Biol. 2005, 15, 119–121. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, R.P.; Santos, V.C.; Paranhos, B.A.G.; Aquino, N.C.; Nascimento, R.R.; Alencar-Filho, E.B. Odorant Binding Protein as a Management Target for Ceratitis capitata: A Window of Opportunities for In Vivo/In Silico Integration. Chemoecology 2025, 35, 47–72. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; De Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology Modelling of Protein Structures and Complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef]
- Krieger, E.; Vriend, G. YASARA View—Molecular Graphics for All Devices—From Smartphones to Workstations. Bioinformatics 2014, 30, 2981–2982. [Google Scholar] [CrossRef]
- Kim, S.; Thiessen, P.A.; Bolton, E.E.; Chen, J.; Fu, G.; Gindulyte, A.; Han, L.; He, J.; He, S.; Shoemaker, B.A.; et al. PubChem Substance and Compound Databases. Nucleic Acids Res. 2016, 44, D1202–D1213. [Google Scholar] [CrossRef]
- Hanwell, M.D.; Curtis, D.E.; Lonie, D.C.; Vandermeersch, T.; Zurek, E.; Hutchison, G.R. SOFTWARE Open Access Avogadro: An Advanced Semantic Chemical Editor, Visualization, and Analysis Platform. J. Cheminform 2012, 4, 17. [Google Scholar] [CrossRef]
- Morris, G.M.; Ruth, H.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. Software News and Updates AutoDock4 and AutoDockTools4: Automated Docking with Selective Receptor Flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2025. [Google Scholar]
- Mendiburu, F.; Simon, R.; De Mendiburu, F. Agricolae-Ten Years of an Open Source Statistical Tool for Experiments in Breeding, Agriculture and Biology. PeerJ Prepr. 2015, 3, e1404v1. [Google Scholar] [CrossRef]
- Doolittle, R.; Cunningham, R.; Mcgovern, T.; Sonnet, P. Trimedlure Enantiomers: Differences in Attraction for Mediterranean Fruit Fly Ceratitis capitata (Wied.) (Diptera: Tephritidae). J. Chem. Ecol. 1991, 17, 475–484. [Google Scholar] [CrossRef]
- Kamala, J.; Kempraj, V.; Aurade, R.; Roy, T.K.; Shivashankara, K.S.; Verghese, A. Computational Reverse Chemical Ecology: Virtual Screening and Predicting Behaviorally Active Semiochemicals for Bactrocera dorsalis. BMC Genom. 2014, 15, 209. [Google Scholar] [CrossRef]
- Warthen, J.D.; Schmidt, W.F.; Doolittle, R.E.; Cunningham, R.T. Structure-Activity Relationship Observations of Trans-Trimedlure Enantiomers. J. Chem. Ecol. 1995, 21, 69–79. [Google Scholar] [CrossRef] [PubMed]
- McGovern, T.P.; Warthen, J.D.; Cunningham, R.T. Relative Attraction of the Mediterranean Fruit Fly (Diptera: Tephritidae) to the Eight Isomers of Trimedlure. J. Econ. Entomol. 1990, 83, 1350–1354. [Google Scholar] [CrossRef]
- McGovern, T.; Cunningham, R.; Leonhardt, B. Cis-Trimedlure: Attraction for the Mediterranean Fruit Fly (Diptera: Tephritidae) and Isomeric Structural Assignments. J. Econ. Entomol. 1986, 79, 98–102. [Google Scholar] [CrossRef]
- Jang, E.B.; Light, D.M.; Dickens, J.C.; Mcgovern, T.P.; Nagata, J.T. Electroantennogram Responses of Mediterranean Fruit Fly, Ceratitis capitata (Diptera: Tephritidae) to Trimedlure and Its trans Isomers. J Chem Ecol 1989, 15, 2219–2231. [Google Scholar] [CrossRef]
- Niogret, J.; Gill, M.; Espinoza, H.; Kendra, P.; Epsky, N. Attraction and Electroantennogram Responses of Male Mediterranean Fruit Fly (Diptera: Tephritidae) to Six Plant Essential Oils. J. Entomol. Zool. Stud. 2017, 5, 958–964. [Google Scholar]
- Epsky, N.D.; Niogret, J. Short Range Attraction of Ceratitis capitata (Diptera: Tephritidae) Sterile Males to Six Commercially Available Plant Essential Oils. Nat. Volatiles Essent. Oils 2017, 4, 1–7. [Google Scholar]
- Shelly, T.; Cowan, E.; Edu, J.; Pahio, E. Mating Succes of Male Mediterranean Fruit Flies Following Exposure to Two Sources of α-Copaene, Manuka Oil and Mango. Fla. Entomol. 2008, 91, 9–15. [Google Scholar] [CrossRef]
- Candia, I.F.; Bautista, V.; Larsson Herrera, S.; Walter, A.; Ortuño Castro, N.; Tasin, M.; Dekker, T. Potential of Locally Sustainable Food Baits and Traps Against the Mediterranean Fruit Fly Ceratitis capitata in Bolivia. Pest. Manag. Sci. 2019, 75, 1671–1680. [Google Scholar] [CrossRef]
- Casaña-Giner, V.; Gandía-Balaguer, A.; Hernández-Alamós, M.M.; Mengod-Puerta, C.; Garrido-Vivas, A.; Primo-Millo, J.; Primo-Yúfera, E. Attractiveness of 79 Compounds and Mixtures to Wild Ceratitis capitata (Diptera: Tephritidae) in Field Trials. J. Econ. Entomol. 2001, 94, 898–904. [Google Scholar] [CrossRef]
- Jang, E.; Khrimian, A.; Holler, T. Field Response of Mediterranean Fruit Flies to Ceralure B1 Relative to Most Active Isomer and Commercial Formulation of Trimedlure. J. Econ. Entomol. 2010, 103, 1586–1593. [Google Scholar] [CrossRef]
- Santiago, F.P.; Alavez-Rosas, D.; Rojas, J.C. Rearing and 60Co Radiation Do Not Affect Attractiveness but Alter the Volatile Profiles Released by Anastrepha obliqua Calling Males. Bull. Entomol. Res. 2024, 114, 237–243. [Google Scholar] [CrossRef]
- Flath, R.; Cunningham, R.; Mon, T.; John, J. Additional Male Mediterranean Fruitfly (Ceratitis capitata Wied.) Attractants from Angelica Seed Oil (Angelica archangelica L.). J. Chem. Ecol. 1994, 20, 1969–1984. [Google Scholar] [CrossRef]
- Flath, R.; Cunningham, R.; Mon, T.; John, J. Male Lures for Mediterranean Fruitfly (Ceratitis capitata Wied.): Structural Analogs of α-Copaene. J. Chem. Ecol. 1994, 20, 2595–2609. [Google Scholar] [CrossRef] [PubMed]
- Niogret, J.; Montgomery, W.S.; Kendra, P.E.; Heath, R.R.; Epsky, N.D. Attraction and Electroantennogram Responses of Male Mediterranean Fruit Fly to Volatile Chemicals from Persea, Litchi and Ficus Wood. J. Chem. Ecol. 2011, 37, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Renthal, R. Arthropod Repellent Interactions with Olfactory Receptors and Ionotropic Receptors Analyzed by Molecular Modeling. Curr. Res. Insect Sci. 2024, 5, 100082. [Google Scholar] [CrossRef] [PubMed]
- Shelly, T.; Oehlschlager, C.; Kurashima, R. Natural Oil Lure Outperforms Trimedlure in Capturing Males of the Mediterranean Fruit Fly, Ceratitis capitata (Diptera: Tephritidae). Neotrop. Entomol. 2023, 52, 1138–1143. [Google Scholar] [CrossRef]
- Sierras-Serra, N.; Marin Garrido, C.; Botta Català, A.; Tait, G.; Merli, D.; Carlin, S.; Malacrida, A.R.; Gasperi, G.; Anfora, G.; Scolari, F. Electrophysiological Responses of the Mediterranean Fruit Fly, Ceratitis capitata, to the Cera Trap® Lure: Exploring Released Antennally-Active Compounds. J. Chem. Ecol. 2021, 47, 265–279. [Google Scholar] [CrossRef]
- Mai, J.; Li, W.; Ledesma-Amaro, R.; Ji, X.J. Engineering Plant Sesquiterpene Synthesis into Yeasts: A Review. J. Agric. Food Chem. 2021, 69, 9498–9510. [Google Scholar] [CrossRef]
- Anton, S.; Rössler, W. Plasticity and Modulation of Olfactory Circuits in Insects. Cell Tissue Res. 2021, 383, 149–164. [Google Scholar] [CrossRef]
- Serra, S. Enzyme-Mediated Synthesis of Sesquiterpenes. Nat. Prod. Commun. 2015, 10, 157–166. [Google Scholar] [CrossRef]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Share and Cite
Ordaz-Pérez, D.; Rojas, J.C.; Alavez-Rosas, D. Integrating Molecular Docking and Electrophysiology Reveals Sesquiterpenes as Candidate Attractants for Ceratitis capitata Wiedemann (Diptera: Tephritidae). Insects 2026, 17, 251. https://doi.org/10.3390/insects17030251
Ordaz-Pérez D, Rojas JC, Alavez-Rosas D. Integrating Molecular Docking and Electrophysiology Reveals Sesquiterpenes as Candidate Attractants for Ceratitis capitata Wiedemann (Diptera: Tephritidae). Insects. 2026; 17(3):251. https://doi.org/10.3390/insects17030251
Chicago/Turabian StyleOrdaz-Pérez, Daniela, Julio C. Rojas, and David Alavez-Rosas. 2026. "Integrating Molecular Docking and Electrophysiology Reveals Sesquiterpenes as Candidate Attractants for Ceratitis capitata Wiedemann (Diptera: Tephritidae)" Insects 17, no. 3: 251. https://doi.org/10.3390/insects17030251
APA StyleOrdaz-Pérez, D., Rojas, J. C., & Alavez-Rosas, D. (2026). Integrating Molecular Docking and Electrophysiology Reveals Sesquiterpenes as Candidate Attractants for Ceratitis capitata Wiedemann (Diptera: Tephritidae). Insects, 17(3), 251. https://doi.org/10.3390/insects17030251