
Genomic Insights and Bioconversion Potential in the Black Soldier Fly (Hermetia illucens): Current Advances and Future Directions
 ,
,  , and
, and
Simple Summary
Abstract
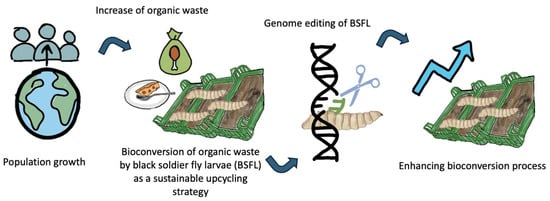
1. Introduction
2. The Black Soldier Fly Origin
3. The Black Soldier Fly Genetic Profile
3.1. BSF Affinity for Organic Substrates
3.2. BSF Environmental Adaptation
4. Genome Editing to Improve Performance Traits in the BSF
5. Limitations, Regulatory Frameworks and Industrial Considerations
6. Future Perspectives and Conclusions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| BSF | Black soldier fly |
| BSFL | Black soldier fly larvae |
| CRISPR/Cas9 | Clustered Regularly Interspaced Palindromic Repeats/CRISPR-associated protein 9 |
| EU | European Union |
| GMOs | Genetically modified organisms |
References
- United Nations. World Population Prospects 2022: Summary of Results; United Nations: New York, NY, USA, 2022. [Google Scholar]
- European Commission. Clean Air and Agriculture in the EU. 2023. Available online: https://agriculture.ec.europa.eu/sustainability/environmental-sustainability/natural-resources/clean-air_en (accessed on 15 October 2025).
- Kaza, S.; Yao, L.; Bhada-Tata, P.; Van Woerden, F. What a Waste 2.0: A Global Snapshot of Solid Waste Management to 2050; World Bank: Washington, DC, USA, 2018. [Google Scholar]
- Buzby, J. Food Waste and Its Links to Greenhouse Gases and Climate Change. U.S. Department of Agriculture. 2022. Available online: https://www.usda.gov/media/blog/2022/01/24/food-waste-and-its-links-greenhouse-gases-and-climate-change (accessed on 15 October 2025).
- Raksasat, R.; Lim, J.W.; Kiatkittipong, W.; Kiatkittipong, K.; Ho, Y.C.; Lam, M.K.; Font-Palma, C.; Zaid, H.F.M.; Cheng, C.K. A review of organic waste enrichment for inducing palatability of black soldier fly larvae: Wastes to valuable resources. Environ. Pollut. 2020, 267, 115488. [Google Scholar] [CrossRef]
- Siddiqui, S.A.; Ristow, B.; Rahayu, T.; Putra, N.S.; Yuwono, N.W.; Nisa’, K.; Mategeko, B.; Smetana, S.; Saki, M.; Nawaz, A.; et al. Black soldier fly larvae (BSFL) and their affinity for organic waste processing. Waste Manag. 2022, 140, 1–13. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. Food Wastage Footprint: Impacts on Natural Resources; FAO: Rome, Italy, 2013. [Google Scholar]
- Van Huis, A. Potential of insects as food and feed in assuring food security. Annu. Rev. Entomol. 2013, 58, 563–583. [Google Scholar] [CrossRef]
- Van Huis, A.; Oonincx, D.G.A.B. The environmental sustainability of insects as food and feed: A review. Agron. Sustain. Dev. 2017, 37, 43. [Google Scholar] [CrossRef]
- Woodley, N.E.; Thompson, F.C. A World Catalog of the Stratiomyidae (Insecta: Diptera); Backhuys Publishers: Leiden, The Netherlands, 2001. Volume 11. Available online: https://www.researchgate.net/publication/303225468 (accessed on 15 October 2025).
- Singh, A.; Kumari, K. An inclusive approach for organic waste treatment and valorisation using black soldier fly larvae: A review. J. Environ. Manag. 2019, 251, 109569. [Google Scholar] [CrossRef]
- Čičková, H.; Newton, G.L.; Lacy, R.C.; Kozánek, M. The use of fly larvae for organic waste treatment. Waste Manag. 2015, 35, 68–80. [Google Scholar] [CrossRef]
- Oliveira, F.; Doelle, K.; List, R.; O’Reilly, J.R. Assessment of Diptera: Stratiomyidae, genus Hermetia illucens (L., 1758) using electron microscopy. J. Entomol. Zool. Stud. 2015, 3, 147–152. Available online: https://www.researchgate.net/publication/305378530 (accessed on 15 October 2025).
- Tomberlin, J.K.; Sheppard, D.C.; Joyce, J.A. Selected life-history traits of black soldier flies (Diptera: Stratiomyidae) reared on three artificial diets. Ann. Entomol. Soc. Am. 2002, 95, 379–386. [Google Scholar] [CrossRef]
- Dortmans, B.M.A.; Diener, S.; Verstappen, B.M.; Zurbrügg, C. Black Soldier Fly Biowaste Processing: A Step-By-Step Guide; Swiss Federal Institute of Aquatic Science and Technology Eawag: Zürich, Switzerland, 2017. [Google Scholar]
- Wang, Y.-S.; Shelomi, M. Review of black soldier fly (Hermetia illucens) as animal feed and human food. Foods 2017, 6, 91. [Google Scholar] [CrossRef]
- Moretta, A.; Salvia, R.; Scieuzo, C.; Di Somma, A.; Vogel, H.; Pucci, P.; Sgambato, A.; Wolff, M.; Falabella, P. A bioinformatic study of antimicrobial peptides identified in the black soldier fly (BSF) Hermetia illucens (Diptera: Stratiomyidae). Sci. Rep. 2020, 10, 16875. [Google Scholar] [CrossRef]
- Park, S.-I.; Chang, B.S.; Yoe, S.M. Detection of antimicrobial substances from larvae of the black soldier fly, Hermetia illucens (Diptera: Stratiomyidae). Entomol. Res. 2014, 44, 58–64. [Google Scholar] [CrossRef]
- Lopes, I.G.; Lalander, C.; Vidotti, R.M.; Vinnerås, B. Reduction of bacteria in relation to feeding regimes when treating aquaculture waste in fly larvae composting. Front. Microbiol. 2020, 11, 1616. [Google Scholar] [CrossRef]
- Pang, W.; Hou, D.; Nowar, E.E.; Chen, H.; Zhang, J.; Zhang, G.; Li, Q.; Wang, S. The influence on carbon, nitrogen recycling, and greenhouse gas emissions under different C/N ratios by black soldier fly. Environ. Sci. Pollut. Res. 2020, 27, 42767–42777. [Google Scholar] [CrossRef]
- Pang, W.; Hou, D.; Chen, J.; Nowar, E.E.; Li, Z.; Hu, R.; Tomberlin, J.K.; Yu, Z.; Li, Q.; Wang, S. Reducing greenhouse gas emissions and enhancing carbon and nitrogen conversion in food wastes by the black soldier fly. J. Environ. Manag. 2020, 260, 110066. [Google Scholar] [CrossRef]
- Mertenat, A.; Diener, S.; Zurbrügg, C. Black soldier fly biowaste treatment—Assessment of global warming potential. Waste Manag. 2019, 84, 173–181. [Google Scholar] [CrossRef]
- Matos, J.S.; de Aráujo, L.P.; Allaman, I.B.; Lôbo, I.P.; de Oliva, S.T.; Tavares, T.M.; Neto, J.A.d.A. Evaluation of the reduction of methane emission in swine and bovine manure treated with black soldier fly larvae (Hermetia illucens L.). Environ. Monit. Assess. 2021, 193, 480. [Google Scholar] [CrossRef]
- Ermolaev, E.; Lalander, C.; Vinnerås, B. Greenhouse gas emissions from small-scale fly larvae composting with Hermetia illucens. Waste Manag. 2019, 96, 65–74. [Google Scholar] [CrossRef]
- European Commission. Circular Economy—Overview. Eurostat. Available online: https://ec.europa.eu/eurostat/web/circular-economy (accessed on 17 October 2025).
- Greenwood, M.P.; Hull, K.L.; Brink-Hull, M.; Lloyd, M.; Rhode, C. Feed and host genetics drive microbiome diversity with resultant consequences for production traits in mass-reared Black Soldier Fly (Hermetia illucens) larvae. Insects 2021, 12, 1082. [Google Scholar] [CrossRef]
- Muchina, P.; Kinyua, J.; Khamis, F.; Tanga, C.M.; Gebreyesus, G.; Cai, Z.; Sahana, G. The genomic landscape of Hermetia illucens: Exploring the potential of the Black Soldier Fly through molecular insights. J. Insects Food Feed 2025, 11, 2049–2074. [Google Scholar] [CrossRef]
- Suryati, T.; Julaeha, E.; Farabi, K.; Ambarsari, H.; Hidayat, A.T. Lauric acid from the black soldier fly (Hermetia illucens) and its potential applications. Sustainability 2023, 15, 10383. [Google Scholar] [CrossRef]
- Wang, H.; Rehman, K.U.; Liu, X.; Yang, Q.; Zheng, L.; Li, W.; Cai, M.; Li, Q.; Zhang, J.; Yu, Z. Insect biorefinery: A green approach for conversion of crop residues into biodiesel and protein. Biotechnol. Biofuels 2017, 10, 304. [Google Scholar] [CrossRef]
- Giani, M.; Valentino, C.; Vigani, B.; Ruggeri, M.; Guarnieri, A.; Salvia, R.; Scieuzo, C.; Falabella, P.; Sandri, G.; Rossi, S. Hermetia illucens-derived chitosan as a promising sustainable biomaterial for wound healing applications: Development of sponge-like scaffolds. Int. J. Biol. Macromol. 2025, 304, 140903. [Google Scholar] [CrossRef]
- International Platform of Insects for Food and Feed. Insect Frass as Fertiliser—EU Insect Producer Guidelines. IPIFF. 2025. Available online: https://ipiff.org/insects-frass/ (accessed on 17 October 2025).
- Kaya, C.; Generalovic, T.N.; Ståhls, G.; Hauser, M.; Samayoa, A.C.; Nunes-Silva, C.G.; Roxburgh, H.; Wohlfahrt, J.; Ewusie, E.A.; Kenis, M.; et al. Global population genetic structure and demographic trajectories of the black soldier fly, Hermetia illucens. BMC Biol. 2021, 19, 94. [Google Scholar] [CrossRef]
- Guilliet, J.; Baudouin, G.; Pollet, N.; Filée, J. The natural history of the black soldier fly, Hermetia illucens: Insights from complete mitochondrial genome sequences. bioRxiv 2021. [Google Scholar] [CrossRef]
- Hull, K.L.; Greenwood, M.P.; Lloyd, M.; Brink-Hull, M.; Bester-van der Merwe, A.E.; Rhode, C. Drivers of genomic diversity and phenotypic development in early phases of domestication in Hermetia illucens. Insect Mol. Biol. 2024, 33, 756–776. [Google Scholar] [CrossRef]
- Sandrock, C.; Leupi, S.; Wohlfahrt, J.; Kaya, C.; Heuel, M.; Terranova, M.; Blanckenhorn, W.U.; Windisch, W.; Kreuzer, M.; Leiber, F. Genotype-by-diet interactions for larval performance and body composition traits in the black soldier fly, Hermetia illucens. Insects 2022, 13, 424. [Google Scholar] [CrossRef]
- Muchina, P.; Kinyua, J.; Khamis, F.; Tanga, C.M.; Bett, R.; Ssepuuya, G.; Nakimbugwe, D.; Sinding, M.-H.S.; Gebreyesus, G.; Sahana, G.; et al. Human-mediated dispersal and breeding reshape global genomic patterns in black soldier flies. bioRxiv 2025. [Google Scholar] [CrossRef]
- Zhan, S.; Fang, G.; Cai, M.; Kou, Z.; Xu, J.; Cao, Y.; Bai, L.; Zhang, Y.; Jiang, Y.; Luo, X.; et al. Genomic landscape and genetic manipulation of the black soldier fly Hermetia illucens, a natural waste recycler. Cell Res. 2020, 30, 50–60. [Google Scholar] [CrossRef]
- Generalovic, T.N.; McCarthy, S.A.; Warren, I.A.; Wood, J.M.D.; Torrance, J.; Sims, Y.; Quail, M.; Howe, K.; Pipan, M.; Durbin, R.; et al. A high-quality, chromosome-level genome assembly of the black soldier fly (Hermetia illucens L.). G3 Genes Genomes Genet. 2021, 11, jkab085. [Google Scholar] [CrossRef]
- Albalawneh, A.; Al-Shagour, B.; Alarsan, S.; Sadder, M.; Alshorman, B.; Aledwan, M.; Deah, M.A.; Znaimah, S.A.; AlBalawnah, Y.; Neimat, E.; et al. Complete mitochondrial genome of a novel black soldier fly isolate from Jordan. Sci. Rep. 2025, 15, 27817. [Google Scholar] [CrossRef]
- Qi, Y.; Xu, J.; Tian, X.; Bai, Y.; Gu, X. The complete mitochondrial genome of Hermetia illucens (Diptera: Stratiomyidae). Mitochondrial DNA B Resour. 2017, 2, 189–190. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ståhls, G.; Meier, R.; Sandrock, C.; Hauser, M.; Šašić Zorić, L.; Laiho, E.; Aracil, A.; Doderović, J.; Badenhorst, R.; Unadirekkul, P.; et al. The puzzling mitochondrial phylogeography of the black soldier fly (Hermetia illucens), the commercially most important insect protein species. BMC Evol. Biol. 2020, 20, 60. [Google Scholar] [CrossRef] [PubMed]
- Gasco, L.; Biancarosa, I.; Liland, N.S. From waste to feed: A review of recent knowledge on insects as producers of protein and fat for animal feeds. Curr. Opin. Green Sustain. Chem. 2020, 23, 67–79. [Google Scholar] [CrossRef]
- Zhang, B.; Yang, R.; He, S.; Dai, S.; Hu, Q.; Li, X.; Su, H.; Shi, J.; Zhao, Z.; Wu, D. Swill and pig manure substrates differentially affected transcriptome and metabolome of the black soldier fly larvae. Int. J. Mol. Sci. 2024, 25, 12147. [Google Scholar] [CrossRef]
- Zhu, Z.; Rehman, K.U.; Yu, Y.; Liu, X.; Wang, H.; Tomberlin, J.K.; Sze, S.-H.; Cai, M.; Zhang, J.; Yu, Z.; et al. De novo transcriptome sequencing and analysis revealed the molecular basis of rapid fat accumulation by black soldier fly (Hermetia illucens, L.) for development of insectival biodiesel. Biotechnol. Biofuels 2019, 12, 194. [Google Scholar] [CrossRef]
- Silvaraju, S.; Zhang, Q.; Kittelmann, S.; Puniamoorthy, N. Genetics, age, and diet influence gut bacterial communities and performance of black soldier fly larvae (Hermetia illucens). Anim. Microbiome 2024, 6, 56. [Google Scholar] [CrossRef]
- Lin, S.-W.; Shelomi, M. Black soldier fly (Hermetia illucens) microbiome and microbe interactions: A scoping review. Animals 2024, 14, 3183. [Google Scholar] [CrossRef]
- IJdema, F.; Arias-Giraldo, L.M.; Vervoort, E.; Struyf, T.; Van den Ende, W.; Raaijmakers, J.M.; Lievens, B.; De Smet, J. Metagenome-based identification of functional traits of the black soldier fly gut microbiome associated with larval performance. BMC Microbiol. 2025, 25, 612. [Google Scholar] [CrossRef]
- Koch, H.; Paten, A.M.; Morgan, M.J. Microbiome enrichment from a wild relative improves black soldier fly larval survival, pathogen suppression and growth. bioRxiv 2025. [Google Scholar] [CrossRef]
- Li, Z.; Zhou, Y.; Lv, L.; Long, Y.; Dong, M.; Xiao, Q.; Jia, X.; Ju, D.; Sun, X.; Huang, F.; et al. Host-mediated environmental microbiome recruitment by black soldier fly (Hermetia illucens) enhances waste biotransformation. Waste Manag. 2025, 204, 114886. [Google Scholar] [CrossRef]
- Erickson, M.C.; Islam, M.; Sheppard, C.; Liao, J.; Doyle, M.P. Reduction of Escherichia coli O157:H7 and Salmonella enterica serovar Enteritidis in chicken manure by larvae of the black soldier fly. J. Food Prot. 2004, 67, 1486–1490. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Tomberlin, J.K.; Brady, J.A.; Sanford, M.R.; Yu, Z. Black soldier fly (Diptera: Stratiomyidae) larvae reduce Escherichia coli in dairy manure. Environ. Entomol. 2008, 37, 1525–1530. [Google Scholar] [CrossRef] [PubMed]
- Gabler, F. Using Black Soldier Fly for Waste Recycling and Effective Salmonella spp. Reduction. Project Thesis, Swedish University of Agricultural Sciences, Uppsala, Sweden, 2014. [Google Scholar]
- Van Moll, L.; De Smet, J.; Paas, A.; Tegtmeier, D.; Vilcinskas, A.; Cos, P.; Van Campenhout, L. In vitro evaluation of antimicrobial peptides from the Black Soldier Fly (Hermetia illucens) against a selection of human pathogens. Microbiol. Spectr. 2022, 10, e01664-21. [Google Scholar] [CrossRef] [PubMed]
- Vogel, H.; Müller, A.; Heckel, D.G.; Gutzeit, H.; Vilcinskas, A. Nutritional immunology: Diversification and diet-dependent expression of antimicrobial peptides in the black soldier fly Hermetia illucens. Dev. Comp. Immunol. 2018, 78, 141–148. [Google Scholar] [CrossRef]
- Koutsos, E.; Modica, B.; Freel, T. Immunomodulatory potential of black soldier fly larvae: Applications beyond nutrition in animal feeding programs. Transl. Anim. Sci. 2022, 6, txac084. [Google Scholar] [CrossRef]
- Elhag, O.; Zhou, D.; Song, Q.; Soomro, A.A.; Cai, M.; Zheng, L.; Yu, Z.; Zhang, J. Screening, expression, purification and functional characterization of novel antimicrobial peptide genes from Hermetia illucens (L.). PLoS ONE 2017, 12, e0169582. [Google Scholar] [CrossRef]
- Bosch, J.A.; Colbeth, R.; Zirin, J.; Perrimon, N. Gene knock-ins in Drosophila using homology-independent insertion of universal donor plasmids. Genetics 2020, 214, 75–89. [Google Scholar] [CrossRef]
- Huang, Y.; Liu, Z.; Rong, Y.S. Genome editing: From Drosophila to non-model insects and beyond. J. Genet. Genom. 2016, 43, 263–272. [Google Scholar] [CrossRef]
- Xu, J.; Xu, X.; Zhan, S.; Huang, Y. Genome editing in insects: Current status and challenges. Natl. Sci. Rev. 2019, 6, 399–401. [Google Scholar] [CrossRef]
- Redman, M.; King, A.; Watson, C.; King, D. What is CRISPR/Cas9? Arch. Dis. Child.-Educ. Pract. 2016, 101, 213–215. [Google Scholar] [CrossRef]
- Sun, D.; Guo, Z.; Liu, Y.; Zhang, Y. Progress and prospects of CRISPR/Cas systems in insects and other arthropods. Front. Physiol. 2017, 8, 608. [Google Scholar] [CrossRef]
- Kou, Z.; Luo, X.; Jiang, Y.; Chen, B.; Song, Y.; Wang, Y.; Xu, J.; Tomberlin, J.K.; Huang, Y. Establishment of highly efficient transgenic system for black soldier fly (Hermetia illucens). Insect Sci. 2023, 30, 888–900. [Google Scholar] [CrossRef] [PubMed]
- Masumoto, M.; Ohde, T.; Shiomi, K.; Yaginuma, T.; Niimi, T. A Baculovirus Immediate-Early Gene, ie1, Promoter Drives Efficient Expression of a Transgene in Both Drosophila melanogaster and Bombyx mori. PLoS ONE 2012, 7, e49323. [Google Scholar] [CrossRef] [PubMed]
- Pfitzner, C.; Tepper, K.; Kumar, S.; Retief, C.; McNab, J.M.; Harrell, R.A.; Maselko, M. Rapid generation and screening of transgenic black soldier fly (Hermetia illucens). bioRxiv 2024. [Google Scholar] [CrossRef]
- Kou, Z.; Wang, S.; Luo, X.; Xu, J.; Tomberlin, J.K.; Huang, Y. Wingless strain created using binary transgenic CRISPR/Cas9 alleviates concerns about mass rearing of Hermetia illucens. Commun. Biol. 2024, 7, 1652. [Google Scholar] [CrossRef]
- Gunther, D.; Alford, R.; Johnson, J.; Neilsen, P.; Zhang, L.; Harrell, R.; Day, C. Transgenic black soldier flies for production of carotenoids. Insect Biochem. Mol. Biol. 2024, 168, 104110. [Google Scholar] [CrossRef]
- Wang, S.; Kou, Z.; Jiang, Y.; Tomberlin, J.K.; Zhang, W.; Wang, S.; Huang, Y. Truncated Higiant enhances the bioconversion ability of Hermetia illucens. Commun. Biol. 2025, 8, 1164. [Google Scholar] [CrossRef]
- Directive 2001/18/EC of the European Parliament and of the Council of 12 March 2001 on the deliberate release into the environment of genetically modified organisms and repealing Council Directive 90/220/EEC. Off. J. Eur. Communities L 2001, 106, 1–39. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32001L0018 (accessed on 22 December 2025).
- Regulation (EC) No 1829/2003 of the European Parliament and of the Council of 22 September 2003 on genetically modified food and feed. Off. J. Eur. Union L 2003, 268, 1–23. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32003R1829 (accessed on 22 December 2025).
- European Commission Fact Sheet: Questions and Answers on EU’s Policies on GMOs. 2015. Available online: https://ec.europa.eu/commission/presscorner/detail/en/memo_15_4778 (accessed on 22 December 2025).
- Food and Agriculture Organization of the United Nations; FAO. Genetically Modified Crops: Safety, Benefits, Risks and Global Status. 2022. Available online: https://openknowledge.fao.org/server/api/core/bitstreams/7364d823-4317-46c5-871c-5b36966bd482/content (accessed on 22 December 2025).
- Tepper, K.; Edwards, O.; Sunna, A.; Paulsen, I.T.; Maselko, M. Diverting organic waste from landfills via insect biomanufacturing using engineered black soldier flies (Hermetia illucens). Commun. Biol. 2024, 7, 862. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Feature | Key Findings | Industrial Application | References |
|---|---|---|---|
| Genetic origin | South America: 52 haplotypes identified globally. | Identification of lineages with desirable traits for selective breeding and maintaining genetic diversity in industrial stocks. | [32,33] |
| Genome composition | Genome size: 1102 Mb; Number of genes: 16,770; mitochondrial genome length: 15,698 bp. | Provides reference for genomic studies and development of molecular tools to monitor or manipulate BSF populations. | [37,40] |
| Affinity for organic matter | 326 genes expressed in larvae across different substrates; major genes involved in ribosome function and digestion. Gut microbiota varies between genetically distinct lines, indicating host genetics influences bioconversion efficiency. | Highlights genetic basis for substrate utilization; guides strain selection and optimisation for industrial bioconversion. | [37,46] |
| Immune system/environmental adaptation | 31 PGRPs, 16 GNBPs, 57 AMP genes including cecropins and defensins, sarcotoxin and stomoxyn genes; phenoloxidase system; antimicrobial activity against Gram-positive/negative bacteria, fungi, viruses. | Ensures larval survival and biosafety during industrial-scale by-products conversion. | [18,19,37,50,51,52,53,54,55,56] |
| Genome editing | CRISPR/Cas9 and piggyBac systems enable targeted gene modifications and creation of transgenic BSF lines. Reported advances include PTTH mutation to extend the larval stage (larger larvae), CarRA/CarB insertion to produce carotenoids, and fluorescent lines for traceability. A recent binary CRISPR/Cas9 system using germline promoters generated stable Cas9-expressing lines with normal development, enabling highly efficient germline mutagenesis when crossed with sgRNA lines (e.g., white). Targeting vestigial produced wingless, sterile adults for safe, controllable mass rearing. | Enables optimisation of growth, nutrient content, and traceability in industrial bioconversion while providing biosafety controls. | [37,62,64,65,66] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Share and Cite
Oliveira, J.; Guilgur, L.G.; Assunção, R.; Murta, D.; Trindade, A. Genomic Insights and Bioconversion Potential in the Black Soldier Fly (Hermetia illucens): Current Advances and Future Directions. Insects 2026, 17, 70. https://doi.org/10.3390/insects17010070
Oliveira J, Guilgur LG, Assunção R, Murta D, Trindade A. Genomic Insights and Bioconversion Potential in the Black Soldier Fly (Hermetia illucens): Current Advances and Future Directions. Insects. 2026; 17(1):70. https://doi.org/10.3390/insects17010070
Chicago/Turabian StyleOliveira, Joana, Leonardo Gaston Guilgur, Ricardo Assunção, Daniel Murta, and Alexandre Trindade. 2026. "Genomic Insights and Bioconversion Potential in the Black Soldier Fly (Hermetia illucens): Current Advances and Future Directions" Insects 17, no. 1: 70. https://doi.org/10.3390/insects17010070
APA StyleOliveira, J., Guilgur, L. G., Assunção, R., Murta, D., & Trindade, A. (2026). Genomic Insights and Bioconversion Potential in the Black Soldier Fly (Hermetia illucens): Current Advances and Future Directions. Insects, 17(1), 70. https://doi.org/10.3390/insects17010070