

Knocked-Out Bombyx mori Protein Disulfide Isomerase Decreases Silk Yields and Mechanical Properties by Affecting the Post-Translational Modification of Silk Proteins
 and
and 
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Silkworm Rearing and Sample Collection
2.2. Bioinformatics Analysis
2.3. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.4. sgRNA Design and Vector Construction
2.5. Embryonic Microinjection and Screening
2.6. Phenotypic and Economic Trait Analysis
2.7. Scanning Electron Microscopy of Cocoon Silk
2.8. Tensile Properties of Cocoon Silk
2.9. Fourier Transform Infrared Spectroscopy
2.10. Extraction Analysis of Silk Proteins
2.11. Sample Preparation for LC-MS/MS Detection
2.12. LC-MS/MS Analysis
2.13. Statistical Analysis
3. Results
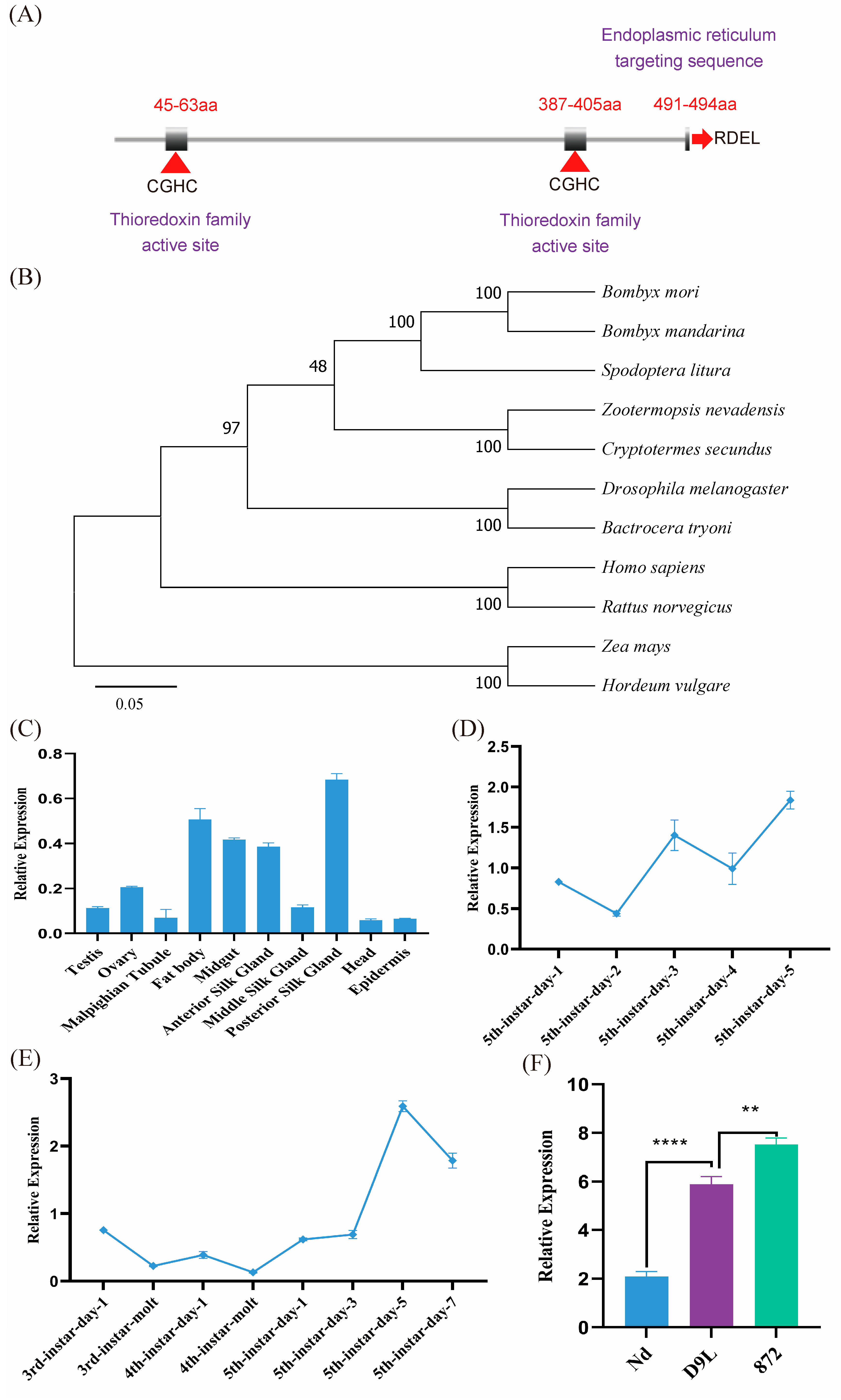
3.1. BmPDI Is Evolutionarily Conserved with the Highest Expression in PSG of Silkworm
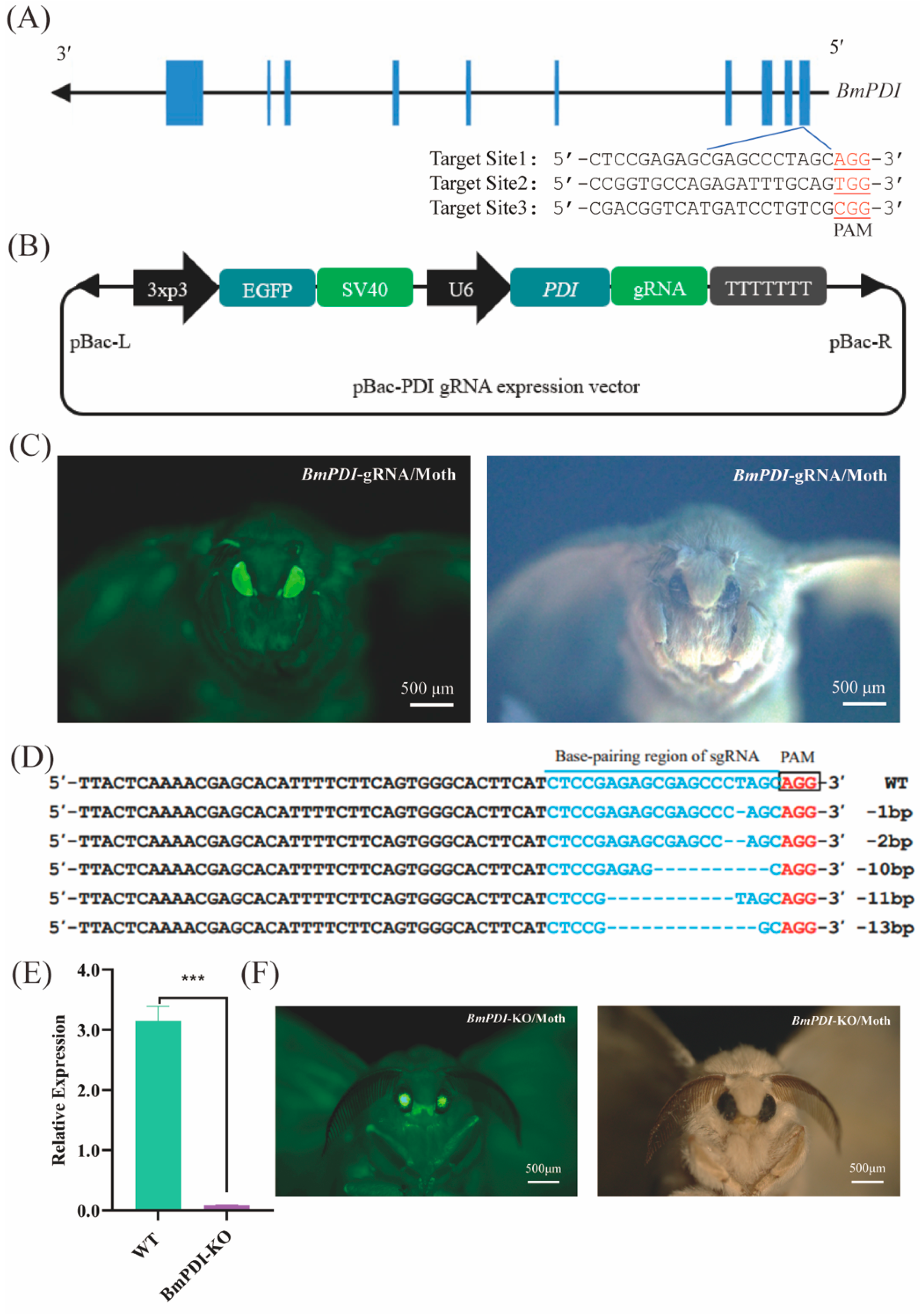
3.2. CRISPR/Cas9-Mediated BmPDI Knock-Out
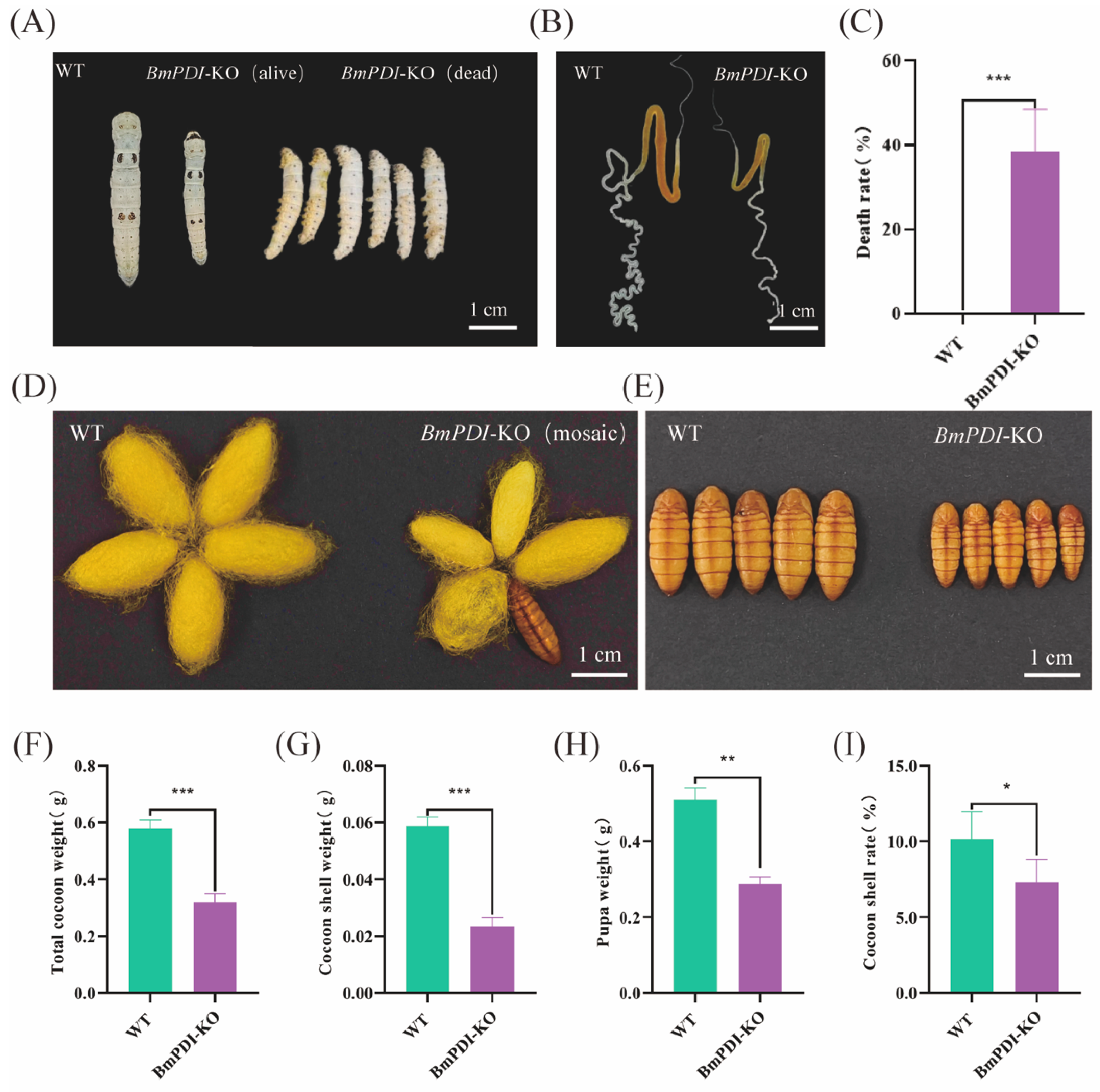
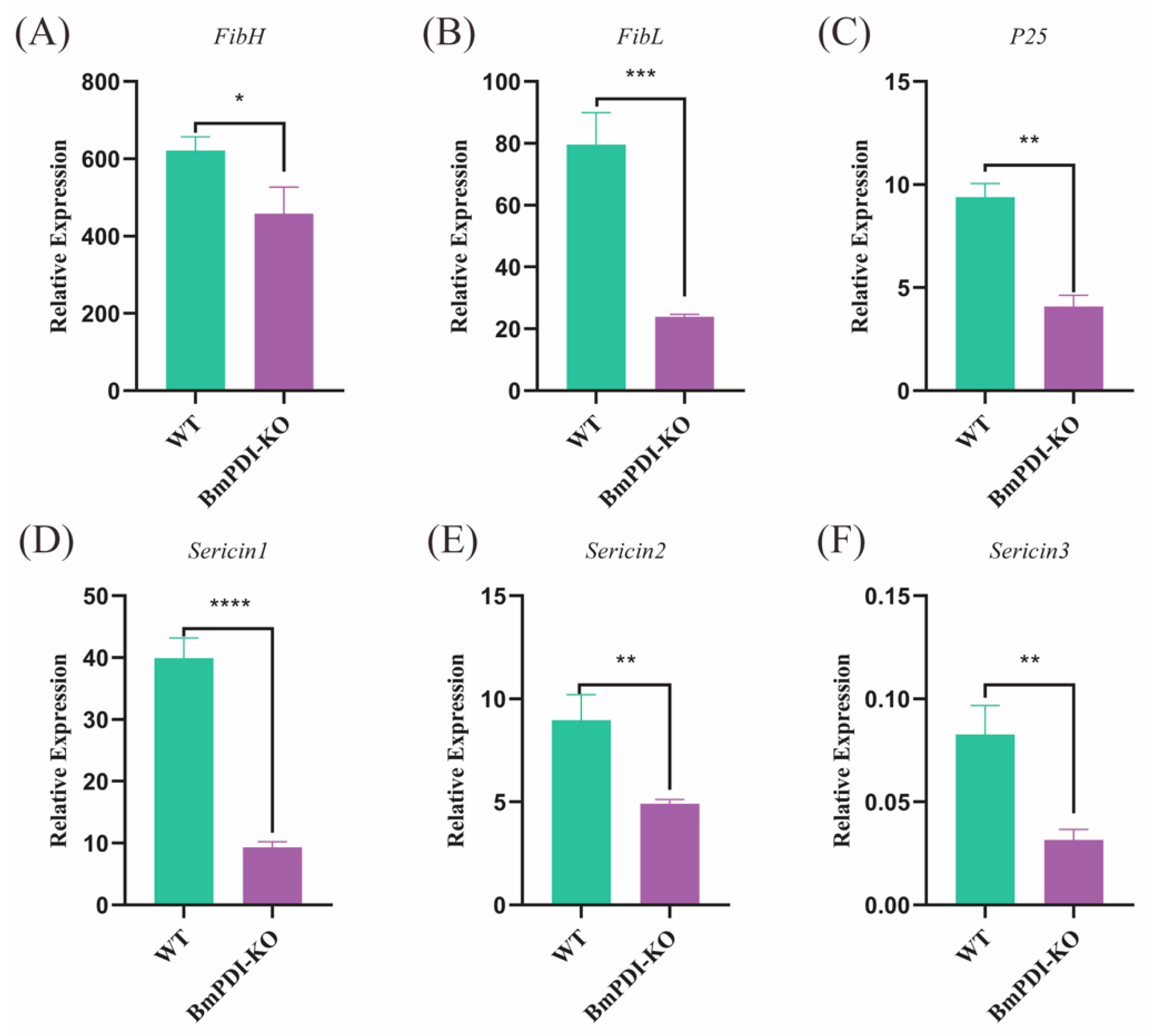
3.3. Deletion of BmPDI Inhibits the Development of Silk Glands and Decreases the Silk Yields
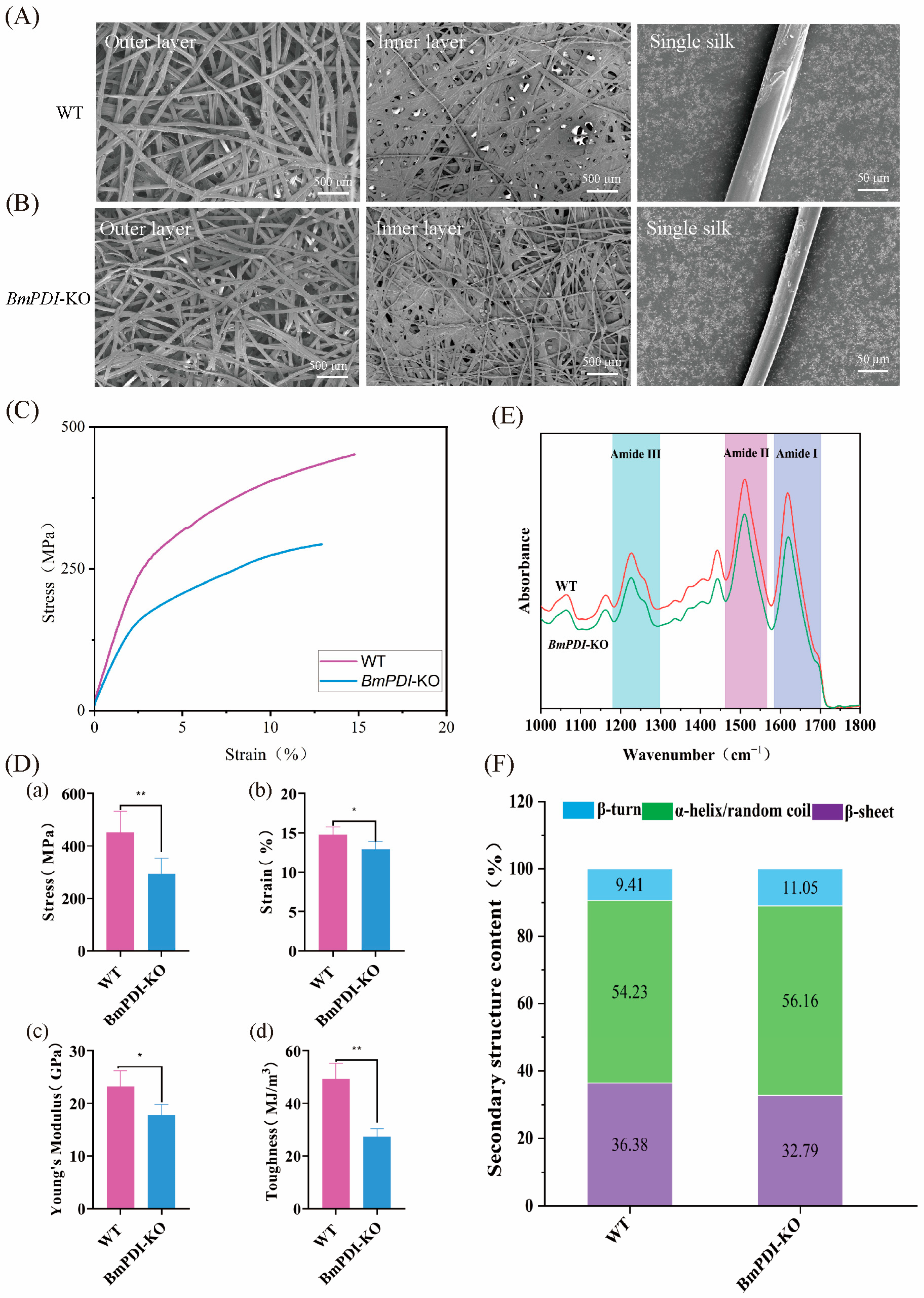
3.4. Deletion of BmPDI Reduces the β-Sheet Crystal of Silk and Decreases the Silk’s Mechanical Properties
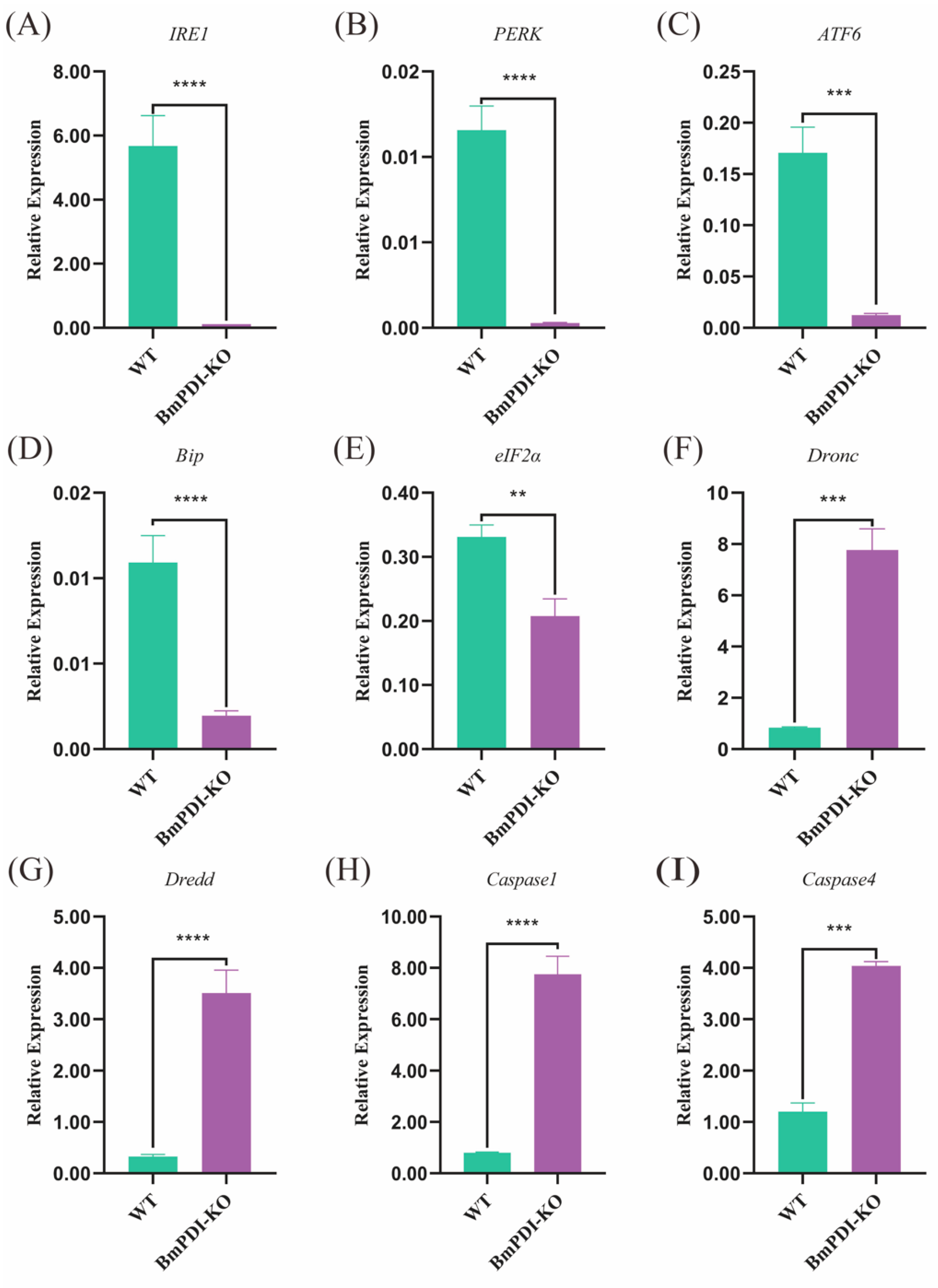
3.5. BmPDI Knock-Out Decreases the Silk Yields and Mechanical Properties by Regulating Post-Translational Modification of Silk Proteins
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| ASG | Anterior silk gland |
| MSG | Middle silk gland |
| PSG | Posterior silk gland |
| Nd | Naked pupa |
| qRT-PCR | Real-time quantitative-PCR |
| FibH | Fibroin heavy protein |
| FibL | Fibroin light protein |
| P25 | Protein 25 kDa |
| Ser1 | Sericin-1 |
| Ser2 | Sericin-2 |
| Ser3 | Sericin-3 |
| PDI | Protein disulfide isomerase |
| ER | Endoplasmic reticulum |
| ERS | Endoplasmic reticulum stress |
| UPR | Unfolded protein response |
| Bip | Immunoglobulin-binding protein |
| IRE1 | Inositol-requiring protein 1 |
| ATF6 | Activating transcription factor 6 |
| PERK | Protein kinase RNA-like ER kinase |
| eIF2α | eukaryotic translation initiation 2 |
| gRNA | guide RNA |
| PAM | Protospacer-associated motif |
| CDS | Coding sequence |
| WT | Wild type |
| aa | Amino acids |
| Dronc | Drosophila Nedd2-like caspase |
| Dredd | Death related ced-3/Nedd2-like protein |
References
- Xia, Q.; Li, S.; Feng, Q. Advances in silkworm studies accelerated by the genome sequencing of Bombyx mori. Annu. Rev. Entomol. 2014, 59, 513–536. [Google Scholar] [CrossRef] [PubMed]
- Gamo, T.; Inokuchi, T.; Laufer, H. Polypeptides of fibroin and sericin secreted from the different sections of the silk gland in Bombyx mori. Insect Biochem. 1977, 7, 285–295. [Google Scholar] [CrossRef]
- Inoue, S.; Tanaka, K.; Arisaka, F.; Kimura, S.; Ohtomo, K.; Mizuno, S. Silk fibroin of Bombyx mori is secreted, assembling a high molecular mass elementary unit consisting of H-chain, L-chain, and P25, with a 6:6:1 molar ratio. J. Biol. Chem. 2000, 275, 40517–40528. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Kajiyama, N.; Ishikura, K.; Waga, S.; Kikuchi, A.; Ohtomo, K.; Takagi, T.; Mizuno, S. Determination of the site of disulfide linkage between heavy and light chains of silk fibroin produced by Bombyx mori. Biochim. Biophys. Acta 1999, 1432, 92–103. [Google Scholar] [CrossRef]
- Takasu, Y.; Yamada, H.; Tsubouchi, K. Isolation of three main sericin components from the cocoon of the silkworm, Bombyx mori. Biosci. Biotechnol. Biochem. 2002, 66, 2715–2718. [Google Scholar] [CrossRef]
- Takasu, Y.; Hata, T.; Uchino, K.; Zhang, Q. Identification of Ser2 proteins as major sericin components in the non-cocoon silk of Bombyx mori. Insect Biochem. Mol. Biol. 2010, 40, 339–344. [Google Scholar] [CrossRef]
- Sahoo, J.K.; Hasturk, O.; Falcucci, T.; Kaplan, D.L. Silk chemistry and biomedical material designs. Nat. Rev. Chem. 2023, 7, 302–318. [Google Scholar] [CrossRef]
- Lv, Q.; Li, Q.; Cao, P.; Wei, C.; Li, Y.; Wang, Z.; Wang, L. Designing Silk Biomaterials toward Better Future Healthcare: The Development and Application of Silk-Based Implantable Electronic Devices in Clinical Diagnosis and Therapy. Adv. Mater. 2024, 37, e2411946. [Google Scholar] [CrossRef]
- Wang, F.; Guo, C.; Yang, Q.; Li, C.; Zhao, P.; Xia, Q.; Kaplan, D.L. Protein composites from silkworm cocoons as versatile biomaterials. Acta Biomater. 2021, 121, 180–192. [Google Scholar] [CrossRef]
- Mauney, J.R.; Nguyen, T.; Gillen, K.; Kirker-Head, C.; Gimble, J.M.; Kaplan, D.L. Engineering adipose-like tissue in vitro and in vivo utilizing human bone marrow and adipose-derived mesenchymal stem cells with silk fibroin 3D scaffolds. Biomaterials 2007, 28, 5280–5290. [Google Scholar] [CrossRef]
- Lawrence, B.D.; Marchant, J.K.; Pindrus, M.A.; Omenetto, F.G.; Kaplan, D.L. Silk film biomaterials for cornea tissue engineering. Biomaterials 2009, 30, 1299–1308. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Lei, H.; Tian, C.; Ji, Y.; Wang, F.; Deng, H.; Zhou, H.; Chen, S.; Zhou, Y.; Meng, Z.; et al. An Efficient Biosynthetic System for Developing Functional Silk Fibroin-Based Biomaterials. Adv. Mater. 2024, 37, e2414878. [Google Scholar] [CrossRef] [PubMed]
- Aramwit, P.; Damrongsakkul, S.; Kanokpanont, S.; Srichana, T. Properties and antityrosinase activity of sericin from various extraction methods. Biotechnol. Appl. Biochem. 2010, 55, 91–98. [Google Scholar] [CrossRef]
- Manosroi, A.; Boonpisuttinant, K.; Winitchai, S.; Manosroi, W.; Manosroi, J. Free radical scavenging and tyrosinase inhibition activity of oils and sericin extracted from Thai native silkworms (Bombyx mori). Pharm. Biol. 2010, 48, 855–860. [Google Scholar] [CrossRef]
- Wang, H.Y.; Wang, Y.J.; Zhou, L.X.; Zhu, L.; Zhang, Y.Q. Isolation and bioactivities of a non-sericin component from cocoon shell silk sericin of the silkworm Bombyx mori. Food Funct. 2012, 3, 150–158. [Google Scholar] [CrossRef]
- Wang, F.; Ning, A.F.; Sun, X.C.; Zhou, Y.J.; Deng, H.X.; Zhou, H.J.; Chen, S.Y.; He, M.Y.; Meng, Z.H.; Wang, Y.S.; et al. Fabrication of a transforming growth factor β1 functionalized silk sericin hydrogel through genetical engineering to repair alveolar bone defects in rabbit. Biomaterials 2025, 316, 122986. [Google Scholar] [CrossRef]
- Wilkinson, B.; Gilbert, H.F. Protein disulfide isomerase. Biochim. Biophys. Acta 2004, 1699, 35–44. [Google Scholar] [CrossRef]
- Tian, G.; Xiang, S.; Noiva, R.; Lennarz, W.J.; Schindelin, H. The crystal structure of yeast protein disulfide isomerase suggests cooperativity between its active sites. Cell 2006, 124, 61–73. [Google Scholar] [CrossRef]
- Xu, S.; Sankar, S.; Neamati, N. Protein disulfide isomerase: A promising target for cancer therapy. Drug Discov. Today 2014, 19, 222–240. [Google Scholar] [CrossRef]
- Matsusaki, M.; Kanemura, S.; Kinoshita, M.; Lee, Y.H.; Inaba, K.; Okumura, M. The Protein Disulfide Isomerase Family: From proteostasis to pathogenesis. Biochim. Biophys. Acta Gen. Subj. 2020, 1864, 129338. [Google Scholar] [CrossRef]
- Bilches Medinas, D.; Malik, S.; Yıldız-Bölükbaşı, E.; Borgonovo, J.; Saaranen, M.J.; Urra, H.; Pulgar, E.; Afzal, M.; Contreras, D.; Wright, M.T.; et al. Mutation in protein disulfide isomerase A3 causes neurodevelopmental defects by disturbing endoplasmic reticulum proteostasis. EMBO J. 2022, 41, e105531. [Google Scholar] [CrossRef] [PubMed]
- Goo, T.W.; Yun, E.Y.; Hwang, J.S.; Kang, S.W.; Park, S.; You, K.H.; Kwon, O.Y. Molecular characterization of a Bombyx mori protein disulfide isomerase (bPDI). Cell Stress Chaperones 2002, 7, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ma, S.; Chang, J.; Zhang, T.; Wang, X.; Shi, R.; Zhang, J.; Lu, W.; Liu, Y.; Xia, Q. Tissue-specific genome editing of laminA/C in the posterior silk glands of Bombyx mori. J. Genet. Genom. 2017, 44, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wang, R.Y.; Zhong, D.B.; Zhao, P.; Xia, Q.Y. Highly efficient expression of human extracellular superoxide dismutase (rhEcSOD) with ultraviolet-B-induced damage-resistance activity in transgenic silkworm cocoons. Insect Sci. 2023, 31, 1150–1164. [Google Scholar] [CrossRef]
- Zhang, X.; Dong, Z.; Guo, K.; Jiang, W.; Wu, X.; Duan, J.; Jing, X.; Xia, Q.; Zhao, P. Identification and functional study of fhx-L1, a major silk component in Bombyx mori. Int. J. Biol. Macromol. 2023, 232, 123371. [Google Scholar] [CrossRef]
- Liu, Y.; Ma, S.; Wang, X.; Chang, J.; Gao, J.; Shi, R.; Zhang, J.; Lu, W.; Liu, Y.; Zhao, P.; et al. Highly efficient multiplex targeted mutagenesis and genomic structure variation in Bombyx mori cells using CRISPR/Cas9. Insect Biochem. Mol. Biol. 2014, 49, 35–42. [Google Scholar] [CrossRef]
- Peng, Z.; Yang, X.; Liu, C.; Dong, Z.; Wang, F.; Wang, X.; Hu, W.; Zhang, X.; Zhao, P.; Xia, Q. Structural and Mechanical Properties of Silk from Different Instars of Bombyx mori. Biomacromolecules 2019, 20, 1203–1216. [Google Scholar] [CrossRef]
- Boulet-Audet, M.; Vollrath, F.; Holland, C. Identification and classification of silks using infrared spectroscopy. J. Exp. Biol. 2015, 218, 3138–3149. [Google Scholar] [CrossRef]
- Fang, G.; Sapru, S.; Behera, S.; Yao, J.; Shao, Z.; Kundu, S.C.; Chen, X. Exploration of the tight structural-mechanical relationship in mulberry and non-mulberry silkworm silks. J. Mater. Chem. B 2016, 4, 4337–4347. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, X.; Tan, X.; Xie, X.; Li, Y.; Zhao, P.; Xi, Q. A strategy for improving the mechanical properties of silk fiber by directly injection of ferric ions into silkworm. Mater. Des. 2018, 146, 134–141. [Google Scholar] [CrossRef]
- Sun, M.; Zhang, Y.; Zhao, Y.; Shao, H.; Hu, X. The structure–property relationships of artificial silk fabricated by dry-spinning process. J. Mater. Chem. 2012, 22, 18372. [Google Scholar] [CrossRef]
- Nguyen, A.T.; Huang, Q.L.; Yang, Z.; Lin, N.; Xu, G.; Liu, X.Y. Crystal networks in silk fibrous materials: From hierarchical structure to ultra performance. Small 2015, 11, 1039–1054. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Zhang, J.; Wang, X.; Nguyen, A.T.; Liu, X.Y.; Kaplan, D.L. Comparative Study of Strain-Dependent Structural Changes of Silkworm Silks: Insight into the Structural Origin of Strain-Stiffening. Small 2017, 13, 1702266. [Google Scholar] [CrossRef]
- Puig, A.; Gilbert, H.F. Protein disulfide isomerase exhibits chaperone and anti-chaperone activity in the oxidative refolding of lysozyme. J. Biol. Chem. 1994, 269, 7764–7771. [Google Scholar] [CrossRef]
- Ron, D.; Walter, P. Signal integration in the endoplasmic reticulum unfolded protein response. Nat. Rev. Mol. Cell Biol. 2007, 8, 519–529. [Google Scholar] [CrossRef]
- Gamo, T. Ultrastructural study of the posterior silkgland in the Nd, Nd-s and Nd-sD mutants with a defect of fibroin synthesis. J. Sericult. Sci. Jpn. 1985, 54, 412–419. [Google Scholar]
- Mori, K.; Tanaka, K.; Kikuchi, Y.; Waga, M.; Waga, S.; Mizuno, S. Production of a chimeric fibroin light-chain polypeptide in a fibroin secretion-deficient naked pupa mutant of the silkworm Bombyx mori. J. Mol. Biol. 1995, 251, 217–228. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, P.; Li, Y.; Yi, Q.; Ma, S.; Xie, K.; Chen, H.; Xia, Q. Modifying the Mechanical Properties of Silk Fiber by Genetically Disrupting the Ionic Environment for Silk Formation. Biomacromolecules 2015, 16, 3119–3125. [Google Scholar] [CrossRef]
- Guo, C.; Zhang, J.; Jordan, J.S.; Wang, X.; Henning, R.W.; Yarger, J.L. Structural Comparison of Various Silkworm Silks: An Insight into the Structure-Property Relationship. Biomacromolecules 2018, 19, 906–917. [Google Scholar] [CrossRef]
- Cheng, T.; Zhang, X.; Peng, Z.; Fan, Y.; Zhang, L.; Liu, C. Effects of Osiris9a on Silk Properties in Bombyx mori Determined by Transgenic Overexpression. Int. J. Mol. Sci. 2020, 21, 1888. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bombyx mori Line | G0 Eggs Microinjected | G0 Eggs Hatched | G1 Moth Broods | Positive G1 Moth Broods | Positive Ratio |
|---|---|---|---|---|---|
| D9L | 300 | 57 | 20 | 2 | 10% |
| Category Name | Number of Disulfide-Linked Peptides in WT Lines | Number of Disulfide-Linked Peptides in BmPDI-KO Lines |
|---|---|---|
| FibH protein | 1 | 0 |
| FibL protein | 56 | 41 |
| P25 protein | 70 | 53 |
| Total | 127 | 94 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, S.; He, M.; Li, X.; Dong, H.; Lei, H.; Wang, F.; Deng, H.; Zhou, H.; Chen, S.; Zhou, Y.; et al. Knocked-Out Bombyx mori Protein Disulfide Isomerase Decreases Silk Yields and Mechanical Properties by Affecting the Post-Translational Modification of Silk Proteins. Insects 2025, 16, 684. https://doi.org/10.3390/insects16070684
Yang S, He M, Li X, Dong H, Lei H, Wang F, Deng H, Zhou H, Chen S, Zhou Y, et al. Knocked-Out Bombyx mori Protein Disulfide Isomerase Decreases Silk Yields and Mechanical Properties by Affecting the Post-Translational Modification of Silk Proteins. Insects. 2025; 16(7):684. https://doi.org/10.3390/insects16070684
Chicago/Turabian StyleYang, Shifeng, Mengyao He, Xian Li, Huan Dong, Hexu Lei, Fangyu Wang, Hanxin Deng, Hongji Zhou, Siyu Chen, Yujuan Zhou, and et al. 2025. "Knocked-Out Bombyx mori Protein Disulfide Isomerase Decreases Silk Yields and Mechanical Properties by Affecting the Post-Translational Modification of Silk Proteins" Insects 16, no. 7: 684. https://doi.org/10.3390/insects16070684
APA StyleYang, S., He, M., Li, X., Dong, H., Lei, H., Wang, F., Deng, H., Zhou, H., Chen, S., Zhou, Y., Meng, Z., Tu, D., Wang, H., Xia, Q., & Wang, F. (2025). Knocked-Out Bombyx mori Protein Disulfide Isomerase Decreases Silk Yields and Mechanical Properties by Affecting the Post-Translational Modification of Silk Proteins. Insects, 16(7), 684. https://doi.org/10.3390/insects16070684