
Sublethal Effects of Abamectin and Acetamiprid on the Longevity, Fecundity and Detoxification Enzyme Activity of Rhopalosiphum padi


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Insecticides and Reagents
2.3. Bioassay
2.4. Sublethal Effects on the Parent Generation of R. padi
2.5. Sublethal Effects on the Traits of the Offspring of R. padi
2.6. Age-Specific Life Table and Population Projection
2.7. Determination of Detoxification Enzyme Activities in R. padi
2.8. Data Analysis
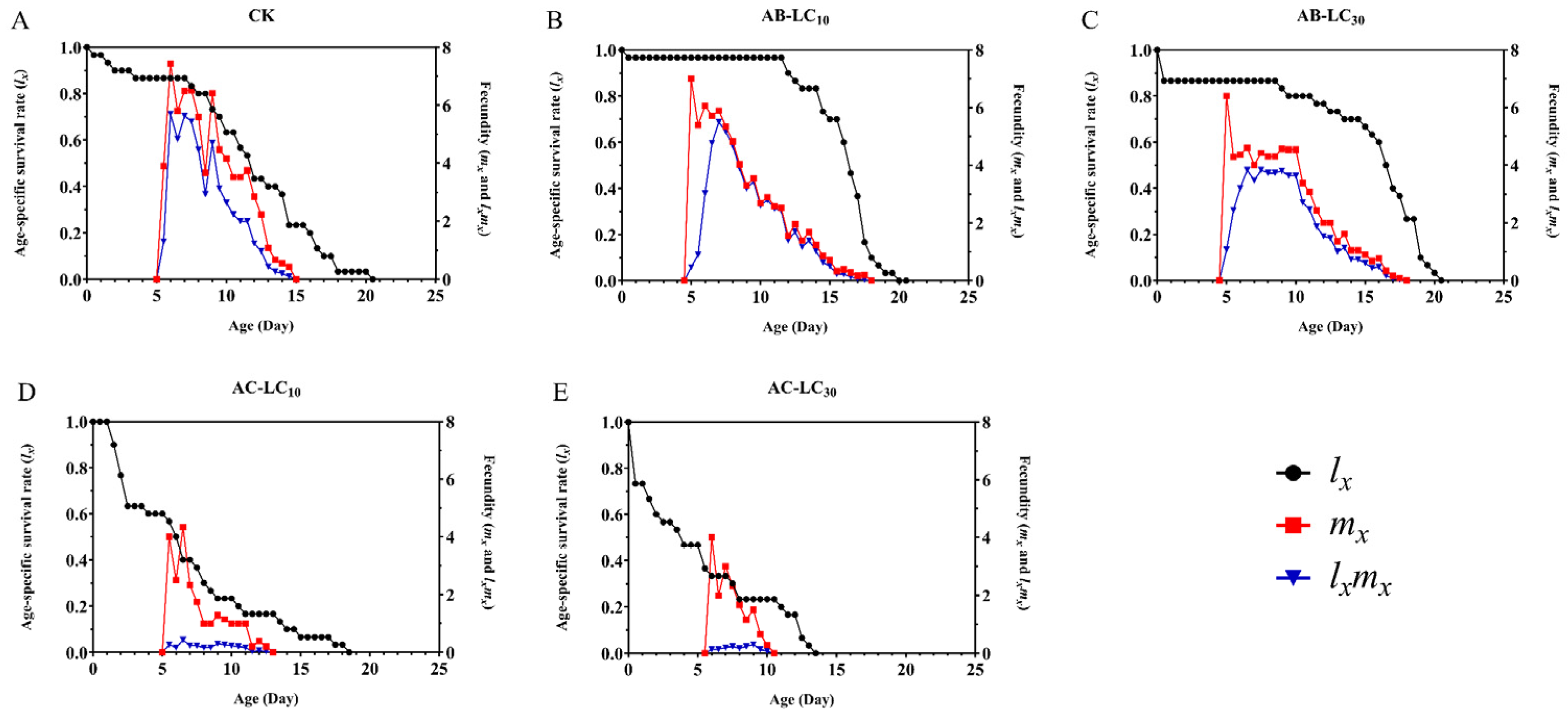
3. Results
3.1. Determination of the Toxicity of Abamectin and Acetamiprid to R. padi
3.2. Sublethal Effects on the Survial, Longevity and Fecundity of the F0 Generation of R. padi
3.3. Sublethal Effects on the Development and Fecundity of the F1 Generation of R. padi
3.4. Sublethal Effects on the Population Parameters of the R. padi F1 Generation
3.5. Population Projection of R. padi Exposure to Abamectin and Acetamiprid
3.6. Detoxifying Enzymes Activity of the F0 Generation in R. padi
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Deng, D.; Duan, W.; Wang, H.; Zhang, K.; Guo, J.; Yuan, L.; Wang, L.; Wu, S. Assessment of the effects of lethal and sublethal exposure to dinotefuran on the wheat aphid Rhopalosiphum padi (Linnaeus). Ecotoxicology 2019, 28, 825–833. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Lu, H.; Guo, K.; Yao, S.; Cui, F. Insecticide resistance status and detoxification enzymes of wheat aphids Sitobion avenae and Rhopalosiphum padi. Sci. China Life Sci. 2017, 60, 927–930. [Google Scholar] [CrossRef] [PubMed]
- Ministry of Agriculture and Rural Affairs of the People’s Republic of China (MARAC). Announcement No. 333 of the Ministry of Agriculture and Rural Affairs of the People’s Republic of China; MARAC: Beijing, China, 2020. Available online: https://www.moa.gov.cn/nybgb/2020/202010/202011/t20201130_6357326.htm (accessed on 12 October 2024).
- Miao, J.; Guo, P.; Zhang, Y.; Tan, X.; Chen, J.; Li, Y.; Wu, Y. Effect of High Temperature and Natural Enemies on the Interspecies Competition Between Two Wheat Aphid Species, Rhopalosiphum padi and Sitobion miscanthi. J. Econ. Entomol. 2022, 115, 539–544. [Google Scholar] [CrossRef]
- Gong, P.; Li, X.; Gao, H.; Wang, C.; Li, M.; Zhang, Y.; Li, X.; Liu, E.; Zhu, X. Field evolved resistance to pyrethroids, neonicotinoids, organophosphates and macrolides in Rhopalosiphum padi (Linnaeus) and Sitobion avenae (Fabricius) from China. Chemosphere 2021, 269, 128747. [Google Scholar] [CrossRef] [PubMed]
- Gul, H.; Haq, I.U.; Güncan, A.; Abbas, A.; Khan, S.; Yaseen, A.; Ullah, F.; Desneux, N.; Liu, X. Thiamethoxam-Induced Intergenerational Sublethal Effects on the Life History and Feeding Behavior of Rhopalosiphum padi. Plants 2024, 13, 865. [Google Scholar] [CrossRef]
- Cui, L.; Wang, G.; Yang, D.; Nahiyoon, S.A.; Yan, X.; Yuan, H. Biocidal radiuses of abamectin, thiamethoxam and sulfoxaflor droplets controlling against wheat aphid (Sitobion avenae). PLoS ONE 2018, 13, e0205598. [Google Scholar] [CrossRef]
- Xu, T.; Zhang, S.; Liu, Y.; Ma, L.; Li, X.; Zhang, Y.; Fan, Y.; Song, D.; Gao, X. Slow resistance evolution to neonicotinoids in field populations of wheat aphids revealed by insecticide resistance monitoring in China. Pest Manag. Sci. 2022, 78, 1428–1437. [Google Scholar] [CrossRef]
- Zhang, A.; Xu, L.; Liu, Z.; Zhang, J.; Zhao, K.; Han, L. Effects of Acetamiprid at Low and Median Lethal Concentrations on the Development and Reproduction of the Soybean Aphid Aphis glycines. Insects 2022, 13, 87. [Google Scholar] [CrossRef] [PubMed]
- Desneux, N.; Decourtye, A.; Delpuech, J.M. The sublethal effects of pesticides on beneficial arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef]
- Guedes, R.N.C.; Walse, S.S.; Throne, J.E. Sublethal exposure, insecticide resistance, and community stress. Curr. Opin. Insect Sci. 2017, 21, 47–53. [Google Scholar] [CrossRef]
- Qu, Y.; Ullah, F.; Luo, C.; Monticelli, L.S.; Lavoir, A.-V.; Gao, X.; Song, D.; Desneux, N. Sublethal effects of beta-cypermethrin modulate interspecific interactions between specialist and generalist aphid species on soybean. Ecotox. Environ. Saf. 2020, 206, 111302. [Google Scholar] [CrossRef] [PubMed]
- Iftikhar, A.; Hafeez, F.; Aziz, M.A.; Hashim, M.; Naeem, A.; Yousaf, H.K.; Saleem, M.J.; Hussain, S.; Hafeez, M.; Ali, Q.; et al. Assessment of sublethal and transgenerational effects of spirotetramat, on population growth of cabbage aphid, Brevicoryne brassicae L. (Hemiptera: Aphididae). Front. Physiol. 2022, 13, 1014190. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Fu, Z.; Zhu, Y.; Gao, X.; Liu, T.-X.; Liang, P. Sublethal and transgenerational effects of afidopyropen on biological traits of the green peach aphid Myzus persicae (Sluzer). Pestic. Biochem. Physiol. 2022, 180, 104981. [Google Scholar] [CrossRef]
- Daniels, M.; Bale, J.S.; Newbury, H.J.; Lind, R.J.; Pritchard, J. A sublethal dose of thiamethoxam causes a reduction in xylem feeding by the bird cherry-oat aphid (Rhopalosiphum padi), which is associated with dehydration and reduced performance. J. Insect Physiol. 2009, 55, 758–765. [Google Scholar] [CrossRef]
- Li, X.; Zhang, W.; Zhang, H.; Zhang, X.; Song, C.; Zhang, P.; Li, G.; Zhu, X.; Zhang, B. The sublethal concentration of acetamiprid suppresses the population growth of 2 species of wheat aphids, Sitobion miscanthi and Schizaphis graminum (Hemiptera: Aphididae). J. Econ. Entomol. 2024, 117, 1315–1323. [Google Scholar] [CrossRef]
- Ayyanath, M.M.; Cutler, G.C.; Scott-Dupree, C.D.; Sibley, P.K. Transgenerational Shifts in Reproduction Hormesis in Green Peach Aphid Exposed to Low Concentrations of Imidacloprid. PLoS ONE 2013, 8, e74532. [Google Scholar] [CrossRef]
- Huangfu, N.; Guo, L.; Shang, J.; Wang, L.; Zhang, K.; Li, D.; Gao, X.; Zhu, X.; Ji, J.; Luo, J.; et al. Hormetic dose response induced by sublethal-dose sulfoxaflor leads to reproductive stimulation of Aphis gossypii. Pestic. Biochem. Physiol. 2024, 204, 106061. [Google Scholar] [CrossRef] [PubMed]
- Ullah, F.; Gul, H.; Desneux, N.; Gao, X.; Song, D. Imidacloprid-induced hormesis effects on demographic traits of the melon aphid, Aphis gossypii. Entomol. Gen. 2019, 39, 325–337. [Google Scholar] [CrossRef]
- Ullah, F.; Gul, H.; Desneux, N.; Qu, Y.; Xiao, X.; Khattak, A.M.; Gao, X.; Song, D. Acetamiprid-induced hormetic effects and vitellogenin gene (Vg) expression in the melon aphid, Aphis gossypii. Entomol. Gen. 2019, 39, 259–270. [Google Scholar] [CrossRef]
- Ullah, F.; Gul, H.; Tariq, K.; Desneux, N.; Gao, X.; Song, D. Thiamethoxam induces transgenerational hormesis effects and alteration of genes expression in Aphis gossypii. Pestic. Biochem. Physiol. 2020, 165, 104557. [Google Scholar] [CrossRef]
- Yu, Y.; Shen, G.; Zhu, H.; Lu, Y. Imidacloprid-induced hormesis on the fecundity and juvenile hormone levels of the green peach aphid Myzus persicae (Sulzer). Pestic. Biochem. Physiol. 2010, 98, 238–242. [Google Scholar] [CrossRef]
- Tang, Q.; Ma, K.; Chi, H.; Hou, Y.; Gao, X. Transgenerational hormetic effects of sublethal dose of flupyradifurone on the green peach aphid, Myzus persicae (Sulzer) (Hemiptera: Aphididae). PLoS ONE 2019, 14, e0208058. [Google Scholar] [CrossRef]
- Fouad, E.A.; El-Sherif, S.A.N.; Mokbel, E.-S.M.S. Flupyradifurone induces transgenerational hormesis effects in the cowpea aphid, Aphis craccivora. Ecotoxicology 2022, 31, 909–918. [Google Scholar] [CrossRef]
- Gul, H.; Haq, I.U.; Ullah, F.; Khan, S.; Yaseen, A.; Tariq, K.; Güncan, A.; Desneux, N.; Liu, X. Hormetic effects of thiamethoxam on Schizaphis graminum: Demographics and feeding behavior. Ecotoxicology 2024, 33, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Ju, D.; Liu, Y.-X.; Liu, X.; Dewer, Y.; Mota-Sanchez, D.; Yang, X.-Q. Exposure to lambda-cyhalothrin and abamectin drives sublethal and transgenerational effects on the development and reproduction of Cydia pomonella. Ecotoxicol. Environ. Saf. 2023, 252, 114581. [Google Scholar] [CrossRef]
- Gul, H.; Guncan, A.; Ullah, F.; Desneux, N.; Liu, X. Intergenerational Sublethal Effects of Flonicamid on Cotton Aphid, Aphis gossypii: An Age-Stage, Two-Sex Life Table Study. Insects 2024, 15, 529. [Google Scholar] [CrossRef]
- Haynes, K.F. Sublethal effects of neurotoxic insecticides on insect behavior. Annu. Rev. Entomol. 1988, 33, 149–168. [Google Scholar] [CrossRef] [PubMed]
- Booth, L.H.; Wratten, S.D.; Kehrli, P. Effects of reduced rates of two insecticides on enzyme activity and mortality of an aphid and its lacewing predator. J. Econ. Entomol. 2007, 100, 11–19. [Google Scholar] [CrossRef]
- Kumrungsee, N.; Pluempanupat, W.; Koul, O.; Bullangpoti, V. Toxicity of essential oil compounds against diamondback moth, Plutella xylostella, and their impact on detoxification enzyme activities. J. Pest Sci. 2014, 87, 721–729. [Google Scholar] [CrossRef]
- Su, S.; Jian, C.; Zhang, X.; Fang, S.; Peng, X.; Pinero, J.C.; Chen, M. Sublethal Effects of Abamectin on the Development, Reproduction, Detoxification Enzyme Activity, and Related Gene Expression of the Oriental Fruit Moth (Lepidoptera: Tortricidae). J. Econ. Entomol. 2021, 114, 2430–2438. [Google Scholar] [CrossRef]
- Balakrishnan, B.; Su, S.; Zhang, C.; Chen, M. Identification and functional characterization of two sigma glutathione S-transferase genes from bird cherry-oat aphid (Hemiptera: Aphididae). J. Econ. Entomol. 2019, 112, 416–424. [Google Scholar] [CrossRef] [PubMed]
- Finney, D.J. Statisical logic in the monitoring of reactions to therapeutic drugs. Method. Inform. Med. 1971, 10, 237–245. [Google Scholar]
- Chi, H. TWOSEX-MSChart: A Computer Program for the Age-Stage, Two-Sex Life Table Analysis. 2023. Available online: http://www.faas.cn/cms/sitemanage/index.shtml?siteId=810640925913080000 (accessed on 24 August 2024).
- Huang, Y.B.; Chi, H. Life tables of Bactrocera cucurbitae (Diptera: Tephritidae): With an invalidation of the jackknife technique. J. Appl. Entomol. 2013, 137, 327–339. [Google Scholar] [CrossRef]
- Bai, B.; Zhang, S.P.; Li, Y.T.; Gao, P.; Yang, X.Q. Quercetin stimulates an accelerated burst of oviposition-based reproductive strategy in codling moth controlled by juvenile hormone signaling pathway. Sci. Total Environ. 2024, 913, 169643. [Google Scholar] [CrossRef]
- Chi, H.; Kavousi, A.; Gharekhani, G.; Atlihan, R.; Ozgokce, M.S.; Guncan, A.; Gokce, A.; Smith, C.L.; Benelli, G.; Guedes, R.N.C.; et al. Advances in theory, data analysis, and application of the age-stage, two-sex life table for demographic research, biological control, and pest management. Entomol. Gen. 2023, 43, 705–732. [Google Scholar] [CrossRef]
- Hu, C.; Liu, Y.X.; Zhang, S.-P.; Wang, Y.Q.; Gao, P.; Li, Y.T.; Yang, X.Q. Transcription Factor AhR Regulates Glutathione S-Transferases Conferring Resistance to lambda-Cyhalothrin in Cydia pomonella. J. Agric. Food Chem. 2023, 71, 5230–5239. [Google Scholar] [CrossRef]
- Gandara, L.; Jacoby, R.; Laurent, F.; Spatuzzi, M.; Vlachopoulos, N.; Borst, N.O.; Ekmen, G.; Potel, C.M.; Garrido-Rodriguez, M.; Boehmert, A.L.; et al. Pervasive sublethal effects of agrochemicals on insects at environmentally relevant concentrations. Science 2024, 386, 446–453. [Google Scholar] [CrossRef]
- Calabrese, E.J.; Baldwin, L.A. Hormesis: The dose-response revolution. Annu. Rev. Pharmacol. Toxicol. 2003, 43, 175–197. [Google Scholar] [CrossRef]
- Gong, Y.; Cheng, S.; Desneux, N.; Gao, X.; Xiu, X.; Wang, F.; Hou, M. Transgenerational hormesis effects of nitenpyram on fitness and insecticide tolerance/resistance of Nilaparvata lugens. J. Pest Sci. 2023, 96, 161–180. [Google Scholar] [CrossRef]
- Müller, T.; Gesing, M.A.; Segeler, M.; Müller, C. Sublethal insecticide exposure of an herbivore alters the response of its predator. Environ. Pollut. 2019, 247, 39–45. [Google Scholar] [CrossRef]
- Lashkari, M.R.; Sahragard, A.; Ghadamyari, M. Sublethal effects of imidacloprid and pymetrozine on population growth parameters of cabbage aphid, Brevicoryne brassicae on rapeseed, Brassica napus L. Insect Sci. 2007, 14, 207–212. [Google Scholar] [CrossRef]
- Ibrahim, Y.B.; Yee, T.S. Influence of sublethal exposure to abamectin on the biological performance of Neoseiulus longispinosus (Acari: Phytoseiidae). J. Econ. Entomol. 2000, 93, 1085–1089. [Google Scholar] [CrossRef] [PubMed]
- Bostanian, N.J.; Akalach, M. The effect of indoxacarb and five other insecticides on Phytoseiulus persimilis (Acari: Phytoseiidae), Amblyseius fallacis (Acari: Phytoseiidae) and nymphs of Orius insidiosus (Hemiptera: Anthocoridae). Pest Manag. Sci. 2006, 62, 334–339. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Zhou, C.; Yang, X.B.; Long, G.Y.; Jin, D.C. Effects of Insecticide Stress on Expression of NlABCG Transporter Gene in the Brown Planthopper, Nilaparvata lugens. Insects 2019, 10, 334. [Google Scholar] [CrossRef]
- Yin, X.H.; Wu, Q.J.; Li, X.F.; Zhang, Y.J.; Xu, B.Y. Sublethal effects of spinosad on Plutella xylostella (Lepidoptera: Yponomeutidae). Crop Prot. 2008, 27, 1385–1391. [Google Scholar] [CrossRef]
- Xiao, D.; Yang, T.; Desneux, N.; Han, P.; Gao, X. Assessment of Sublethal and Transgenerational Effects of Pirimicarb on the Wheat Aphids Rhopalosiphum padi and Sitobion avenae. PLoS ONE 2015, 10, e0128936. [Google Scholar] [CrossRef]
- Guedes, R.N.C.; Smagghe, G.; Stark, J.D.; Desneux, N. Pesticide-Induced Stress in Arthropod Pests for Optimized Integrated Pest Management Programs. Annu. Rev. Entomol. 2016, 61, 43–62. [Google Scholar] [CrossRef]
- Li, X.; Li, Y.; Zhu, X.; Li, X.; Cheng, D.; Zhang, Y. Effects of acetamiprid-induced hormesis on the development and reproduction of the rose-grain aphid Metopolophium dirhodum (Hemiptera: Aphididae). Front. Physiol. 2023, 14, 1113464. [Google Scholar] [CrossRef]
- Jiang, M.; Qian, X.; Zhou, Z.; Liu, Y.; Zhang, M.; Yang, Y. Impacts of Sublethal Doses of Spinetoram on the Biological Traits and Detoxifying Enzymes of the Tomato Leaf Miner, Tuta absoluta (Lepidoptera: Gelechiidae). Insects 2024, 15, 990. [Google Scholar] [CrossRef]
- Amezian, D.; Nauen, R.; Le Goff, G. Transcriptional regulation of xenobiotic detoxification genes in insects-An overview. Pestic. Biochem. Physiol. 2021, 174, 104822. [Google Scholar] [CrossRef]
- Zhang, L.; Wu, H.Z.; He, P.Y.; Cao, H.Q.; Zhang, W.N.; Peng, Y.C.; Sheng, C.W. Sublethal effect and detoxifying metabolism of metaflumizone and indoxacarb on the fall armyworm, Spodoptera frugiperda. Pestic. Biochem. Physiol. 2024, 201, 105879. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.H.; Zheng, X.S.; Gao, X.W. Sublethal effects of imidacloprid on the fecundity, longevity, and enzyme activity of Sitobion avenae (Fabricius) and Rhopalosiphum padi (Linnaeus). Bull. Entomol. Res. 2016, 106, 551–559. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Insecticides | n | Slope ± SE | x2 | df | Concentration (mg/L) (95% CI) | |
|---|---|---|---|---|---|---|
| LC10 | LC30 | |||||
| abamectin | 403 | 1.259 ± 0.158 | 9.228 | 3 | 0.063 (0.034–0.101) | 0.252 (0.158–0.348) |
| acetamiprid | 417 | 1.370 ± 0.177 | 14.313 | 3 | 0.065 (0.023–0.136) | 0.293 (0.126–0.457) |
| Parameter | CK | AB-LC10 | AB-LC30 | AC-LC10 | AC-LC30 |
|---|---|---|---|---|---|
| Longevity (d) | 9.70 ± 0.88 b | 14.63 ± 1.24 a | 11.97 ± 1.45 b | 4.8 ± 0.54 c | 6.3 ± 0.69 c |
| Fecundity (nymphs/female) | 31.37 ± 3.55 a | 26.73 ± 3.63 b | 20.77 ± 3.77 c | 16.90 ± 1.45 d | 20.43 ± 1.90 c |
| Parameters | CK | AB-LC10 | AB-LC30 | AC-LC10 | AC-LC30 |
|---|---|---|---|---|---|
| N1 (d) | 1.52 ± 0.05 ab | 1.57 ± 0.05 b | 1.75 ± 0.08 a | 1.46 ± 0.05 b | 1.60 ± 0.08 b |
| N2 (d) | 1.44 ± 0.06 b | 1.62 ± 0.06 b | 1.23 ± 0.06 c | 1.89 ± 0.12 a | 1.97 ± 0.11 a |
| N3 (d) | 1.42 ± 0.05 b | 1.41 ± 0.07 b | 1.29 ± 0.06 c | 1.88 ± 0.12 a | 1.77 ± 0.17 a |
| N4 (d) | 1.48 ± 0.03 b | 1.60 ± 0.06 b | 1.40 ± 0.05 b | 2.31 ± 0.25 a | 2.57 ± 0.20 a |
| Nymph period (d) | 5.88 ± 0.07 c | 6.21 ± 0.11 b | 5.67 ± 0.10 c | 7.44 ± 0.51 a | 7.93 ± 0.52 a |
| Oviposition period (d) | 5.90 ± 0.44 b | 8.47 ± 0.27 a | 8.17 ± 0.38 a | 2.93 ± 0.76 c | 1.92 ± 0.30 c |
| Adult longevity (d) | 13.08 ± 0.68 b | 16.26 ± 0.38 a | 16.52 ± 0.6 a | 6.12 ± 1.32 c | 4.43 ± 0.69 c |
| Total longevity (d) | 11.58 ± 0.92 b | 15.73 ± 0.64 a | 14.38 ± 1.14 a | 6.82 ± 0.91 c | 5.25 ± 0.85 c |
| Fecundity (offspring per female) | 57.92 ± 3.50 a | 55.76 ± 1.24 a | 59.73 ± 1.98 a | 11.12 ± 3.59 b | 6.86 ± 1.91 b |
| Parameters | CK | AB-LC10 | AB-LC30 | AC-LC10 | AC-LC30 |
|---|---|---|---|---|---|
| Net reproductive rate (R0, nymphs) | 50.20 ± 1.92 a | 53.90 ± 2.17 a | 51.77 ± 2.01 a | 2.97 ± 1.27 b | 1.60 ± 0.67 b |
| Mean generation time (T, d) | 8.15 ± 0.14 b | 8.55 ± 0.29 a | 8.44 ± 0.17 a | 8.70 ± 1.57 a | 8.82 ± 2.25 a |
| Intrinsic rate of increase (r, d−1) | 0.48 ± 0.01 a | 0.47 ± 0.01 a | 0.47 ± 0.01 a | 0.11 ± 0.06 b | 0.04 ± 0.06 b |
| Finite rate of increase (λ, d−1) | 1.62 ± 0.01 a | 1.59 ± 0.01 a | 1.60 ± 0.01 a | 1.12 ± 0.07 b | 1.05 ± 0.06 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, B.; Hui, H.; Li, X.; Yang, X.; Li, Y. Sublethal Effects of Abamectin and Acetamiprid on the Longevity, Fecundity and Detoxification Enzyme Activity of Rhopalosiphum padi. Insects 2025, 16, 629. https://doi.org/10.3390/insects16060629
Wang B, Hui H, Li X, Yang X, Li Y. Sublethal Effects of Abamectin and Acetamiprid on the Longevity, Fecundity and Detoxification Enzyme Activity of Rhopalosiphum padi. Insects. 2025; 16(6):629. https://doi.org/10.3390/insects16060629
Chicago/Turabian StyleWang, Bokun, Hongming Hui, Xingye Li, Xueqing Yang, and Yuting Li. 2025. "Sublethal Effects of Abamectin and Acetamiprid on the Longevity, Fecundity and Detoxification Enzyme Activity of Rhopalosiphum padi" Insects 16, no. 6: 629. https://doi.org/10.3390/insects16060629
APA StyleWang, B., Hui, H., Li, X., Yang, X., & Li, Y. (2025). Sublethal Effects of Abamectin and Acetamiprid on the Longevity, Fecundity and Detoxification Enzyme Activity of Rhopalosiphum padi. Insects, 16(6), 629. https://doi.org/10.3390/insects16060629