
Effects of Orally Delivered Double-Stranded RNA of Trehalose-6-Phosphate Synthase on the Population of Frankliniella occidentalis
 , and
, and
Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. RNA Extraction and cDNA Synthesis
2.3. Synthesis of dsRNA
2.4. Bacterially Expressed dsRNAs
2.5. Effect of In Vitro-Synthesized dsFoTPS on WFTs
2.6. Construction of Life Table of WFT Population
2.7. Expression Level Analysis
2.8. Data Analysis
3. Results
3.1. Sequence Characteristics of FoTPS
3.2. Expression Pattern of FoTPS Across Different Developmental Stages
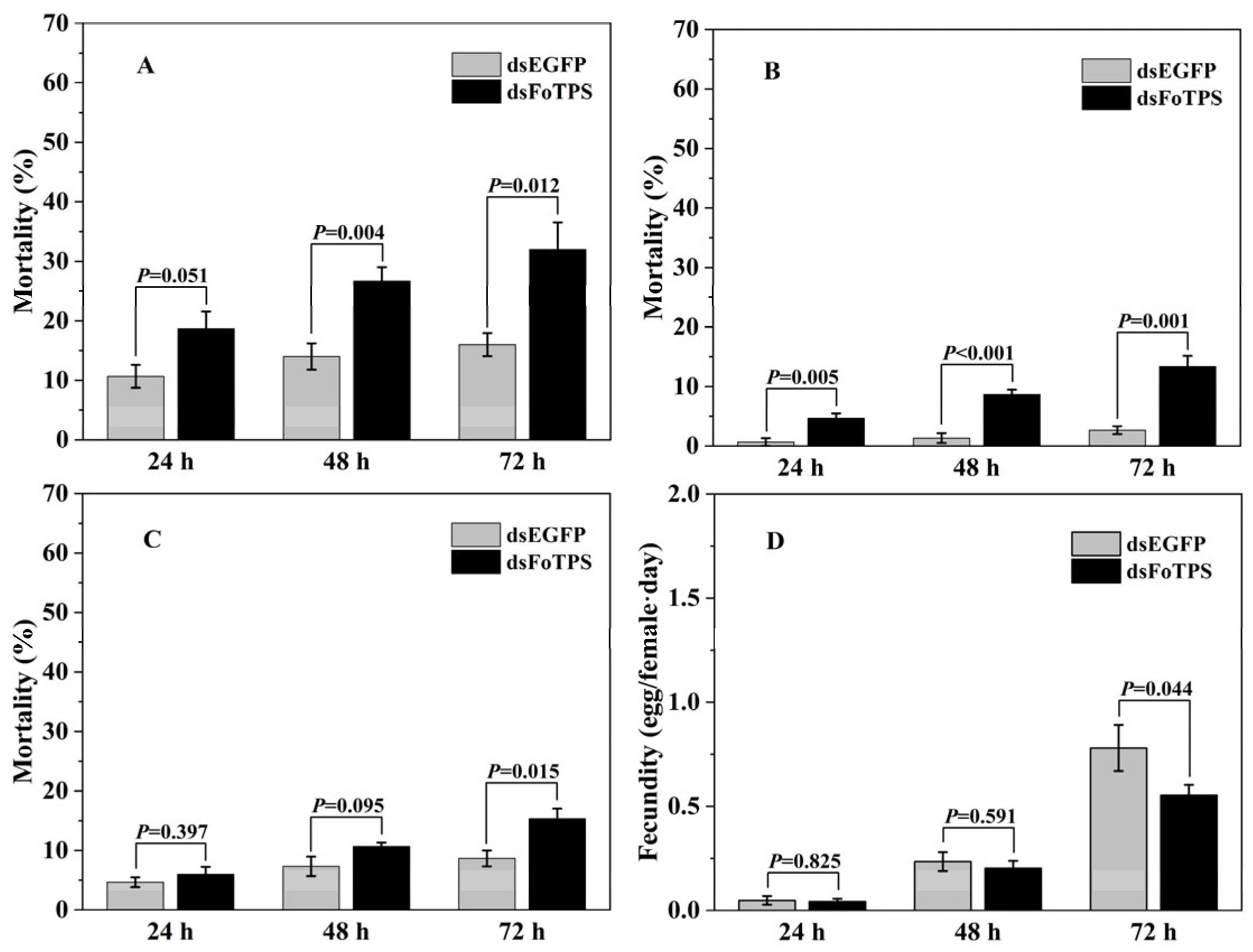
3.3. Effects of Ingestion of Synthesized dsFoTPS on Mortality and Egg-Laying Behavior of WFTs
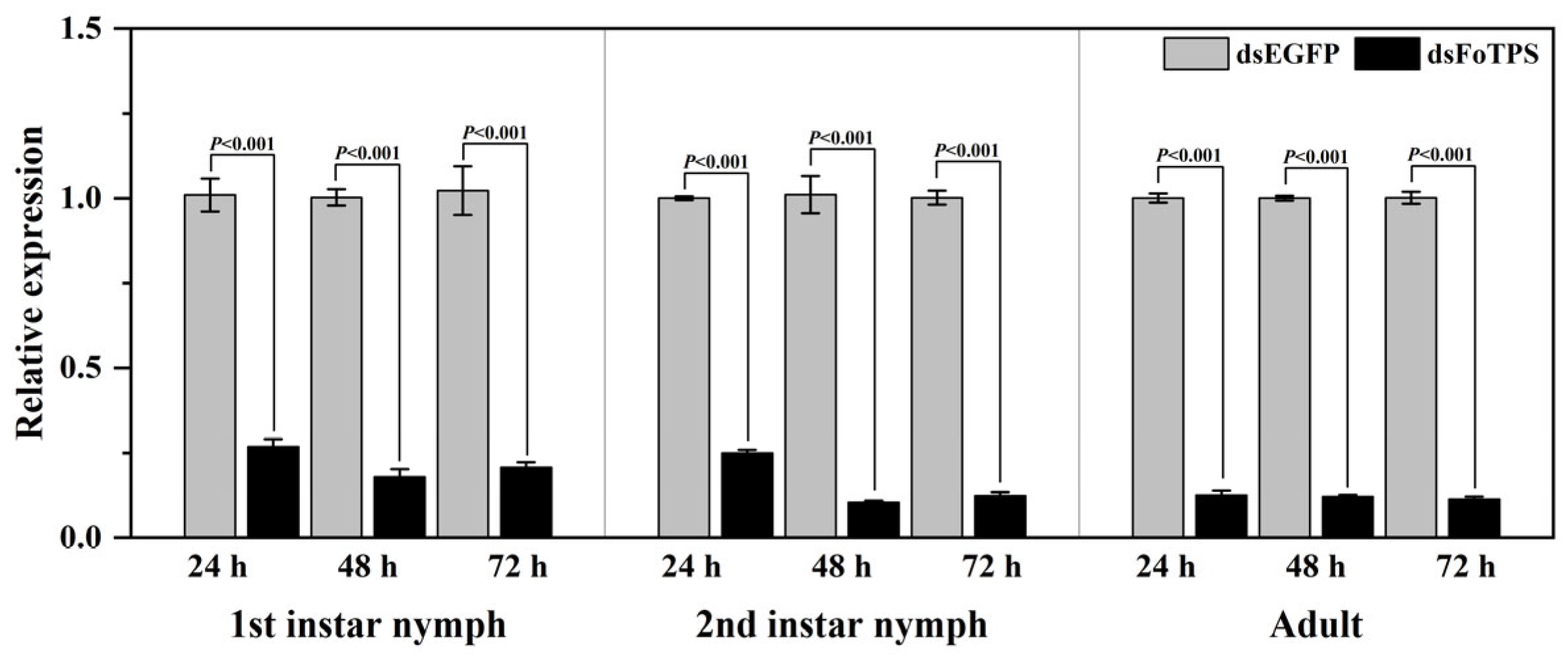
3.4. Effects of Ingestion of Synthesized dsFoTPS on Gene Expression in WFTs
3.5. Effects of E. coli Expressing dsFoTPS on Life-Table Traits of WFT Population
3.6. Effects of E. coli Expressing dsFoTPS on Population Parameters and Population Projection
3.7. Effects of Ingestion of E. coli Expressing dsFoTPS on Gene Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Reitz, S.R. Biology and ecology of the western flower thrips (Thysanoptera: Thripidae): The making of a pest. Florida Entomol. 2009, 92, 7–13. [Google Scholar] [CrossRef]
- Morse, J.G.; Hoddle, M.S. Invasion biology of thrips. Annu. Rev. Entomol. 2006, 51, 67–89. [Google Scholar] [CrossRef] [PubMed]
- Gilbertson, R.L.; Batuman, O.; Webster, C.G.; Adkins, S. Role of the insect supervectors Bemisia tabaci and Frankliniella occidentalis in the emergence and global spread of plant viruses. Annu. Rev. Virol. 2015, 2, 67–93. [Google Scholar] [CrossRef]
- Mocchetti, A.; Dermauw, W.; Van Leeuwen, T. Incidence and molecular mechanisms of insecticide resistance in Frankliniella occidentalis, Thrips tabaci and other economically important thrips species. Entomol. Gen. 2023, 43, 587–604. [Google Scholar] [CrossRef]
- Mouden, S.; Sarmiento, K.F.; Klinkhamer, P.G.L.; Leiss, K.A. Integrated pest management in western flower thrips: Past, present and future. Pest Manag. Sci. 2017, 73, 813–822. [Google Scholar] [CrossRef]
- Wu, M.T.; Dong, Y.; Zhang, Q.; Li, S.C.; Chang, L.; Loiacono, F.V.; Ruf, S.; Zhang, J.; Bock, R. Efficient control of western flower thrips by plastid-mediated RNA interference. Proc. Natl. Acad. Sci. USA 2022, 119, e2120081119. [Google Scholar] [CrossRef]
- Zhu, K.Y.; Palli, S.R. Mechanisms, applications, and challenges of insect RNA interference. Annu. Rev. Entomol. 2020, 65, 293–311. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Nauen, R.; Reitz, S.; Alyokhin, A.; Zhang, J.; Mota-sanchez, D.; Kim, Y.; Palli, S.R.; Rondon, S.I.; Nault, B.A.; et al. The new kid on the block in insect pest management: Sprayable RNAi goes commercial. Sci. China Life Sci. 2024, 67, 1766–1768. [Google Scholar] [CrossRef]
- Palli, S.R. RNAi turns 25:contributions and challenges in insect science. Front. Insect Sci. 2023, 3, 1209478. [Google Scholar] [CrossRef]
- Wei, H.; Tan, S.; Yan, S.; Li, Z.; Shen, J.; Liu, X. Nanocarrier-mediated transdermal dsRNA-NPF1 delivery system contributes to pest control via inhibiting feeding behavior in Grapholita molesta. J. Pest Sci. 2022, 95, 983–995. [Google Scholar] [CrossRef]
- Lu, J.; Shen, J. Target genes for RNAi in pest control: A comprehensive overview. Entomol. Gen. 2024, 44, 95–114. [Google Scholar] [CrossRef]
- Silver, K.; Cooper, A.M.W.; Zhu, K.Y. Strategies for enhancing the efficiency of RNA interference in insects. Pest Manag. Sci. 2021, 77, 2645–2658. [Google Scholar] [CrossRef] [PubMed]
- Badillo-Vargas, I.E.; Rotenberg, D.; Schneweis, B.A.; Whitfield, A.E. RNA interference tools for the western flower thrips, Frankliniella occidentalis. J. Insect Physiol. 2015, 76, 36–46. [Google Scholar] [CrossRef]
- Han, S.H.; Kim, J.H.; Kim, K.; Lee, S.H. Selection of lethal genes for ingestion RNA interference against western flower thrips, Frankliniella occidentalis, via leaf disc-mediated dsRNA delivery. Pestic. Biochem. Physiol. 2019, 161, 47–53. [Google Scholar] [CrossRef]
- Zhang, T.; Liu, L.; Zhi, J.R.; Jia, Y.L.; Yue, W.B.; Zeng, G.; Li, D.-Y. Copper chaperone for superoxide dismutase FoCCS1 in Frankliniella occidentalis may be associated with feeding adaptation after host shifting. Insects 2022, 13, 782. [Google Scholar] [CrossRef]
- Andongma, A.A.; Greig, C.; Dyson, P.J.; Flynn, N.; Whitten, M.M.A. Optimization of dietary RNA interference delivery to western flower thrips Frankliniella occidentalis and onion thrips Thrips tabaci. Arch. Insect Biochem. Physiol. 2020, 103, e21645. [Google Scholar] [CrossRef]
- Venkatesh, J.; Kim, S.J.; Siddique, M.I.; Kim, J.H.; Lee, S.H.; Kang, B.C. CopE and TLR6 RNAi-mediated tomato resistance to western flower thrips. J. Integr. Agric. 2023, 22, 471–480. [Google Scholar] [CrossRef]
- Whitten, M.M.A.; Facey, P.D.; del Sol, R.; Fernández-Martínez, L.T.; Evans, M.C.; Mitchell, J.J.; Bodger, O.G.; Dyson, P.J. Symbiont-mediated RNA interference in insects. Proc. R. Soc. B Biol. Sci. 2016, 283, 20160042. [Google Scholar] [CrossRef] [PubMed]
- Caccia, S.; Astarita, F.; Barra, E.; Di Lelio, I.; Varricchio, P.; Pennacchio, F. Enhancement of Bacillus thuringiensis toxicity by feeding Spodoptera littoralis larvae with bacteria expressing immune suppressive dsRNA. J. Pest Sci. 2020, 93, 303–314. [Google Scholar] [CrossRef]
- Zhu, F.; Xu, J.J.; Palli, R.S.; Ferguson, J.; Palli, S.R. Ingested RNA interference for managing the populations of the Colorado potato beetle, Leptinotarsa decemlineata. Pest Manag. Sci. 2011, 67, 175–182. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, H.J.; Chen, J.Y.; Shen, Q.D.; Wang, S.G.; Xu, H.X.; Tang, B. Glycogen phosphorylase and glycogen synthase: Gene cloning and expression analysis reveal their role in trehalose metabolism in the brown planthopper, Nilaparvata lugens Stål (Hemiptera: Delphacidae). J. Insect Sci. 2017, 17, 42. [Google Scholar] [CrossRef]
- Zhao, L.N.; Yang, M.M.; Shen, Q.D.; Liu, X.J.; Shi, Z.K.; Wang, S.G.; Tang, B. Functional characterization of three trehalase genes regulating the chitin metabolism pathway in rice brown planthopper using RNA interference. Sci. Rep. 2016, 6, 27841. [Google Scholar] [CrossRef] [PubMed]
- Elbein, A.D.; Pan, Y.T.; Pastuszak, I.; Carroll, D. New insights on trehalose: A multifunctional molecule. Glycobiology 2003, 13, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.Z.; Huang, Y.L.; Lu, Z.J.; Zhang, Q.; Su, H.N.; Du, Y.M.; Yi, L.; Zhong, B.; Chen, C. Inhibition of trehalase affects the trehalose and chitin metabolism pathways in Diaphorina citri (Hemiptera: Psyllidae). Insect Sci. 2021, 28, 718–734. [Google Scholar] [CrossRef]
- Mitsumasu, K.; Kanamori, Y.; Fujita, M.; Iwata, K.I.; Tanaka, D.; Kikuta, S.; Watanabe, M.; Cornette, R.; Okuda, T.; Kikawada, T. Enzymatic control of anhydrobiosis-related accumulation of trehalose in the sleeping chironomid, Polypedilum vanderplanki. FEBS J. 2010, 277, 4215–4228. [Google Scholar] [CrossRef]
- Chen, Q.F.; Enbo, M.A.; Behar, K.L.; Xu, T.; Haddad, G.G. Role of trehalose phosphate synthase in anoxia tolerance and development in Drosophila melanogaster. J. Biol. Chem. 2002, 277, 3274–3279. [Google Scholar] [CrossRef] [PubMed]
- Jin, T.T.; Gao, Y.L.; He, K.L.; Ge, F. Expression profiles of the trehalose-6-phosphate synthase gene associated with thermal stress in Ostrinia furnacalis (Lepidoptera: Crambidae). J. Insect Sci. 2018, 18, 7. [Google Scholar] [CrossRef]
- Tang, B.; Wang, S.; Wang, S.G.; Wang, H.J.; Zhang, J.Y.; Cui, S.Y. Invertebrate trehalose-6-phosphate synthase gene: Genetic architecture, biochemistry, physiological function, and potential applications. Front. Physiol. 2018, 9, 30. [Google Scholar] [CrossRef]
- Yang, M.M.; Zhao, L.N.; Shen, Q.D.; Xie, G.Q.; Wang, S.G.; Tang, B. Knockdown of two trehalose-6-phosphate synthases severely affects chitin metabolism gene expression in the brown planthopper Nilaparvata lugens. Pest Manag. Sci. 2017, 73, 206–216. [Google Scholar] [CrossRef]
- Wang, G.; Gou, Y.; Guo, S.; Zhou, J.J.; Liu, C. RNA interference of trehalose-6-phosphate synthase and trehalase genes regulates chitin metabolism in two color morphs of Acyrthosiphon pisum Harris. Sci. Rep. 2021, 11, 948. [Google Scholar] [CrossRef]
- Zhang, J.H.; Qi, L.Z.; Chen, B.Y.; Li, H.Y.; Hu, L.L.; Wang, Q.T.; Wang, S.; Xi, J. Trehalose-6-phosphate synthase contributes to rapid cold hardening in the invasive insect Lissorhoptrus oryzophilus (Coleoptera: Curculionidae) by regulating trehalose metabolism. Insects 2023, 14, 903. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.X.; Lyu, Z.H.; Wang, C.Y.; Cheng, J.; Lin, T. RNA interference of a trehalose-6-phosphate synthase gene reveals its roles in the biosynthesis of chitin and lipids in Heortia vitessoides (Lepidoptera: Crambidae). Insect Sci. 2020, 27, 212–223. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Y.; Zou, Z.W.; Zhang, C.; Liu, X.; Wang, J.; Xin, T.R.; Xia, B. Knockdown of the trehalose-6-phosphate synthase gene using rna interference inhibits synthesis of trehalose and increases lethality rate in asian citrus psyllid, Diaphorina citri (Hemiptera: Psyllidae). Insects 2020, 11, 605. [Google Scholar] [CrossRef] [PubMed]
- Song, J.C.; Lu, Z.J.; Yi, L.; Yu, H.Z. Functional characterization of a trehalose-6-phosphate synthase in Diaphorina citri revealed by rna interference and transcriptome sequencing. Insects 2021, 12, 1074. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.; You, Y.; Zeng, Z.H.; Chen, Y.X.; Hu, J.F.; Lin, S.; Hu, Q.; Yang, F.; Wei, H. Temperature-dependent demography of Thrips hawaiiensis (Thysanoptera: Thripidae): Implications for prevention and control. Environ. Entomol. 2021, 50, 1455–1465. [Google Scholar] [CrossRef]
- Chi, H.; Güncan, A.; Kavousi, A.; Gharakhani, G.; Atlihan, R.; Özgökçe, M.S.; Shirazi, J.; Amir-Maafi, M.; Maroufpoor, M.; Roya, T. TWOSEX-MSChart: The key tool for life table research and education. Entomol. Gen. 2022, 42, 845–849. [Google Scholar] [CrossRef]
- Chi, H. TWOSEX-MSChart: A Computer Program for the Age-Stage, Two-Sex Life Table Analysis. 2023. Available online: http://140.120.197.173/Ecology/ (accessed on 25 December 2024).
- Chi, H. TIMING-MSChart: A Computer Program for the Population Projection Based on Age-Stage, Two-Sex Life Table. 2023. Available online: http://140.120.197.173/Ecology/ (accessed on 25 December 2024).
- Argüelles, J.C. Why can’t vertebrates synthesize trehalose? J. Mol. Evol. 2014, 79, 111–116. [Google Scholar] [CrossRef]
- Zhang, H.Z.; Wang, M.Q.; Xie, Y.Q.; Xiang, M.; Li, P.; Li, Y.Y.; Zhang, L.S. Gene cloning and expression analysis of trehalose-6-phosphate synthase, glycogen synthase and glycogen phosphorylase reveal the glycometabolism in the diapause process of Aphidius gifuensis. J. Asia Pac. Entomol. 2020, 23, 641–645. [Google Scholar] [CrossRef]
- Zhang, Y.M.; Wang, F.; Feng, Q.; Wang, H.X.; Tang, T.; Huang, D.W.; Liu, F. Involvement of trehalose-6-phosphate synthase in innate immunity of Musca domestica. Dev. Comp. Immunol. 2019, 91, 85–92. [Google Scholar] [CrossRef]
- Xiong, K.C.; Wang, J.; Li, J.H.; Deng, Y.Q.; Pu, P.; Fan, H.; Liu, Y.-H. RNA interference of a trehalose-6-phosphate synthase gene reveals its roles during larval-pupal metamorphosis in Bactrocera minax (Diptera: Tephritidae). J. Insect Physiol. 2016, 91–92, 84–92. [Google Scholar] [CrossRef]
- Lü, X.; Han, S.C.; Li, Z.G.; Li, L.Y.; Li, J. Gene characterization and enzymatic activities related to trehalose metabolism of in vitro reared Trichogramma dendrolimi Matsumura (Hymenoptera: Trichogrammatidae) under sustained cold stress. Insects 2020, 11, 767. [Google Scholar] [CrossRef] [PubMed]
- Gong, C.; Yang, Z.Z.; Hu, Y.; Wu, Q.J.; Wang, S.L.; Guo, Z.J.; Zhang, Y. Silencing of the BtTPS genes by transgenic plant-mediated RNAi to control Bemisia tabaci MED. Pest Manag. Sci. 2022, 78, 1128–1137. [Google Scholar] [CrossRef]
- Souza-Ferreira, P.S.; Mansur, J.F.; Berni, M.; Moreira, M.F.; Dos Santos, R.E.; Araújo, H.M.M.; de Souza, W.; Ramos, I.B.; Masuda, H. Chitin deposition on the embryonic cuticle of Rhodnius prolixus: The reduction of CHS transcripts by CHS-dsRNA injection in females affects chitin deposition and eclosion of the first instar nymph. Insect Biochem. Mol. Biol. 2014, 51, 101–109. [Google Scholar] [CrossRef]
- Yu, C.; Zhao, R.N.; Zhou, W.; Pan, Y.N.; Tian, H.; Yin, Z.Y.; Chen, W. Fruit fly in a challenging environment: Impact of short-term temperature stress on the survival, development, reproduction, and trehalose metabolism of Bactrocera dorsalis (Diptera: Tephritidae). Insects 2022, 13, 753. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.L.; Lei, G.K.; Chen, Y.T.; You, M.S.; You, S.J. PxTret1-like affects the temperature adaptability of a cosmopolitan pest by altering trehalose tissue distribution. Int. J. Mol. Sci. 2022, 23, 9019. [Google Scholar] [CrossRef]
- Li, Y.; Wu, S.J.; Xu, Y.H.; Li, Y.Y.; Liu, Y.H.; Wang, J. Transcriptomic identification and characterization of trehalose-6-phosphate synthase in fat body of the oriental fruit fly, Bactrocera dorsalis. J. Agric. Food Chem. 2024, 72, 5725–5733. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.Z.; Cao, L.J.; Chen, J.C.; Chen, W.B.; Shen, X.J.; Song, W.; Yang, F.; Wei, S.-J. A nanocarrier-mediated dsRNA oral delivery enhances RNAi efficiency in thrips. Entomol. Gen. 2024, 44, 601–611. [Google Scholar] [CrossRef]
- Gillet, F.X.; Garcia, R.A.; Macedo, L.L.P.; Albuquerque, E.V.S.; Silva, M.C.M.; Grossi-de-Sa, M.F. Investigating engineered ribonucleoprotein particles to improve oral RNAi delivery in crop insect pests. Front. Physiol. 2017, 8, 256. [Google Scholar] [CrossRef]
- Liang, H.Y.; Yang, X.M.; Sun, L.J.; Zhao, C.D.; Chi, H.; Zheng, C.Y. Sublethal effect of spirotetramat on the life table and population growth of Frankliniella occidentalis (Thysanoptera: Thripidae). Entomol. Gen. 2021, 41, 219–231. [Google Scholar] [CrossRef]
- Zhu, Q.; Arakane, Y.; Beeman, R.W.; Kramer, K.J.; Muthukrishnan, S. Functional specialization among insect chitinase family genes revealed by RNA interference. Proc. Natl. Acad. Sci. USA 2008, 105, 6650–6655. [Google Scholar] [CrossRef]
- Tang, B.; Wei, P.; Zhao, L.; Shi, Z.; Shen, Q.; Yang, M.; Xie, G.; Wang, S. Knockdown of five trehalase genes using RNA interference regulates the gene expression of the chitin biosynthesis pathway in Tribolium castaneum. BMC Biotechnol. 2016, 16, 67. [Google Scholar] [CrossRef] [PubMed]
- Chi, H.; Kavousi, A.; Gharekhani, G.; Atlihan, R.; Özgökçe, M.S.; Güncan, A.; Gökçe, A.; Smith, C.L.; Benelli, G.; Guedes, R.N.C.; et al. Advances in theory, data analysis, and application of the age-stage, two-sex life table for demographic research, biological control, and pest management. Entomol. Gen. 2023, 43, 705–732. [Google Scholar] [CrossRef]
- Wu, L.Y.; Wei, G.H.; Yan, Y.; Zhou, X.G.; Zhu, X.; Zhang, Y.H.; Li, X. Effects of miR-306 perturbation on life parameters in the English grain aphid, Sitobion avenae (Homoptera: Aphididae). Int. J. Mol. Sci. 2024, 25, 5680. [Google Scholar] [CrossRef]
- Cedden, D.; Güney, G.; Scholten, S.; Rostás, M. Lethal and sublethal effects of orally delivered double-stranded RNA on the cabbage stem flea beetle, Psylliodes chrysocephala. Pest Manag. Sci. 2023, 80, 2282–2293. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.X.; Chen, J.Z.; Li, M.W.; Zha, B.H.; Huang, P.R.; Chu, X.M.; Chen, J.; Yang, G. The combination of Bacillus thuringiensis and its engineered strain expressing dsRNA increases the toxicity against Plutella xylostella. Int. J. Mol. Sci. 2022, 23, 444. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Cui, L.; Wang, Q.Q.; Chang, Y.P.; Huang, W.L.; Rui, C.H. Sulfoxaflor resistance in Aphis gossypii: Resistance mechanism, feeding behavior and life history changes. J. Pest Sci. 2022, 95, 811–825. [Google Scholar] [CrossRef]
- Coll, M.; Shakya, S.; Shouster, I.; Nenner, Y.; Steinberg, S. Decision-making tools for Frankliniella occidentalis management in strawberry: Consideration of target markets. Entomol. Exp. Appl. 2007, 122, 59–67. [Google Scholar] [CrossRef]
- Reitz, S.R.; Gao, Y.; Kirk, W.D.J.; Hoddle, M.S.; Leiss, K.A.; Funderburk, J.E. Invasion biology, ecology, and management of western flower thrips. Annu. Rev. Entomol. 2020, 65, 17–37. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage (Parameter) | n | HT115-EGFP | n | HT115-TPS |
|---|---|---|---|---|
| Duration of egg/d | 40 | 4.3 ± 0.1 | 37 | 4.1 ± 0.1 |
| Duration of nymph/d | 38 | 7.4 ± 0.3 | 28 | 7.3 ± 0.2 |
| Duration of pupa/d | 34 | 5.4 ± 0.1 | 26 | 5.2 ± 0.2 |
| Duration of preadult/d | 34 | 16.9 ± 0.2 | 26 | 16.9 ± 0.2 |
| Adult pre-reproductive period, APOP/d | 18 | 2.1 ± 0.2 | 14 | 1.9 ± 0.3 |
| Total pre-reproductive period, TPOP/d | 18 | 18.8 ± 0.2 | 14 | 19.3 ± 0.1 * |
| Total female longevity/d | 18 | 65.3 ± 4.7 | 14 | 43.1 ± 2.6 * |
| Total male longevity/d | 16 | 54.2 ± 4.9 | 12 | 39.8 ± 2.6 * |
| Age of 50% survival/d | 40 | 54.0 ± 6.5 | 40 | 31.9 ± 4.9 * |
| Oviposition days/d | 18 | 45.5 ± 4.4 | 14 | 23.3 ± 2.4 * |
| Fecundity | 18 | 209.2 ± 16.7 | 14 | 54.1 ± 4.2 * |
| Peak reproduction value | 40 | 47.4 ± 2.0 | 40 | 26.0 ± 1.5 * |
| Age of peak reproduction value/d | 40 | 21.0 ± 0.3 | 40 | 21.0 ± 0.5 |
| Intrinsic rate of increase, r | 40 | 0.1448 ± 0.0071 | 40 | 0.1074 ± 0.0091 * |
| Finite rate of increase, λ | 40 | 1.1558 ± 0.0082 | 40 | 1.1134 ± 0.0101 * |
| Net reproductive rate, R0 | 40 | 94.1 ± 18.1 | 40 | 19.0 ± 4.3 * |
| Mean generation time, T/d | 40 | 31.4 ± 0.6 | 40 | 27.4 ± 0.4 * |
| Stages | n | HT115-EGFP | n | HT115-TPS |
|---|---|---|---|---|
| Egg | 40 | 0.000 ± 0.000 | 40 | 0.075 ± 0.042 |
| Nymph | 40 | 0.050 ± 0.034 | 40 | 0.225 ± 0.066 * |
| Pupa | 40 | 0.100 ± 0.047 | 40 | 0.050 ± 0.034 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, T.; Chen, X.; Chen, Y.; Chen, T.; Liang, X.; Wei, H.; Yang, G. Effects of Orally Delivered Double-Stranded RNA of Trehalose-6-Phosphate Synthase on the Population of Frankliniella occidentalis. Insects 2025, 16, 614. https://doi.org/10.3390/insects16060614
Lin T, Chen X, Chen Y, Chen T, Liang X, Wei H, Yang G. Effects of Orally Delivered Double-Stranded RNA of Trehalose-6-Phosphate Synthase on the Population of Frankliniella occidentalis. Insects. 2025; 16(6):614. https://doi.org/10.3390/insects16060614
Chicago/Turabian StyleLin, Tao, Xiaoyu Chen, Ying Chen, Ting Chen, Xueyi Liang, Hui Wei, and Guang Yang. 2025. "Effects of Orally Delivered Double-Stranded RNA of Trehalose-6-Phosphate Synthase on the Population of Frankliniella occidentalis" Insects 16, no. 6: 614. https://doi.org/10.3390/insects16060614
APA StyleLin, T., Chen, X., Chen, Y., Chen, T., Liang, X., Wei, H., & Yang, G. (2025). Effects of Orally Delivered Double-Stranded RNA of Trehalose-6-Phosphate Synthase on the Population of Frankliniella occidentalis. Insects, 16(6), 614. https://doi.org/10.3390/insects16060614