
Organic Farming Shapes Population Dynamics and Genetic Diversity of Euborellia annulipes in Banana Groves


Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sampling Design
2.3. Statistical Analysis
2.4. DNA Extraction, Amplification, and Sequencing
2.5. Genetic Analyses
3. Results
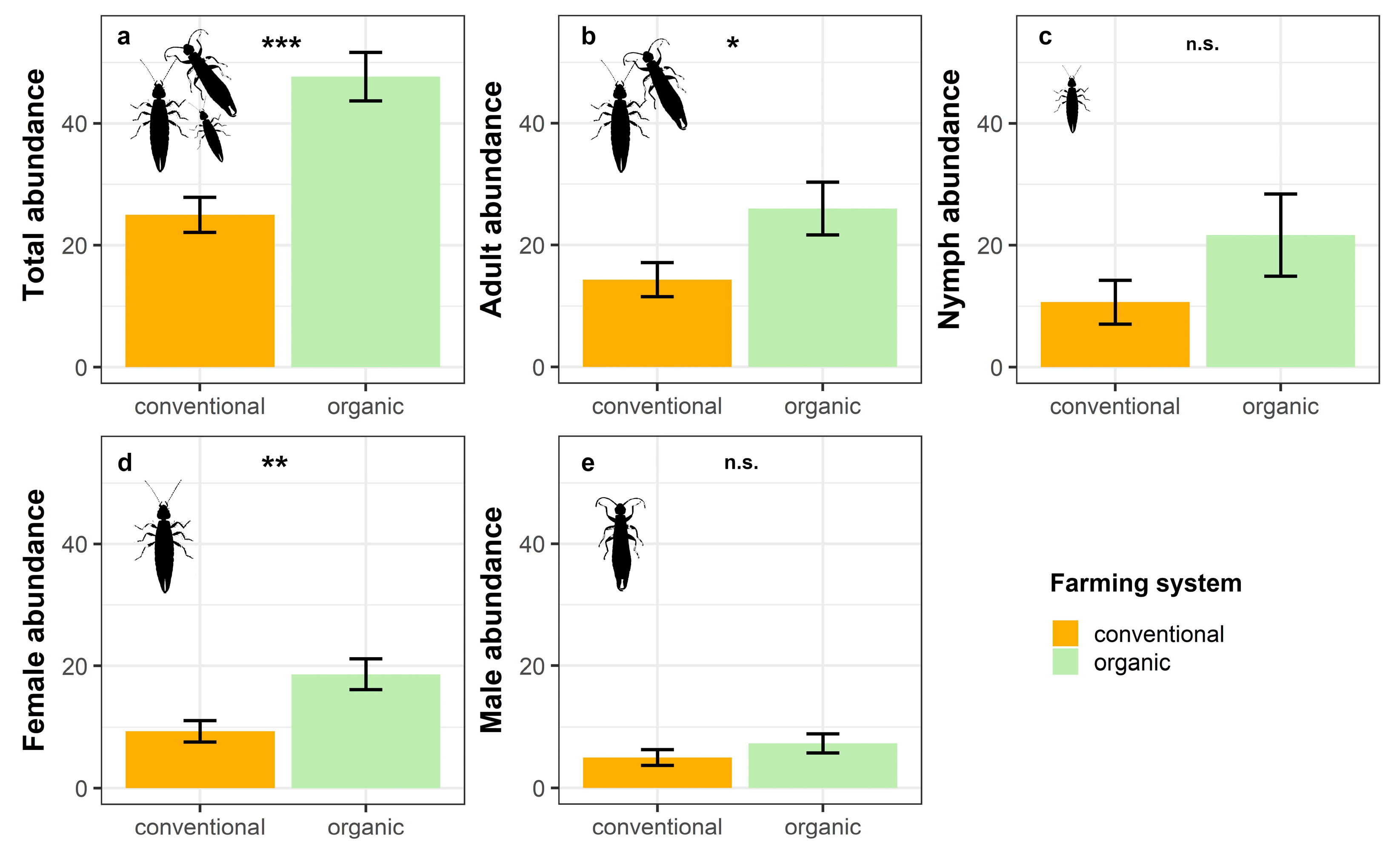
3.1. Effects of Farming System on the Abundance of Euborellia annulipes
3.2. Genetic Diversity
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reiff, J.M.; Kolb, S.; Entling, M.H.; Herndl, T.; Möth, S.; Walzer, A.; Kropf, M.; Hoddmann, C.; Winter, S. Organic farming and cover-crop management reduce pest predation in Austrian vineyards. Insects 2021, 12, 220. [Google Scholar] [CrossRef] [PubMed]
- Mburu, S.W.; Koskey, G.; Kimiti, J.M.; Ombori, O.; Maingi, J.M.; Njeru, E.M. Agrobiodiversity conservation enhances food security in subsistence-based farming systems of Eastern Kenya. Agric. Food Secur. 2016, 5, 19. [Google Scholar] [CrossRef]
- FAO—Food and Agriculture Organization of the United Nations. Risk Reduction and Mainstreaming Biodiversity in Agriculture. 2024. Available online: https://www.fao.org/pest-and-pesticide-management/pesticide-risk-reduction/risk-reduction-mainstreaming-biodiversity/en/ (accessed on 5 May 2025).
- Salem, K.F.; Ibrahim, A.A. Plant Biodiversity in the Context of Food Security Under Climate Change. In Sustainable Utilization and Conservation of Plant Genetic Diversity; Al-Khayri, J.M., Mohan Jain, S., Suprasanna, P., Eds.; Springer Nature: Singapore, 2024; pp. 3–30. [Google Scholar]
- Uchida, K.; Ushimaru, A. Biodiversity declines due to abandonment and intensification of agricultural lands: Patterns and mechanisms. Ecol. Monogr. 2014, 84, 637–658. [Google Scholar] [CrossRef]
- Habel, J.C.; Ulrich, W.; Biburger, N.; Seibold, S.; Schmitt, T. Agricultural intensification drives butterfly decline. Insect Conserv. Divers. 2019, 12, 289–295. [Google Scholar] [CrossRef]
- Stein-Bachinger, K.; Gottwald, F.; Haub, A.; Schmidt, E. To what extent does organic farming promote species richness and abundance in temperate climates? A review. Org. Agric. 2021, 11, 1–12. [Google Scholar] [CrossRef]
- Stein-Bachinger, K.; Fuchs, S. Protection strategies for farmland birds in legume-grass leys as trade-offs between nature conservation and farmers’ needs. Org. Agric. 2012, 2, 145–162. [Google Scholar] [CrossRef]
- Gottwald, F.; Stein-Bachinger, K. Landwirtschaft für Artenvielfalt–Ein Naturschutzmodul für Ökologisch Bewirtschaftete Betriebe. 2016. 208p. Available online: www.landwirtschaft-artenvielfalt.de (accessed on 5 May 2025).
- Landis, D.A. Designing agricultural landscapes for biodiversity-based ecosystem services. Basic Appl. Ecol. 2017, 18, 1–12. [Google Scholar] [CrossRef]
- Tscharntke, T.; Grass, I.; Wanger, T.C.; Westphal, C.; Batáry, P. Beyond organic farming–harnessing biodiversity-friendly landscapes. Trends Ecol. Evol. 2021, 36, 919–930. [Google Scholar] [CrossRef]
- Rosas-Ramos, N.; Asís, J.D.; Tobajas, E.; de Paz, V.; Baños-Picón, L. Disentangling the Benefits of Organic Farming for Beetle Communities (Insecta: Coleoptera) in Traditional Fruit Orchards. Agriculture 2022, 12, 243. [Google Scholar] [CrossRef]
- Mondelaers, K.; Aertsens, J.; Van Huylenbroeck, G. A meta—analysis of the differences in environmental impacts between organic and conventional farming. Br. Food J. 2009, 111, 1098–1119. [Google Scholar] [CrossRef]
- Tuck, S.L.; Winqvist, C.; Mota, F.; Ahnström, J.; Turnbull, L.A.; Bengtsson, J. Land-use intensity and the effects of organic farming on biodiversity: A hierarchical meta-analysis. J. Appl. Ecol. 2014, 51, 746–755. [Google Scholar] [CrossRef] [PubMed]
- Feber, R.E.; Johnson, P.J.; Bell, J.R.; Chamberlain, D.E.; Firbank, L.G.; Fuller, R.J.; Manley, W.; Mathews, F.; Norton, L.R.; Townsend, M.; et al. Organic farming: Biodiversity impacts can depend on dispersal characteristics and landscape context. PLoS ONE 2015, 10, e0135921. [Google Scholar] [CrossRef] [PubMed]
- Froidevaux, J.S.; Louboutin, B.; Jones, G. Does organic farming enhance biodiversity in Mediterranean vineyards? A case study with bats and arachnids. Agric. Ecosyst. Environ. 2017, 249, 112–122. [Google Scholar] [CrossRef] [PubMed]
- Porcel, M.; Andersson, G.K.S.; Pålsson, J.; Tasin, M. Organic management in apple orchards: Higher impacts on biological control than on pollination. J. Appl. Ecol. 2018, 55, 2779–2789. [Google Scholar] [CrossRef]
- Gamage, A.; Gangahagedara, R.; Gamage, J.; Jayasinghe, N.; Kodikara, N.; Suraweera, P.; Merah, O. Role of organic farming for achieving sustainability in agriculture. Farming Syst. 2023, 1, 100005. [Google Scholar] [CrossRef]
- Sidemo-Holm, W.; Brady, M.V.; Carrié, R.; Ekroos, J.; Smith, H.G. Cost-effective biodiversity conservation with organic farming-spatial allocation is key. Biol. Conserv. 2024, 294, 110624. [Google Scholar] [CrossRef]
- Bengtsson, J.; Ahnström, J.; Weibull, A.C. The effects of organic agriculture on biodiversity and abundance: A meta-analysis. J. Appl. Ecol. 2005, 42, 261–269. [Google Scholar] [CrossRef]
- Kehinde, T.; von Wehrden, H.; Samways, M.; Klein, A.M.; Brittain, C. Organic farming promotes bee abundance in vineyards in Italy but not in South Africa. J. Insect Conserv. 2018, 22, 61–67. [Google Scholar] [CrossRef]
- Happe, A.K.; Roquer-Beni, L.; Bosch, J.; Alins, G.; Mody, K. Earwigs and woolly apple aphids in integrated and organic apple orchards: Responses of a generalist predator and a pest prey to local and landscape factors. Agric. Ecosyst. Environ. 2018, 268, 44–51. [Google Scholar] [CrossRef]
- Rosas-Ramos, N.; Banos-Picon, L.; Tormos, J.; Asis, J.D. Natural enemies and pollinators in traditional cherry orchards: Functionally important taxa respond differently to farming system. Agric. Ecosyst. Environ. 2020, 295, 106920. [Google Scholar] [CrossRef]
- Altieri, M.A.; Nicholls, C.I. Biodiveristy and Pest Management in Agroecosystems; Haworth Press: New York, NY, USA, 2004; 252p. [Google Scholar]
- Matzke, D.; Kočárek, P. Description and biology of Euborellia arcanum sp. nov., an alien earwig occupying greenhouses in Germany and Austria (Dermaptera: Anisolabididae). Zootaxa 2015, 3956, 131–139. [Google Scholar] [CrossRef]
- Fried, G.; Chauvel, B.; Reynaud, P.; Sache, I. Decreases in crop production by non-native weeds, pests, and pathogens. In Impact of Biological Invasions on Ecosystem Services; Vilà, M., Hulme, P.E., Eds.; Springer: Cham, Switzerland, 2017; pp. 83–101. [Google Scholar]
- Quarrell, S.R.; Arabi, J.; Suwalski, A.; Veuille, M.; Wirth, T.; Allen, G.R. The invasion biology of the invasive earwig, Forficula auricularia in Australasian ecosystems. Biol. Invasions 2018, 20, 1553–1565. [Google Scholar] [CrossRef]
- Hulme, P.E. Trade, transport and trouble: Managing invasive species pathways in an era of globalization. J. Appl. Ecol. 2009, 46, 10–18. [Google Scholar] [CrossRef]
- Pyšek, P.; Hulme, P.E.; Simberloff, D.; Bacher, S.; Blackburn, T.M.; Carlton, J.T.; Dawson, W.; Essl, F.; Foxcroft, L.C.; Genovesi, P.; et al. Scientists’ warning on invasive alien species. Biol. Rev. 2020, 95, 1511–1534. [Google Scholar] [CrossRef] [PubMed]
- Kenis, M.; Auger-Rozenberg, M.A.; Roques, A.; Timms, L.; Péré, C.; Cock, M.J.; Settele, J.; Augustin, S.; Lopez-Vaamonde, C. Ecological effects of invasive alien insects. Biol. Invasions 2009, 11, 21–45. [Google Scholar] [CrossRef]
- Bradshaw, C.J.; Leroy, B.; Bellard, C.; Roiz, D.; Albert, C.; Fournier, A.; Barbet-Massin, M.; Saller, J.; Simard, F.; Courchamp, F. Massive yet grossly underestimated global costs of invasive insects. Nat. Commun. 2016, 7, 12986. [Google Scholar] [CrossRef] [PubMed]
- McEvoy, P.; Cox, C.; Coombs, E. Successful biological control of ragwort, Senecio jacobaea, by introduced insects in Oregon. Ecol. Appl. 1991, 1, 430–442. [Google Scholar] [CrossRef]
- Baker, B.P.; Green, T.A.; Loker, A.J. Biological control and integrated pest management in organic and conventional systems. Biol. Control 2020, 140, 104095. [Google Scholar] [CrossRef]
- Olden, J.D.; Poff, N.L.; Douglas, M.R.; Douglas, M.E.; Fausch, K.D. Ecological and evolutionary consequences of biotic homogenization. Trends Ecol. Evol. 2004, 19, 18–24. [Google Scholar] [CrossRef]
- Holl, K.D.; Luong, J.C.; Brancalion, P.H. Overcoming biotic homogenization in ecological restoration. Trends Ecol. Evol. 2022, 37, 777–788. [Google Scholar] [CrossRef]
- Gauffre, B.; Boissinot, A.; Quiquempois, V.; Leblois, R.; Grillet, P.; Morin, S.; Picard, D.; Ribout, C.; Lourdais, O. Agricultural intensification alters marbled newt genetic diversity and gene flow through density and dispersal reduction. Mol. Ecol. 2022, 31, 119–133. [Google Scholar] [CrossRef] [PubMed]
- Nolen, Z.J.; Jamelska, P.K.; Lara, A.S.T.; Wahlberg, N.; Runemark, A. Species-specific loss of genetic diversity and accumulation of genetic load following agricultural intensification. bioRxiv 2024. [Google Scholar] [CrossRef]
- Grapputo, A.; Boman, S.; Lindstroem, L.; Lyytinen, A.; Mappes, J. The voyage of an invasive species across continents: Genetic diversity of North American and European Colorado potato beetle populations. Mol. Ecol. 2005, 14, 4207–4219. [Google Scholar] [CrossRef]
- Lambea-Camblor, A.; Morcillo, F.; Muñoz, J.; Perdices, A. Genetic and ecological approaches to introduced populations of pumpkinseed sunfish (Lepomis gibbosus) in Southwestern Europe. Diversity 2023, 15, 1059. [Google Scholar] [CrossRef]
- Mollot, G.; Duyck, P.F.; Lefeuvre, P.; Lescourret, F.; Martin, J.F.; Piry, S.; Canard, E.; Tixier, P. Cover cropping alters the diet of arthropods in a banana plantation: A metabarcoding approach. PLoS ONE 2014, 9, e93740. [Google Scholar] [CrossRef] [PubMed]
- Carval, D.; Resmond, R.; Achard, R.; Tixier, P. Cover cropping reduces the abundance of the banana weevil Cosmopolites sordidus but does not reduce its damage to the banana plants. Biol. Control 2016, 99, 14–18. [Google Scholar] [CrossRef]
- Orpet, R.J.; Crowder, D.W.; Jones, V.P. Biology and management of European earwig in orchards and vineyards. J. Integr. Pest. Manag. 2019, 10, 21. [Google Scholar] [CrossRef]
- Weidner, H. Einschleppung von Ohrwürmern nach Deutschland (Dermaptera). Anz. Für Schädlingskunde Pflanz. -Und Umweltschutz 1974, 47, 145–148. [Google Scholar] [CrossRef]
- Albouy, V.; Caussanel, C. Dermaptères ou Perce-oreilles. Faune de France 75; Fédération Française des Societés de Sciences Naturelles: Paris, France, 1990; 245p. [Google Scholar]
- Guillet, S.; Josselin, N.; Vancassel, M. Multiple introductions of the Forficula auricularia species complex (Dermaptera: Forficulidae) in eastern North America. Can. Entomol. 2000, 132, 49–57. [Google Scholar] [CrossRef]
- Vigna Taglianti, A. Insecta Dermaptera. In Checklist e Distribuzione Della Fauna Italiana; Memorie del Museo Civico di Storia naturale di Verona 2. Serie, Sezione Scienze della Vita; Ruffo, S., Stoch, F., Eds.; Ministero Dell’ambiente e Della Tutela del Territorio: Verona, Italy, 2005; pp. 141–142. [Google Scholar]
- Matzke, D. Aktuell synanthrop lebende Ohrwürmer in Deutschland. Dtsch. Ges. Für Allg. Angew. Entomol. Nachrichten 2018, 32, 6–9. [Google Scholar]
- González-Miguéns, R.; Muñoz-Nozal, E.; Jiménez-Ruiz, Y.; Mas-Peinado, P.; Ghanavi, H.R.; García-París, M. Speciation patterns in the Forficula auricularia species complex: Cryptic and not so cryptic taxa across the western Palaearctic region. Zool. J. Linn. Soc. 2020, 190, 788–823. [Google Scholar] [CrossRef]
- Zafeiriou, S.; Kočárek, P.; Kalaentzis, K. First record of the desert earwig Forficula lucasi Dohrn, 1865 (Dermaptera: Forficulidae) in Greece: A hitchhiker among the refugees or a seldom encounter? J. Insect Biodivers. 2021, 21, 15–17. [Google Scholar] [CrossRef]
- Kalaentzis, K.; Kazilas, C.; Agapakis, G.; Kocarek, P. Hidden in plain sight: First records of the alien earwig Euborellia femoralis (Dohrn, 1863) in Europe. BioInvasions Rec. 2021, 10, 1022–1031. [Google Scholar] [CrossRef]
- Nishikawa, M.; Kusui, Y. Earwigs (Dermaptera) collected in airplanes and ships called at ports in Japan. Tettigonia 2008, 9, 7–11. [Google Scholar]
- Rasplus, J.Y.; Roques, A. Dictyoptera (Blattodea, Isoptera), Orthoptera, Phasmatodea and Dermaptera. Chapter 13.3. BioRisk 2010, 4, 807–831. [Google Scholar] [CrossRef]
- Kocarek, P.; Dvorak, L.; Kirstova, M. Euborellia annulipes (Dermaptera: Anisolabididae), a new alien earwig in Central European greenhouses: Potential pest or beneficial inhabitant? Appl. Entomol. Zool. 2015, 50, 201–206. [Google Scholar] [CrossRef]
- Murányi, D.; Puskás, G. Hungarian occurrence of a potential horticultural pest earwig, Euborellia annulipes (Lucas, 1847) (Dermaptera: Anisolabididae). Növényvédelem 2018, 54, 513–517. [Google Scholar]
- Báez, M.; Relación de Las Categorías de Origen de Las Especies de Artrópodos de Canarias. Banco de Datos de Biodiversidad de Canarias. 2002, 73p. Available online: https://www.biodiversidadcanarias.es/biota/documento/A02090 (accessed on 6 November 2024).
- González-Concepción, C.; Gil-Fariña, M.C.; Pestano-Gabino, C. Multivariate modelling of the Canary Islands banana output. The role of farmer income expectation. J. Soc. Sci. 2008, 4, 88–97. [Google Scholar]
- Fuentes, E.G.; Hernández-Suárez, E.; Simón, O.; Williams, T.; Caballero, P. Chrysodeixis chalcites, a pest of banana crops on the Canary Islands: Incidence, economic losses and current control measures. Crop Prot. 2018, 108, 137–145. [Google Scholar] [CrossRef]
- Petrovan, S.; Aldridge, D.; Smith, R.; White, T.; Sutherland, W. Halyomorpha halys invasion front jumps 1500 kilometres to reach the Canary Islands; a framework for rapid response, identification of urgent questions, and assessment of potential impacts. ARPHA Prepr. 2022, 3, e84924. [Google Scholar]
- FAOSTAT. Food and Agriculture Data [WWW Document]. Available online: http://www.fao.org/faostat/en/#home (accessed on 11 February 2025).
- Robinson, J.C.; Galán-Saúco, V. Bananas and Plantains; CAB International: Wallingford, UK, 2010; Volume 19, 297p. [Google Scholar]
- Medina, F.M.; Nogales, M.; Farnworth, M.J.; Bonnaud, E. Human-cat relationship in an oceanic biosphere reserve: The case of La Palma Island, Canary archipelago. J. Nat. Conserv. 2016, 34, 8–14. [Google Scholar] [CrossRef]
- Irl, S.D.; Harter, D.E.; Steinbauer, M.J.; Gallego Puyol, D.; Fernández-Palacios, J.M.; Jentsch, A.; Beierkuhnlein, C. Climate vs. topography–spatial patterns of plant species diversity and endemism on a high-elevation island. J. Ecol. 2015, 103, 1621–1633. [Google Scholar] [CrossRef]
- Confederación Canaria de Empresarios. 8. Actividad del Sector Primario. In Informe Anual de la Economía Canaria; Confederación Canaria de Empresarios: Las Palmas, Spain, 2020; pp. 171–188. Available online: https://www.ccelpa.org/informe-anual/IA2020/pdf/indicadores/08-2020.pdf (accessed on 2 May 2025).
- Hernández-Hernández, R.; Kluge, J.; Ah-Peng, C.; González-Mancebo, J.M. Natural and human-impacted diversity of bryophytes along an elevational gradient on an oceanic island (La Palma, Canarias). PLoS ONE 2019, 14, e0213823. [Google Scholar] [CrossRef]
- Regulation (EU) 2018/848 of the European Parliament and of the Council of 30 May 2018 on organic production and labelling of organic products and repealing Council Regulation (EC) No 834/2007. Off. J. Eur. Union 2018, 150, 1–92.
- Purnomo, B.; Fahrurrozi, F.; Sariasih, Y.; Muktamar, Z.; Efendi, Z. Determination of potential bacteria from five different types of green biomass enriched liquid organic fertilizer for developing Bio-decomposer. Int. J. Agric. Technol. 2017, 13, 1175–1182. [Google Scholar]
- Excmo. Cabildo Insular de La Palma. Estudio de la Sostenibilidad del Cultivo de Platanera en la Isla de la Palma, Tratados Con Purines Enriquecidos y Compost. 2018. Available online: https://www.ecofincanogales.com/documentos/memoria-proyecto-platanera-ipna-cabildo-de-la-palma.pdf (accessed on 15 March 2025).
- D’hont, A.; Denoeud, F.; Aury, J.M.; Baurens, F.C.; Carreel, F.; Garsmeur, O.; Noel, B.; Bocs, S.; Droc, G.; Rouard, M.; et al. The banana (Musa acuminata) genome and the evolution of monocotyledonous plants. Nature 2012, 488, 213–217. [Google Scholar] [CrossRef]
- Méndez Hernández, C.; Rodríguez Serrano, M. Deshijado de la Platanera. AgroCabildo, Excmo. Cabildo Insular de Tenerife. 2016. 20p. Available online: https://www.agrocabildo.org/publica/Publicaciones/subt_596_platanera.pdf (accessed on 21 November 2024).
- R Core Team. R: A Language and Environment for Statistical Computing (Version 4.5.0); R Foundation for Statistical Computing: Vienna, Austria, 2025; 3940p, Available online: https://cran.r-project.org (accessed on 5 May 2025).
- Simon, C.; Frati, F.; Beckenbach, A.; Crespi, B.; Liu, H.; Flook, P. Evolution, weighting and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction primers. Ann. Entomol. Soc. Am. 1994, 87, 651–701. [Google Scholar] [CrossRef]
- Pons, J. DNA-based identification of preys from non-destructive, total DNA extractions of predators using arthropod universal primers. Mol. Ecol. Notes 2006, 6, 623–626. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony (*and Other Methods) Version 4.0b10; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Paradis, E. pegas: An R package for population genetics with an integrated–modular approach. Bioinformatics 2010, 26, 419–420. [Google Scholar] [CrossRef]
- Kamvar, Z.N.; Tabima, J.F.; Grünwald, N.J. Poppr: An R package for genetic analysis of populations with clonal, partially clonal, and/or sexual reproduction. PeerJ 2014, 2, e281. [Google Scholar] [CrossRef]
- Dib, H.; Sauphanor, B.; Capowiez, Y. Effect of management strategies on arthropod communities in the colonies of rosy apple aphid, Dysaphis plantaginea Passerini (Hemiptera: Aphididae) in south-eastern France. Agric. Ecosyst. Environ. 2016, 216, 203–206. [Google Scholar] [CrossRef]
- Dib, H.; Sauphanor, B.; Capowiez, Y. Report on the life history traits of the generalist predator Forficula auricularia (Dermaptera Forficulidae) in organic apple orchards in southeastern France. Can. Entomol. 2017, 149, 56–72. [Google Scholar] [CrossRef]
- Simon, S.; Riotord, D.; Morel, K.; Fleury, A.; Capowiez, Y. A shift towards softer pest management differently benefits earwig and spider communities in a multiyear experiment in apple orchards. Pest Manag. Sci. 2024, 80, 5088–5098. [Google Scholar] [CrossRef]
- Moerkens, R.; Leirs, H.; Peusens, G.; Gobin, B. Are populations of European earwigs, Forficula auricularia, density dependent? Entomol. Exp. Appl. 2009, 130, 198–206. [Google Scholar] [CrossRef]
- Niedobová, J.; Skalský, S.; Ouředníčková, J.; Michalko, R. Forficula auricularia (Dermaptera) in orchards: Monitoring seasonal activity, the effect of pesticides, and the perception of European fruit growers on its role as a predator or pest. Pest Manag. Sci. 2021, 77, 1694–1704. [Google Scholar]
- Malagnoux, L.; Marliac, G.; Simon, S.; Rault, M.; Capowiez, Y. Management strategies in apple orchards influence earwig community. Chemosphere 2015, 124, 156–162. [Google Scholar] [CrossRef]
- Malagnoux, L.; Capowiez, Y.; Rault, M. Impact of insecticide exposure on the predation activity of the European earwig Forficula auricularia. Environ. Sci. Pollut. Res. 2015, 22, 14116–14126. [Google Scholar] [CrossRef]
- Le Navenant, A.; Brouchoud, C.; Capowiez, Y.; Rault, M.; Suchail, S. How lasting are the effects of pesticides on earwigs? A study based on energy metabolism, body weight and morphometry in two generations of Forficula auricularia from apple orchards. Sci. Total Environ. 2021, 758, 143604. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, A.C.; Stigall, T. Paternity and egg cannibalism in the ringlegged earwig Euborellia annulipes (Dermaptera: Anisolabididae). Entomol. Sci. 2019, 22, 250–257. [Google Scholar] [CrossRef]
- Lemos, W.P.; Ramalho, F.S.; Zanuncio, J.C. Age-dependent fecundity and life-fertility tables for Euborellia annulipes (Lucas) (Dermaptera: Anisolabididae) a cotton boll weevil predator in laboratory studies with an artificial diet. Environ. Entomol. 2003, 32, 592–601. [Google Scholar] [CrossRef]
- Núñez-Pascual, V.; Calleja, F.; Pardo, R.V.; Sarrazin, A.F.; Irles, P. The ring-legged earwig Euborellia annulipes as a new model for oogenesis and development studies in insects. J. Exp. Zool. Part B Mol. Dev. Evol. 2022, 340, 18–33. [Google Scholar] [CrossRef]
- Jamet, C.; Caussanel, C. Données biologiques, fonctionnement des appareils génitaux, comportements sexuels et maternels chez Euborellia annulipes (Lucas) (Dermaptère, Carcinophoridae). Bull. Société Entomol. Fr. 1995, 100, 37–58. [Google Scholar] [CrossRef]
- Fountain, M.T.; Harris, A.L. Non-target consequences of insecticides used in apple and pear orchards on Forficula auricularia L. (Dermaptera: Forficulidae). Biol. Control 2015, 91, 27–33. [Google Scholar] [CrossRef]
- Burton, M.; Burton, R. Earwigs. In International Wildlife Encyclopedia, 3rd ed.; Burton, M., Burton, R., Eds.; Marshall Cavendish Corporation: New York, NY, USA, 2002; pp. 738–740. [Google Scholar]
- Cross, J.; Fountain, M.; Marko, V.; Nagy, C. Arthropod ecosystem services in apple orchards and their economic benefits. Ecol. Entomol. 2015, 40, 82–96. [Google Scholar] [CrossRef]
- Jiang, Z.R.; Kajimura, H. Earwig preying on ambrosia beetle: Evaluating predatory process and prey preference. J. Appl. Entomol. 2020, 144, 743–750. [Google Scholar] [CrossRef]
- Byttebier, B.; Fischer, S. Predation on eggs of Aedes aegypti (Diptera: Culicidae): Temporal dynamics and identification of potential predators during the winter season in a temperate region. J. Med. Entomol. 2019, 56, 737–743. [Google Scholar] [CrossRef]
- Tangkawanit, U.; Seehavet, S.; Siri, N. The potential of Labidura riparia and Euborellia annulipes (Dermaptera) as predators of house fly in livestock. Songklanakarin J. Sci. Technol. 2021, 43, 603–607. [Google Scholar]
- Oliveira, L.V.Q.; Oliveria, R.; Nascimento Júnior, J.L.; Silva, I.T.F.A.; Barbosa, V.O.; Batista, J.L. Capacidade de busca da tesourinha Euborellia annulipes sobre o pulgão Brevicoryne brassicae (Hemiptera: Aphididae). PesquisAgro 2019, 2, 3–10. [Google Scholar] [CrossRef]
- Nunes, G.S.; Dantas, T.A.V.; Souza, M.S.; Nascimento, I.N.; Batista, J.L.; Malaquias, J.B. Life stage and population density of Plutella xylostella affect the predation behavior of Euborellia annulipes. Entomol. Exp. Appl. 2019, 167, 544–552. [Google Scholar] [CrossRef]
- Silva, A.B.; Batista, J.L.; de Brito, C.H. Capacidade predatória de Euborellia annulipes (Lucas, 1847) sobre Spodoptera frugiperda (Smith, 1797). Acta Sci. -Agron. 2009, 31, 7–11. [Google Scholar] [CrossRef]
- Coelho, R.S.; Pec, M.; Silva, A.L.R.; Peñaflor, M.F.; Marucci, R.C. Predation potential of the earwig Euborellia annulipes on fruit fly larvae and trophic interactions with the parasitoid Diachasmimorpha longicaudata. J. Appl. Entomol. 2023, 147, 147–156. [Google Scholar] [CrossRef]
- Marin Arroyo, R.; Mendoça De Souza, J.; Da Silva Nunes, G.; Gomes Ramalho, D.; De Bortoli, S.A. Euborellia annulipes mortality and predation on Diatraea saccharalis eggs after application of chemical and biological insecticides. Agric. Sci. 2023, 14, 11–22. [Google Scholar]
- Morato, R.P.; Cutler, G.C.; Torres, J.B. Insecticide compatibility with the predatory ring-legged earwig Euborelia annulipes increases mortality of diamondback moth. Biocontrol Sci. Technol. 2023, 33, 327–343. [Google Scholar] [CrossRef]
- Morato, R.P.; Nascimento, D.V.D.; Oliveira, G.M.; Bermúdez, N.C.; Lira, R.; Torres, J.B. Indoxacarb, cyantraniliprole, and Euborellia annulipes as options for integrated control of diamondback moth. J. Appl. Entomol. 2024, 148, 1300–1310. [Google Scholar] [CrossRef]
- Peusens, G.; Moerkens, R.; Beliën, T.; Gobin, B. Side effects of plant protection products and biological interactions on the European earwig Forficula auricularia L. Commun. Agric. Appl. Biol. Sci. 2009, 74, 411–417. [Google Scholar]
- Peusens, G.; Belien, T.; Gobin, B. Comparing different test methods for evaluating lethal side effects of some insecticides on the European earwig Forficula auricularia L. IOBC-WPRS Bull. 2010, 55, 95–100. [Google Scholar]
- Meunier, J.; Dufour, J.; Van Meyel, S.; Rault, M.; Lécureuil, C. Sublethal exposure to deltamethrin impairs maternal egg care in the European earwig Forficula auricularia. Chemosphere 2020, 258, 127383. [Google Scholar] [CrossRef]
- Hole, D.G.; Perkins, A.J.; Wilson, J.D.; Alexander, I.H.; Grice, P.V.; Evans, A.D. Does organic farming benefit biodiversity? Biol. Conserv. 2005, 122, 113–130. [Google Scholar] [CrossRef]
- Jacobsen, S.K.; Moraes, G.J.; Sørensen, H.; Sigsgaard, L. Organic cropping practice decreases pest abundance and positively influences predator-prey interactions. Agric. Ecosyst. Environ. 2019, 272, 1–9. [Google Scholar] [CrossRef]
- Galloway, A.D.; Seymour, C.L.; Gaigher, R.; Pryke, J.S. Organic farming promotes arthropod predators, but this depends on neighbouring patches of natural vegetation. Agric. Ecosyst. Environ. 2021, 310, 107295. [Google Scholar] [CrossRef]
- Sinclair, A.R.; Mduma, S.; Brashares, J.S. Patterns of predation in a diverse predator–prey system. Nature 2003, 425, 288–290. [Google Scholar] [CrossRef]
- Allendorf, F.W.; Luikart, G.; Aitken, S.N. Conservation and the Genetics of Populations, 2nd ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2012; 624p. [Google Scholar]
- Frankham, R. Genetics and extinction. Biol. Conserv. 2005, 126, 131–140. [Google Scholar] [CrossRef]
- Chateil, C.; Goldringer, I.; Tarallo, L.; Kerbiriou, C.; Le Viol, I.; Ponge, J.F.; Salmon, S.; Gachet, S.; Porcher, E. Crop genetic diversity benefits farmland biodiversity in cultivated fields. Agric. Ecosyst. Environ. 2013, 171, 25–32. [Google Scholar] [CrossRef]
- Fricaux, T.; Le Navenant, A.; Siegwart, M.; Rault, M.; Coustau, C.; Le Goff, G. The molecular resistance mechanisms of European earwigs from apple orchards subjected to different management strategies. Insects 2023, 14, 944. [Google Scholar] [CrossRef]
- Avise, J.C. Phylogeography: The History and Formation of Species; Harvard University Press: Cambridge, MA, USA, 2000; 447p. [Google Scholar]
- Jurado-Angulo, P.; García-París, M. Historical demographic determinants complement climate model predictions of co-occurring cryptic species. Divers. Distrib. 2025, 31, e70007. [Google Scholar] [CrossRef]
- Cuesta-Segura, A.D.; Jurado-Angulo, P.; Jiménez-Ruiz, Y.; García-París, M. Taxonomy of the Iberian species of Pseudochelidura (Dermaptera: Forficulidae). Eur. J. Taxon. 2023, 860, 81–115. [Google Scholar] [CrossRef]
- Biocan—Banco del Inventario Natural de Canarias. Available online: https://www.biodiversidadcanarias.es (accessed on 5 May 2025).
- Lucas, H. Description de cette nouvelle espèce. Ann. Société Entomol. Fr. 1847, 5, LXXXIV. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Specimen Code | Management | Locality | Nº Haplotype | GenBank Cytb |
|---|---|---|---|---|
| tij21135 | Organic | La Palma: El Granel | I | PV737866 |
| tij21136 | Organic | La Palma: El Granel | II | PV737867 |
| tij21161 | Organic | La Palma: Breña Baja | III | PV737868 |
| tij21162 | Organic | La Palma: Breña Baja | IV | PV737869 |
| tij21228 | Organic | La Palma: El Granel | V | PV737870 |
| tij21229 | Organic | La Palma: El Granel | I | PV737871 |
| tij21281 | Conventional | La Palma: La Polvacera | VI | PV737872 |
| tij21282 | Conventional | La Palma: La Polvacera | VII | PV737873 |
| tij21324 | Conventional | La Palma: Puntallana | I | PV737874 |
| tij21325 | Conventional | La Palma: Puntallana | VIII | PV737875 |
| tij21342 | Conventional | La Palma: Los Cancajos | IX | PV737876 |
| tij21343 | Conventional | La Palma: Los Cancajos | I | PV737877 |
| Response Variable | Factor | Estimate | Std. Error | z Value | p Value | |
|---|---|---|---|---|---|---|
| Total abundance | Intercept | 3.219 | 0.116 | 27.876 | <0.001 | *** |
| (GLMpoisson) | System (organic) | 0.645 | 0.143 | 4.527 | <0.001 | *** |
| Adult abundance | Intercept | 2.663 | 0.195 | 13.627 | <0.001 | *** |
| (GLMnb) | System (organic) | 0.596 | 0.257 | 2.319 | 0.020 | * |
| Nymph abundance | Intercept | 2.367 | 0.336 | 7.044 | <0.001 | *** |
| (GLMnb) | System (organic) | 0.709 | 0.458 | 1.546 | 0.122 | |
| Female abundance | Intercept | 2.234 | 0.189 | 11.819 | <0.001 | *** |
| (GLMpoisson) | System (organic) | 0.693 | 0.232 | 2.995 | 0.003 | ** |
| Male abundance | Intercept | 1.609 | 0.258 | 6.233 | <0.001 | *** |
| (GLMpoisson) | System (organic) | 0.383 | 0.335 | 1.144 | 0.253 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jurado-Angulo, P.; García-París, M.; Rosas-Ramos, N. Organic Farming Shapes Population Dynamics and Genetic Diversity of Euborellia annulipes in Banana Groves. Insects 2025, 16, 606. https://doi.org/10.3390/insects16060606
Jurado-Angulo P, García-París M, Rosas-Ramos N. Organic Farming Shapes Population Dynamics and Genetic Diversity of Euborellia annulipes in Banana Groves. Insects. 2025; 16(6):606. https://doi.org/10.3390/insects16060606
Chicago/Turabian StyleJurado-Angulo, Pilar, Mario García-París, and Natalia Rosas-Ramos. 2025. "Organic Farming Shapes Population Dynamics and Genetic Diversity of Euborellia annulipes in Banana Groves" Insects 16, no. 6: 606. https://doi.org/10.3390/insects16060606
APA StyleJurado-Angulo, P., García-París, M., & Rosas-Ramos, N. (2025). Organic Farming Shapes Population Dynamics and Genetic Diversity of Euborellia annulipes in Banana Groves. Insects, 16(6), 606. https://doi.org/10.3390/insects16060606