
The Ultrastructure of Olfactory Sensilla Across the Antenna of Monolepta signata (Oliver)

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Scanning Electron Microscopy (SEM)
2.3. Transmission Electron Microscopy (TEM)
2.4. Measurement and Data Analysis
3. Results
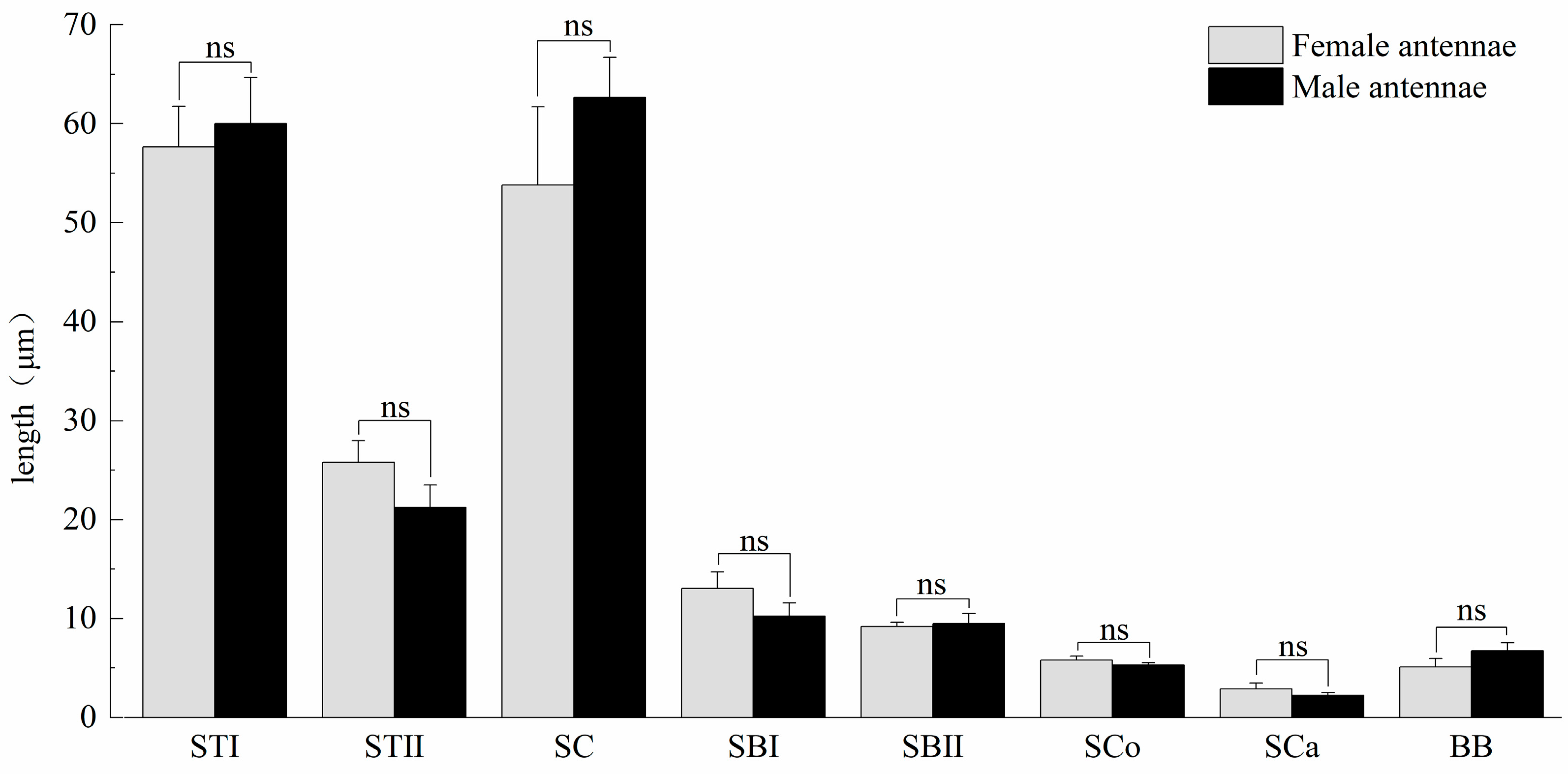
3.1. General Morphology of Antenna
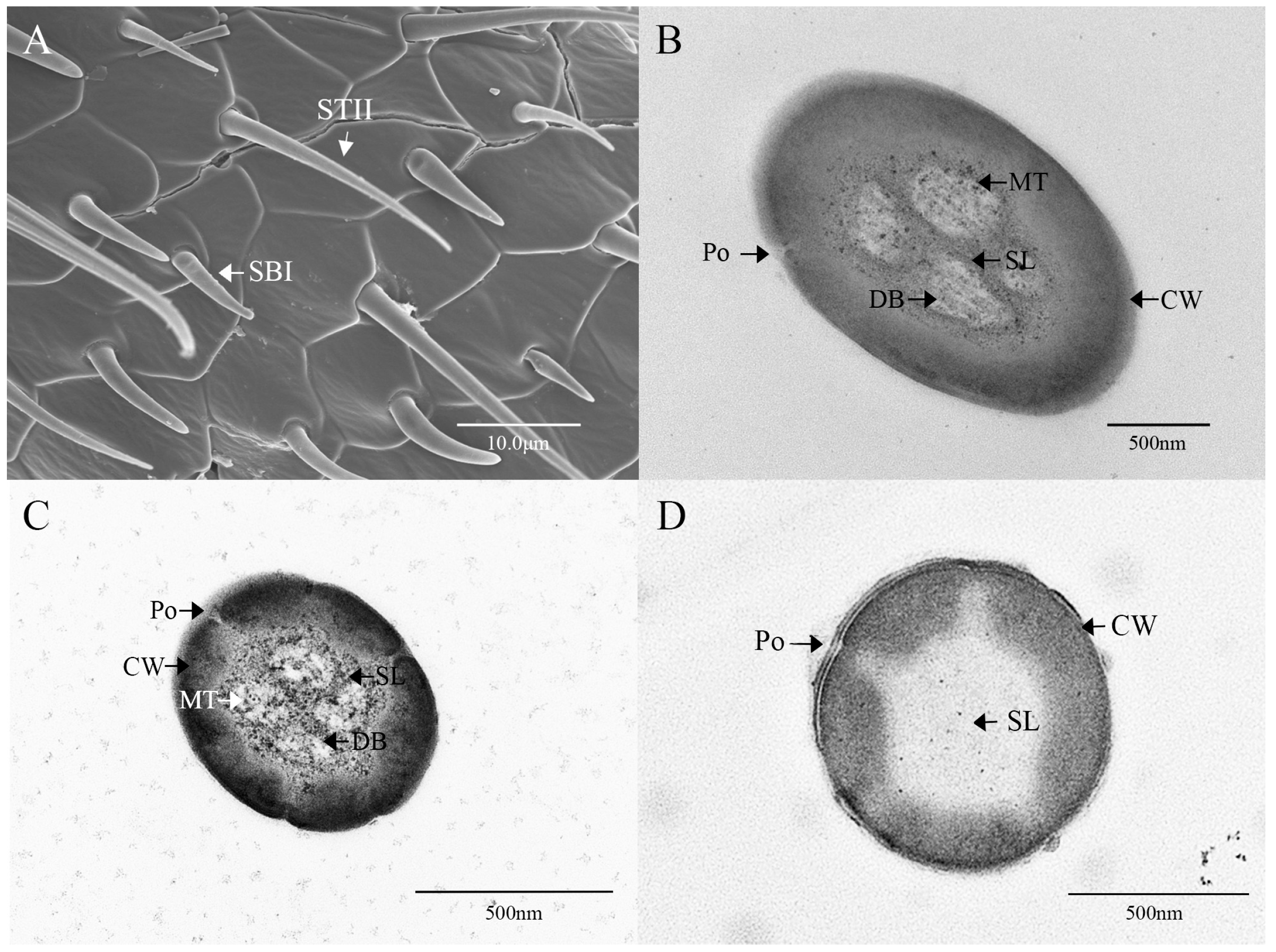
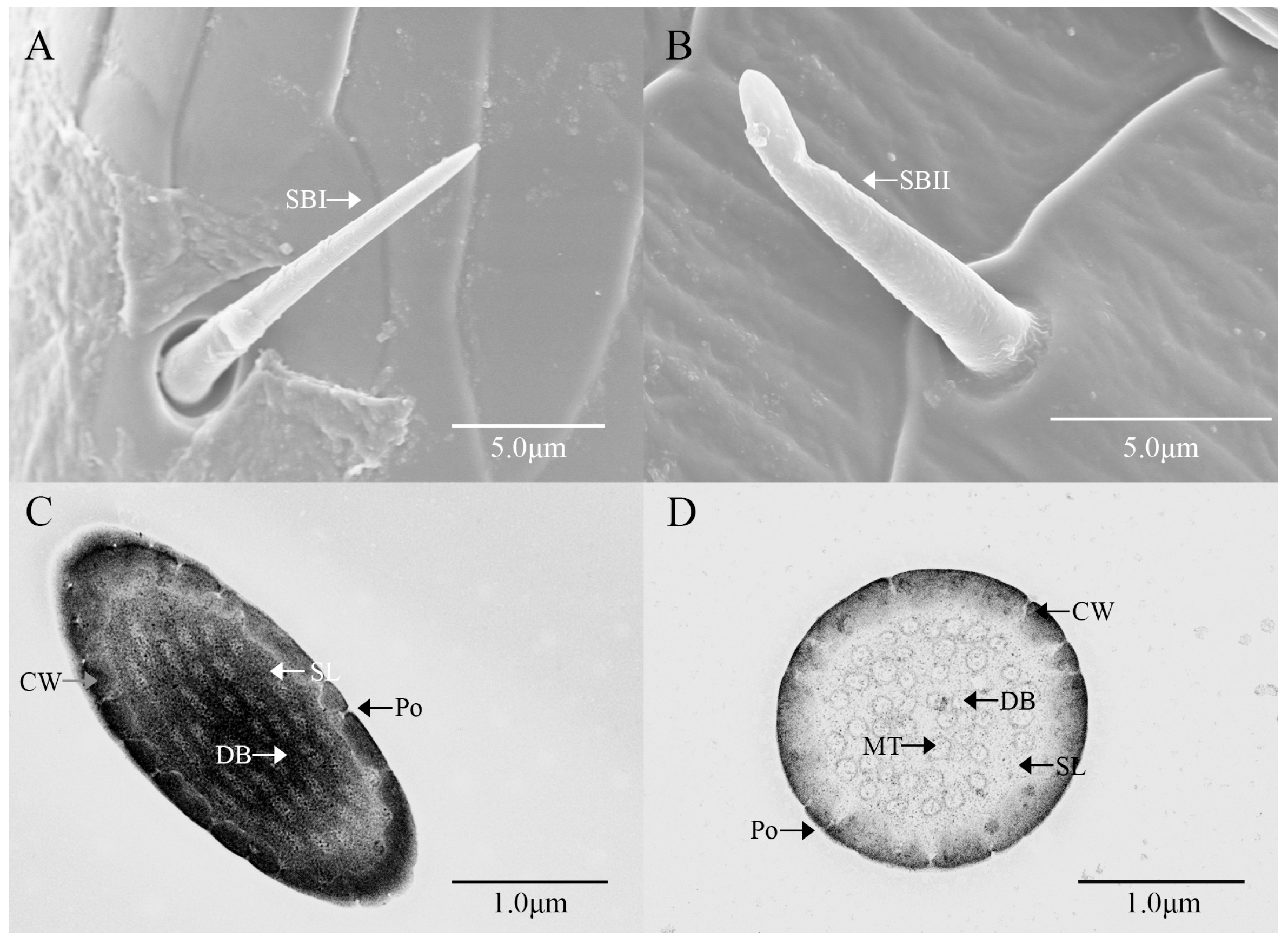
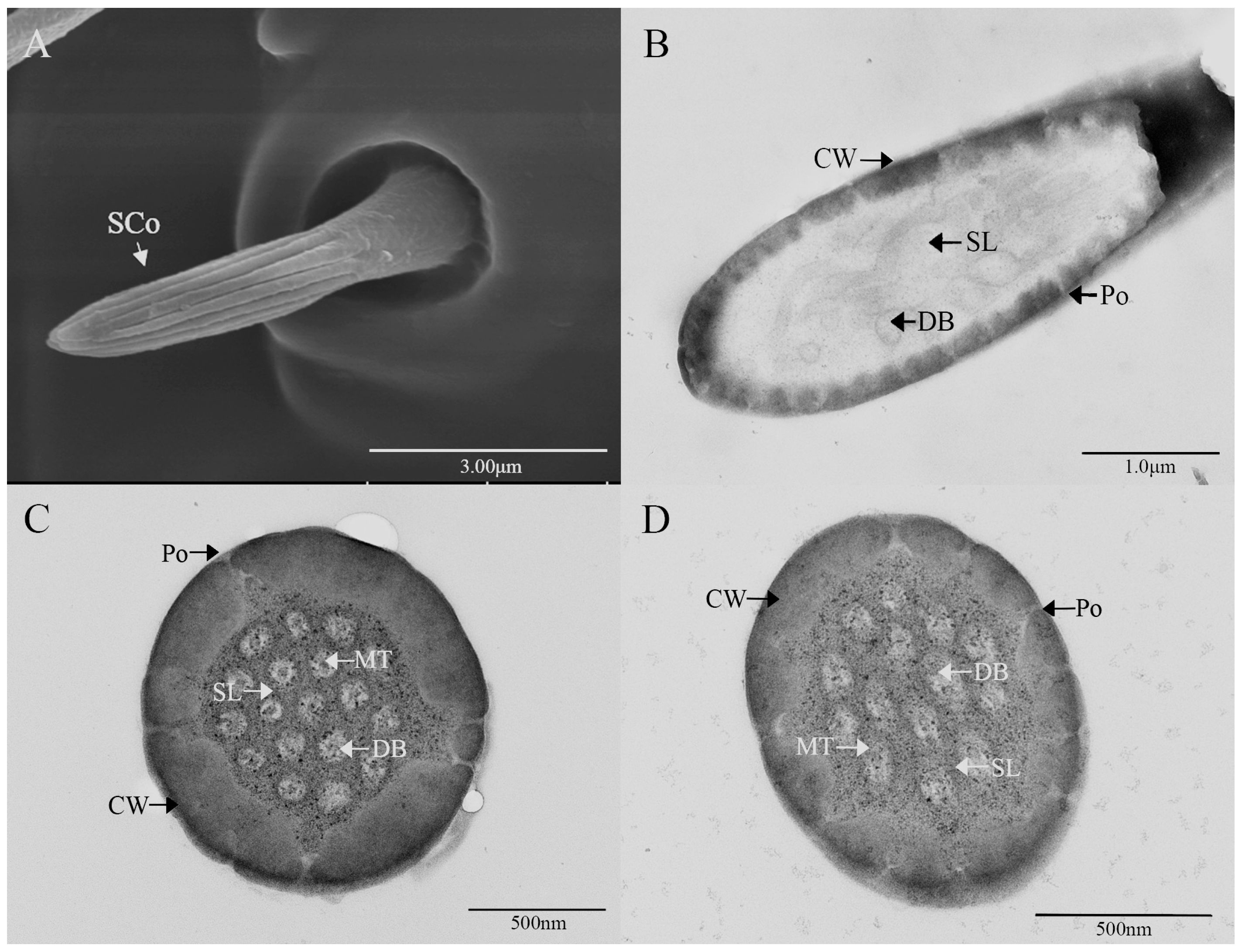
3.2. Ultrastructure of Antennal Olfactory Sensilla
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| MDPI | Multidisciplinary Digital Publishing Institute |
| DOAJ | Directory of open access journals |
| TLA | Three-letter acronym |
| LD | Linear dichroism |
| BB | Böhm bristles |
| SAu | sensilla auricillica |
| SB (I and II) | sensilla basiconca (I and II) |
| SC | sensilla chaetica |
| SCa | sensilla campaniformia |
| SCo | sensilla coeloconica |
| ST (I and II) | sensilla trichodea (I and II) |
| CW | cuticular wall |
| DB | dendritic branches |
| MT | microtubules |
| Po | Pore |
| SL | lymph space |
References
- Zhang, C.; Yuan, Z.H.; Wang, Z.Y.; He, K.L.; Bai, S.X. Population dynamics of Monolepta hieroglyphica (Motschulsky) in cornfields. Chin. J. Appl. Entomol. 2014, 51, 668–675. [Google Scholar]
- Yu, P.Y.; Wang, S.Y.; Yang, X.K. Economic Entomology of China: Volume 54, Coleptera: Chrysomeloidea(II); Science Press: Beijing, China, 1996; p. 169. [Google Scholar]
- Liu, Q.; Zhao, J.; Hu, C.; Ma, J.; Deng, C.; Ma, L.; Qie, X.; Yuan, X.; Yan, X. Predicting the Current and Future Distribution of Monolepta signata (Coleoptera: Chrysomelidae) Based on the Maximum Entropy Model. Insects 2024, 15, 575. [Google Scholar] [CrossRef] [PubMed]
- Romantsov, P.V. On the Knowledge of the Leaf-Beetle Fauna (Coleoptera: Chrysomelidae) of Siberia and the Russian Far East. Entomol. Rev. 2021, 101, 97–120. [Google Scholar] [CrossRef]
- Research Group of Leaf Beetle, Division of Insect Taxonomy; Institute of Zoology, Academia Sinica; Plant Protection Group, Institute of Agriculture of Baxia; Changchiakou District, Hopei Province; Technical Station, Plant Protection Station of Xiheying People’s Commune; Agricultural Bureau of Yu County, Hopei Province. Brief report on the study of Firefly bimaculate. Acta Entomol. Sin. 1979, 1, 115–117. [Google Scholar]
- Shi, R.; Hao, J.; Zhang, Y.; Wang, Q.; Liu, C.; Yang, Q. Impact of Different Temperatures on Activity of the Pest Monolepta hieroglyphica Motschulsky (Coleoptera: Chrysomelidae). Insects 2025, 16, 222. [Google Scholar] [CrossRef]
- Ramzan, M.; Khashaveh, A.; Yi, C.; Shan, S.; Tang, Z.; Liu, W.; Khan, K.A.; Zhang, Y. Biology, ecology, host range, and management approaches for Monolepta spp.(Coleoptera: Chrysomelidae), emerging threats to crops. Int. J. Pest Manag. 2024, 15, 24. [Google Scholar] [CrossRef]
- Han, H.; Zhang, H.; Dang, R.H.; Han, R.; Hao, S.K. Occurrence and comprehensive prevention and control measures of maize double-spotted firefly in Weinan City. Shanxi Agric. Sci. 2013, 59, 82–83. [Google Scholar]
- Gao, Y.; Xu, F.X.; Li, H.Y.; Wang, D.; Shi, S.S. Research progress on comprehensive management of Hotartaria bimaculata in soybean. Soybean Sci. Technol. 2018, 4, 43–46. [Google Scholar]
- Peter, J.D.; Ryan, M.F. Ultrastructure of antennal sensilla of Nebria brevicollis (Fab.) (Coleoptera: Carabidae). Int. J. Insect Morphol. Embryol. 1979, 8, 169–181. [Google Scholar]
- Saïd, I.; Tauban, D.; Renou, M.; Mori, K.; Rochat, D. Structure and function of the antennal sensilla of the palm weevil Rhynchophorus palmarum (Coleoptera: Curculionidae). J. Insect Physiol. 2003, 49, 857–872. [Google Scholar] [CrossRef]
- Urbanek, A.; Łuszczek, D.; Kapusta, M. Antennal and mouthpart sensilla of Acanthoscelides obtectus (Say) (Coleoptera: Bruchidae). Microsc. Res. Tech. 2016, 79, 1230–1235. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zheng, H.; Zhang, Y.; Zhang, X. Morphology and distribution of antennal, maxillary palp and labial palp sensilla of the adult bruchid beetles, Callosobruchus chinensis (L.) (Coleoptera: Bruchidae). Entomol. Res. 2018, 48, 466–479. [Google Scholar] [CrossRef]
- Liu, Z.; Hu, T.; Guo, H.W.; Liang, X.F.; Cheng, Y.Q. Ultrastructure of the olfactory sensilla across the antennae and maxillary palps of Bactrocera dorsalis (Diptera: Tephritidae). Insects 2021, 12, 289. [Google Scholar] [CrossRef] [PubMed]
- Catalá, S.S. Sensilla associated with the rostrum of eight species of Triatominae. J. Morphol. 1996, 228, 195–201. [Google Scholar] [CrossRef]
- Kerkut, G.A.; Gilbert, L. Comprehensive Insect Physiology, Biochemistry, and Pharmacology, 1st ed.; Pergamon Press: Oxford, UK, 1985. [Google Scholar]
- Steinbrecht, R.A. Pore structures in insect olfactory sensilla: A review of data and concepts. Insect Morphol. Embryol. 1997, 26, 229–245. [Google Scholar] [CrossRef]
- Barbarossa, I.T.; Muroni, P.; Dardani, M.; Casula, P.; Angioy, A.M. New insight into the antennal chemosensory function of Opius concolor (Hymenoptera, Braconidae). Ital. J. Zool. 1998, 65, 367–370. [Google Scholar] [CrossRef]
- Zhang, C.; He, K.L.; Bai, S.X. Scanning electron microscopy studies of antennal sensilla of Monolepta hieroglyphica. Chin. J. Appl. Entomology 2012, 49, 756–761. [Google Scholar]
- Pregitzer, P.; Schultze, A.; Raming, K.; Breer, H.; Krieger, J. Expression of a GABAB-receptor in olfactory sensory neurons of sensilla trichodea on the male antennae of the moth Heliothis virescens. Int. J. Biol. Sci. 2013, 9, 707. [Google Scholar] [CrossRef]
- Wei, J.; Zhou, Q.; Hall, L.; Myrick, A.; Hoover, K.; Shields, K.; Baker, T.C. Olfactory sensory neurons of the Asian longhorned beetle, Anoplophora glabripennis, specifically responsive to its two aggregation-sex pheromone components. J. Chem. Ecol. 2018, 44, 637–649. [Google Scholar] [CrossRef]
- Guo, D.D. Ultrastructure of the Head of Monolepta hieroglyphica (Motschulsky) and its Response to Host Volatile Compounds. Master’s Thesis, Shihezi University, Shihezi, China, 2018. [Google Scholar]
- Zhang, Z.H.; Chen, J.; Tang, S.Q.; Zhang, J.; Li, L. Olfactory Behavioral Response of the Monolepta hieroglyphica (Motschulsky) to volatiles of cotton and corn such as Dragosantol. Xinjiang Agric. Sci. 2018, 55, 285–292. [Google Scholar]
- Li, L.; Yang, C.; Chen, J. Identification of Active Components in Feces of Monolepta hieroglyphica (Motschulsky). Xinjiang Agric. Sci. 2020, 57, 1103–1112. [Google Scholar]
- He, W.J.; Meng, H.Y.; Zhi, M.T.; Chen, J. Analysis of antennal transcriptome and differentially expressed genes of female and male Monolepta signata. Xinjiang Agric. Sci. 2024, 61, 984–995. [Google Scholar]
- He, W.J.; Meng, H.Y.; Zhang, Y.; Zhang, G.; Zhi, M.Y.; Li, G.Y.; Chen, J. Identification of candidate chemosensory genes in the antennal transcriptome of Monolepta signata. PLoS ONE 2024, 19, e0301177. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.M.; Ling, B.; Zhao, X.W.; Miao, X.X.; Zhang, M.X. Ultrastructure of antennal sensilla of Phyllotreta striolata Fabricius (Coleoptera: Alticinae). J. Environ. Entomol. 2015, 37, 68–76. [Google Scholar]
- Yu, T.H.; Wang, L.M.; Tan, J.Z.; Li, X.S.; Song, C.; Chen, Z.L. Study on Ultrastructure of Antennal Sensilla of Calosoma maximoviczi Morawitz. Liaoning Agric. Sci. 2020, 6, 1–6. [Google Scholar]
- Keil, T.A.; Steinbrecht, R.A. Diffusion barriers in silkmoth sensory epithelia: Application of lanthanum tracer to olfactory sensilla of Antheraea polyphemus and Bombyx mori. Tissue Cell 1987, 19, 119–134. [Google Scholar] [CrossRef]
- Ghabeish, I.; Allawi, T. Agromyzid Leafminers and their Parasitoids in Jordan. Dirasat Agric. Sci. 2001, 28, 172–177. [Google Scholar]
- Forstner, M.; Breer, H.; Krieger, J. A receptor and binding protein interplay in the detection of a distinct pheromone component in the silkmoth Antheraea polyphemus. Int. J. Biol. Sci. 2009, 5, 745–757. [Google Scholar] [CrossRef]
- Merive, E.; Rahi, M.; Luik, A. Antennal sensilla of the click beetle, Melanotus villosus (Geoffroy) (Coleoptera: Elateridae). Int. J. Insect Morphol. Embryol. 1999, 28, 41–51. [Google Scholar] [CrossRef]
- Ali, S.A.; Diakite, M.M.; Ali, S.; Wang, M.-Q. Effects of the antennal sensilla distribution pattern on the behavioral responses of Tribolium castaneum (Coleoptera: Tenebrionidae). Fla. Entomol. 2016, 99, 52–59. [Google Scholar] [CrossRef]
- Gao, Z.Z.; Wu, W.J.; Liang, G.W. Observation of the antennal sensilla of Campylomma chinensis (Hemiptera: Miridae) by environmental scanning electron microscope. J. South China Agric. 2006, 27, 18–20. [Google Scholar]
- Vogt, R.G.; Rogers, M.E.; Franco, M.D.; Sun, M. A comparative study of odorant binding protein genes: Differential expression of the PBP1-GOBP2 gene cluster in Manduca sexta (Lepidoptera) and the organization of OBP genes in Drosophila melanogaster (Diptera). J. Exp. Biol. 2002, 205, 719–744. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.X.; Yang, B.; Sun, D.D.; Guo, M.B.; Zhang, J.; Wang, G.R. Ionotropic receptor 8a is involved in the attraction of Helicoverpa armigera to acetic acid. Insect Sci. 2022, 29, 657–668. [Google Scholar] [CrossRef] [PubMed]
- Pophof, B. Olfactory responses recorded from sensilla coeloconica of the silkmoth Bombyx mori. Physiol. Entomol. 1997, 22, 239–248. [Google Scholar] [CrossRef]
- Guo, M.; Krieger, J.; Große-Wilde, E.; Mißbach, C.; Zhang, L.; Breer, H. Variant Ionotropic Receptors Are Expressed in Olfactory Sensory Neurons of Coeloconic Sensilla on the Antenna of the Desert Locust (Schistocerca gregaria). Int. J. Biol. Sci. 2014, 10, 1–14. [Google Scholar] [CrossRef]
- Jiang, X.; Breer, H.; Pregitzer, P. Sensilla-Specific Expression of Odorant Receptors in the Desert Locust Schistocerca gregaria. Front. Physiol. 2019, 10, 1052. [Google Scholar] [CrossRef]
- You, Y.; Smith, D.P.; Lv, M.; Zhang, L. A broadly tuned odorant receptor in neurons of trichoid sensilla in locust, Locusta migratoria. Insect Biochem. Mol. Biol. 2016, 79, 66–72. [Google Scholar] [CrossRef]
- Al-Jalely Basman, H.; Xu, W. Olfactory Sensilla and Olfactory Genes in the Parasitoid Wasp Trichogramma pretiosum Riley (Hymenoptera: Trichogrammatidae). Insects 2021, 12, 998. [Google Scholar] [CrossRef]
- Lv, J.Y.; Meng, Z.J. The Olfactory Recognition Mechanism of Herbivore Insects on Plant Volatiles: A Review. Chin. Agric. Sci. Bull. 2022, 38, 122–129. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, J.; He, W.; Li, H.; Zhu, J.; Li, X.; Tian, J.; Luo, M.; Chen, J. The Ultrastructure of Olfactory Sensilla Across the Antenna of Monolepta signata (Oliver). Insects 2025, 16, 573. https://doi.org/10.3390/insects16060573
Cao J, He W, Li H, Zhu J, Li X, Tian J, Luo M, Chen J. The Ultrastructure of Olfactory Sensilla Across the Antenna of Monolepta signata (Oliver). Insects. 2025; 16(6):573. https://doi.org/10.3390/insects16060573
Chicago/Turabian StyleCao, Jiyu, Wanjie He, Huiqin Li, Jiangyan Zhu, Xiaoge Li, Jiahui Tian, Mengdie Luo, and Jing Chen. 2025. "The Ultrastructure of Olfactory Sensilla Across the Antenna of Monolepta signata (Oliver)" Insects 16, no. 6: 573. https://doi.org/10.3390/insects16060573
APA StyleCao, J., He, W., Li, H., Zhu, J., Li, X., Tian, J., Luo, M., & Chen, J. (2025). The Ultrastructure of Olfactory Sensilla Across the Antenna of Monolepta signata (Oliver). Insects, 16(6), 573. https://doi.org/10.3390/insects16060573