
Taxonomic Revision of the Genus Miltotranes Zimmerman, 1994 (Coleoptera: Curculionidae: Molytinae), the Bowenia-Pollinating Cycad Weevils in Australia, with Description of a New Species and Implications for the Systematics of Bowenia †



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Specimen Depositories
2.2. Specimen Preparation and Photography, Measurements and Terminology
2.3. Distribution Maps
3. Results
3.1. Key to the Adults of Miltotranes Zimmerman, 1994
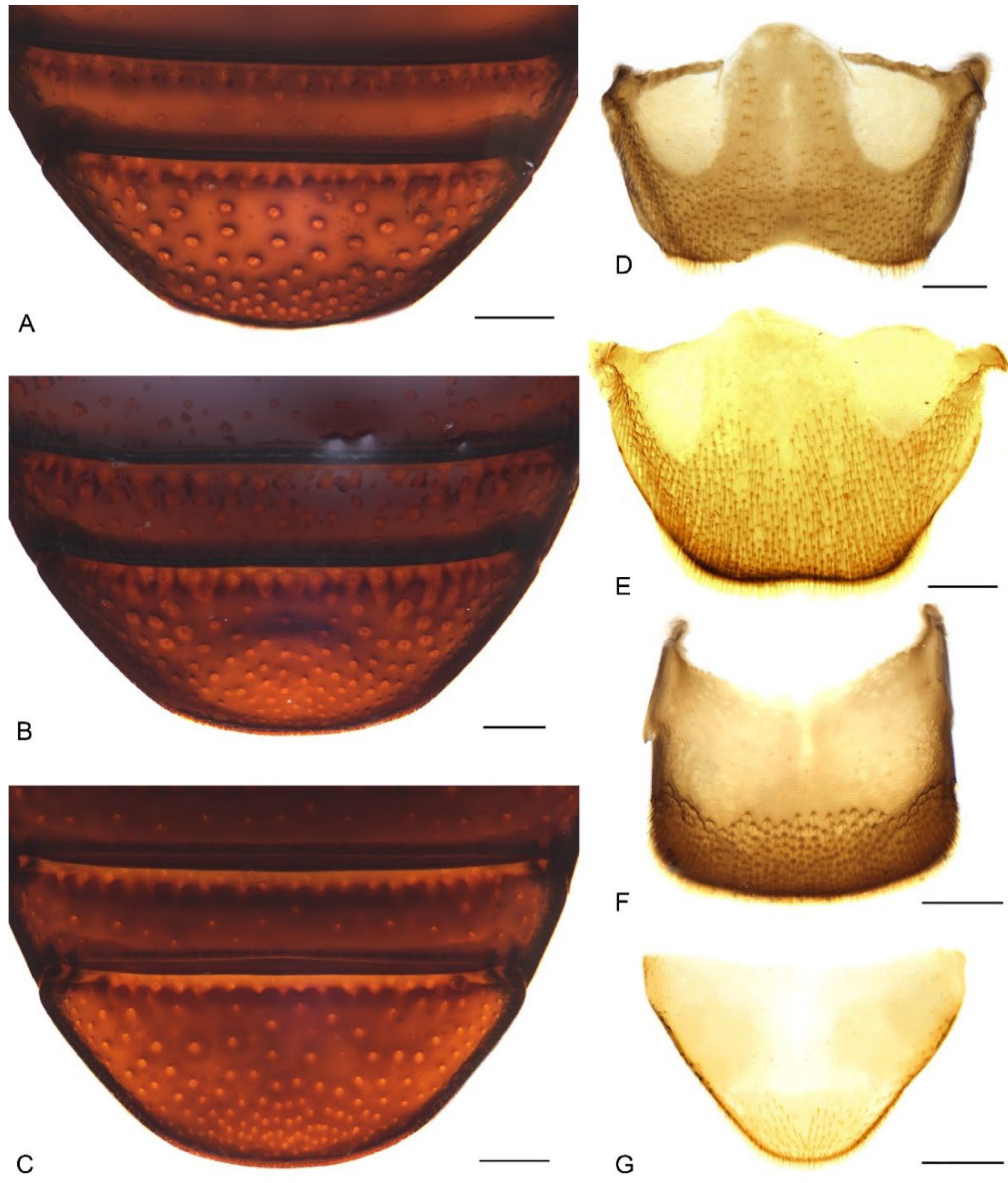
| 1. Body uniformly dark brown (Figure 1C,D); antennae inserted in middle of rostrum in male, slightly behind middle in female (Figure 1C,D and Figure 2C,D); abdominal ventrite 5 in female distinctly depressed posteriorly (Figure 7B); distribution: Byfield district of central-eastern Queensland………………………………………………………………M. subopacus |
| — Body orange to dark red with large black macula in middle of elytra (Figure 1A,B,E,F); antennae inserted slightly before middle in male, in middle in female ( Figure 1A,B,E,F and Figure 2A,B,E,F); abdominal ventrite 5 in female flat, without depression (Figure 7A,C); distribution: northern Queensland……………………………………………………………2 |
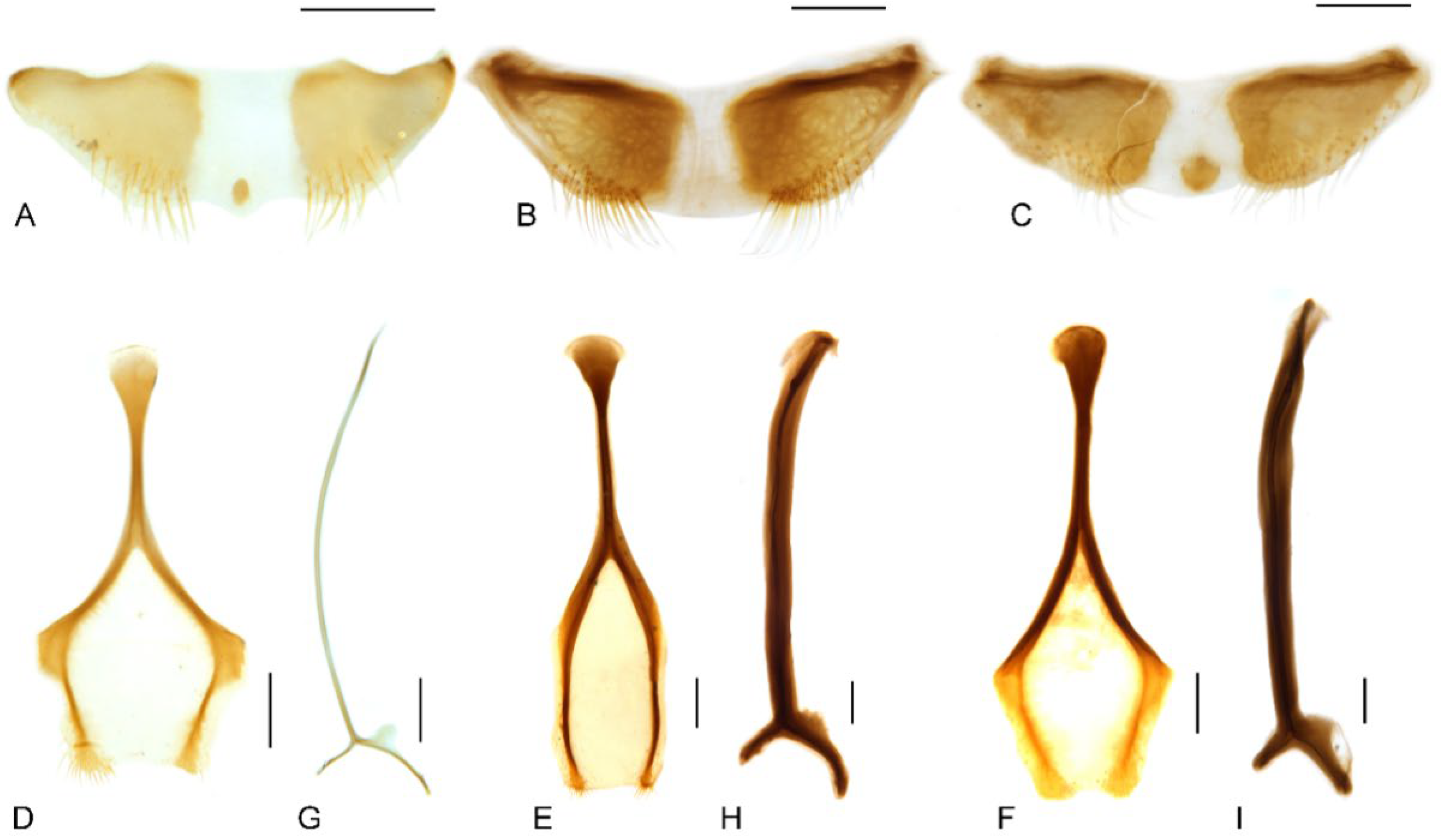
| 2. Pronotum with a pair of triangular black marks at posterior margin and black elytral macula irregular, broken and mosaic (Figure 1A,B); pronotal and elytral setae densely distributed, clustered in parts to somewhat obscure derm (Figure 1A,B and Figure 4A); pronotum ca. 0.8–0.9× as broad as elytra at humeri (Figure 1A,B); protibiae ca. 6.0× longer than wide (Figure 6A); distribution: Wet Tropics of Queensland ……………M. prosternalis |
| — Pronotum unicolorous, without black marks, and black elytral macula entire (Figure 1E,F); pronotal and elytral setae sparsely distributed (Figure 1E,F and Figure 4C); pronotum ca. 0.7–0.8× as broad as elytra at humeri (Figure 1E,F); protibiae ca. 7.5× longer than wide (Figure 6C); distribution: McIlwraith Range of Cape York Peninsula of Queensland ……………………………………………………………………………………………M. wilsoni |
3.2. Redescription of Genus
Miltotranes Zimmerman, 1994
Diagnosis
Redescription







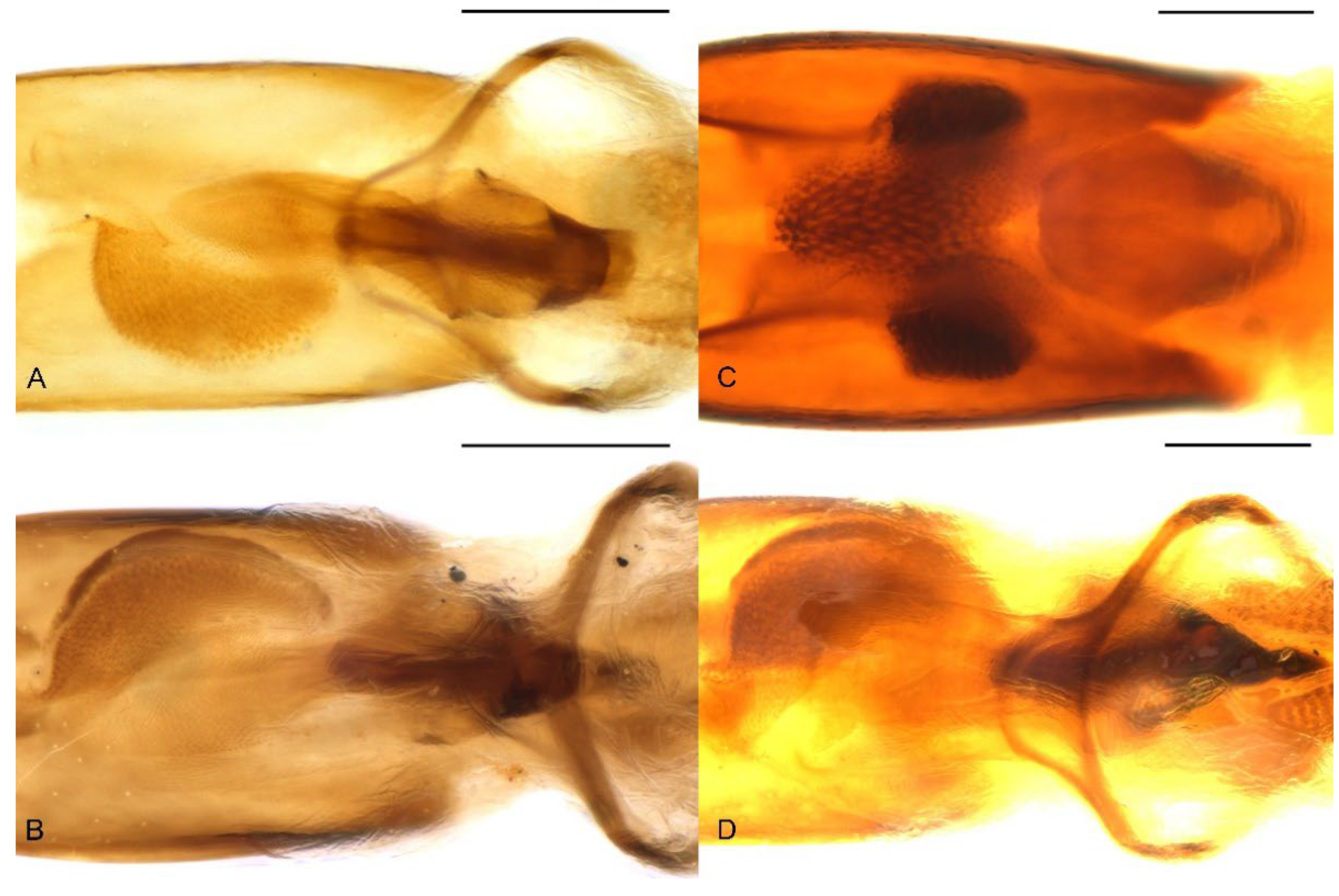
3.3. Redescription of Species
3.3.1. Miltotranes prosternalis (Lea, 1929)
Material Examined
Diagnosis
Redescription





Distribution
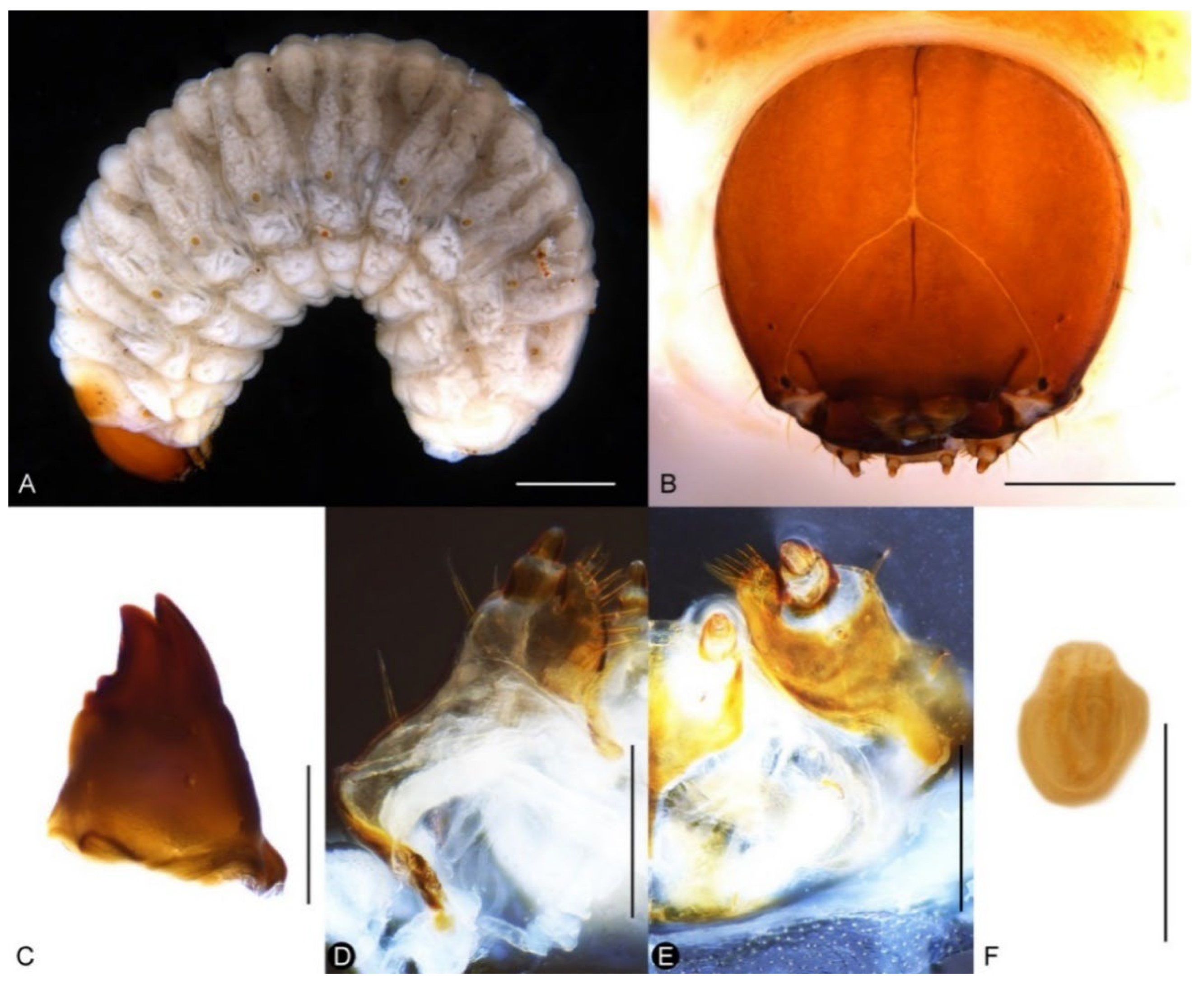
Natural History
Remarks

3.3.2. Miltotranes subopacus(Lea, 1929)
Material Examined
Diagnosis
Redescription
Distribution

Natural History
Remarks
3.3.3. Miltotranes wilsoni Hsiao & Oberprieler, sp. n.
Material Examined
Diagnosis
Redescription
Derivation of Name
Distribution
Natural History
Remarks
4. Discussion
4.1. Systematic Placement of Miltotranes and Evolution of Cycad Pollination in the Tranes Group
4.2. Implications of Miltotranes Systematics on Bowenia Taxonomy
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wilson, G.W. Initial observations of the reproductive behaviour and an insect pollination agent of Bowenia serrulata (W. Bull) Chamberlain. Encephalartos 1993, 36, 13–18. [Google Scholar]
- Wilson, G.W. Focus on Bowenia serrulata (W. Bull) Chamberlain. Encephalartos 2001, 65, 19–23. [Google Scholar]
- Wilson, G.W. Focus on Bowenia spectabilis Hook ex Hook. Encephalartos 2002, 70, 10–14. [Google Scholar]
- Wilson, G.W. Insect pollination in the cycad genus Bowenia Hook. ex Hook. f. (Stangeriaceae). Biotropica 2002, 34, 438–441. [Google Scholar]
- Wilson, G.W. The Biology and Systematics of Bowenia Hook ex. Hook f. (Stangeriaceae: Bowenioideae). Master’s (Research) Thesis, James Cook University, Cairns, Australia, 2004. [Google Scholar]
- Hsiao, Y.; Oberprieler, R.G. Bionomics and rearing of Miltotranes prosternalis (Lea, 1929) (Coleoptera: Curculionidae), a mutualistic cycad pollinator in Australia. Entomol. Sci. 2020, 23, 369–373. [Google Scholar] [CrossRef]
- Hill, K.; Osborne, R. Cycads of Australia; Kangaroo Press: Sydney, Australia, 2001. [Google Scholar]
- Jones, D.L. Cycads of the World, 2nd ed; Reed New Holland Books: Sydney, Australia, 2002. [Google Scholar]
- Hill, K.D. Bowenia serrulata. The IUCN Red List of Threatened Species 2010: E.T41979A10590781. Available online: https://www.iucnredlist.org/species/41979/10590781 (accessed on 16 April 2022).
- Hill, K.D. Bowenia spectabilis. The IUCN Red List of Threatened Species 2010: E.T41980A10591113. Available online: https://www.iucnredlist.org/species/41980/10591113 (accessed on 16 April 2022).
- Oberprieler, R.G. “Evil weevils”—the key to cycad survival and diversification? In The Biology, Structure and Systematics of the Cycadales: Proceedings of the Sixth International Conference on Cycad Biology, Nong Nooch Tropical Botanical Garden, Chonburi, Thailand, 29 July–3 August 2002; Lindstrom, A.J., Ed.; Nong Nooch Tropical Botanical Garden: Chonburi, Thailand, 2004. [Google Scholar]
- Hsiao, Y.; Oberprieler, R.G. A review of the trunk-boring cycad weevils in Australia, with description of a second species of Demyrsus Pascoe, 1872 (Coleoptera: Curculionidae). Austral. Entomol. 2020, 59, 677–700. [Google Scholar] [CrossRef]
- Oberprieler, R.G.; Anderson, R.S.; Marvaldi, A.E. Curculionoidea Latreille, 1802: Introduction, phylogeny. In Handbook of Zoology. Arthropoda: Insecta: Coleoptera, Beetles. Volume 3: Morphology and Systematics (Phytophaga); Leschen, R.A.B., Beutel, R.G., Eds.; Walter de Gruyter: Berlin, Germany, 2014; pp. 285–301. [Google Scholar]
- Zimmerman, E.C. Australian Weevils (Coleoptera: Curculionoidea). Volume I. Orthoceri. Anthribidae to Attelabidae. The Primitive Weevils; CSIRO: Melbourne, Australia, 1994.
- Oberprieler, R.G. The weevils (Coleoptera: Curculionoidea) associated with cycads. 1. Classification relationships, and biology. In Proceedings of the Third International Conference on Cycad Biology, Pretoria, South Africa, 5–9 July 1993. [Google Scholar]
- Oberprieler, R.G. The weevils (Coleoptera: Curculionoidea) associated with cycads. 2. Host specificity and implications for cycad taxonomy. In Proceedings of the Third International Conference on Cycad Biology, Pretoria, South Africa, 5–9 July 1993. [Google Scholar]
- Alonso-Zarazaga, M.A.; Lyal, C.H.C. A World Catalogue of Families and Genera of Curculionoidea (Insecta: Coleoptera) (Excepting Scolytidae and Platypodidae); Entomopraxis: Barcelona, Spain, 1999. [Google Scholar]
- Oberprieler, R.G.; Caldara, R. Siraton devillei Hustache (Coleoptera: Curculionidae), the mysterious weevil from the Isle of Elba: Exiled no longer. Zootaxa 2012, 3573, 55–58. [Google Scholar] [CrossRef]
- Lyal, C.H.C. 3.7.7 Molytinae Schoenherr, 1823. In Handbook of Zoology. Arthropoda: Insecta: Coleoptera, Beetles. Volume 3: Morphology and Systematics (Phytophaga); Leschen, R.A.B., Beutel, R.G., Eds.; Walter de Gruyter: Berlin, Germany, 2014; pp. 529–570. [Google Scholar]
- Pullen, K.R.; Jennings, D.; Oberprieler, R.G. Annotated catalogue of Australian weevils (Coleoptera: Curculionoidea). Zootaxa 2014, 3896, 1–481. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.S.; Oberprieler, R.G.; Setliff, G.P. A review of the Araucaria-associated weevils of the tribe Orthorhinini (Coleoptera: Curculionidae: Molytinae), with description of new species of Ilacuris Pascoe, 1865 and Notopissodes Zimmerman & Oberprieler, 2014 and a new genus, Kuschelorhinus Anderson & Setliff. Diversity 2018, 10, 54. [Google Scholar]
- Legalov, A.A. Annotated key to weevils of the world. Part 2. Subfamily Molytinae (Coleoptera, Curculionidae). Ukr. J. Ecol. 2018, 8, 340–350. [Google Scholar]
- Toon, A.; Terry, L.I.; Tang, W.; Walter, G.H.; Cook, L.G. Insect pollination of cycads. Austral. Ecol. 2020, 45, 1033–1058. [Google Scholar] [CrossRef]
- Lea, A.M. Descriptions of new species of Australian Coleoptera. Part XX. Proc. Linn. Soc. N. S. W. 1929, 54, 519–549. [Google Scholar]
- Schenkling, S.; Marshall, G.A.K. Curculionidae: Subfam. Amalactinae. In Coleopterorum Catalogus. Pars 150.; Junk, W., Schenkling, S., Eds.; W. Junk: Berlin, Germany, 1936; pp. 1–3. [Google Scholar]
- Hsiao, Y.; Oberprieler, R.G. A review of Paratranes Zimmerman, 1994, Xanthorrhoea-associated weevils of the Tranes group (Coleoptera, Curculionidae, Molytinae), with description of a new species. Eur. J. Taxon. 2021, 767, 117–141. [Google Scholar] [CrossRef]
- Shin, S.; Clarke, D.J.; Lemmon, A.R.; Moriarty-Lemmon, E.; Aitken, A.L.; Haddad, S.; Farrell, B.D.; Marvaldi, A.E.; Oberprieler, R.G.; McKenna, D.D. Phylogenomic data yield new and robust insights into the phylogeny and evolution of weevils. Mol. Biol. Evol. 2018, 35, 823–836. [Google Scholar] [CrossRef] [PubMed]
- Nagalingum, N.S.; Marshall, C.R.; Quental, T.B.; Rai, H.S.; Little, D.P.; Mathews, S. Recent synchronous radiation of a living fossil. Science 2011, 334, 769–799. [Google Scholar] [CrossRef] [PubMed]
- Salas-Leiva, D.E.; Meerow, A.W.; Calonje, M.; Griffith, M.P.; Francisco-Ortega, J.; Nakamura, K.; Stevenson, D.W.; Lewis, C.E.; Namoff, S. Phylogeny of the cycads based on multiple single-copy nuclear genes: Congruence of concatenated parsimony, likelihood and species tree inference methods. Ann. Bot. 2013, 112, 1263–1278. [Google Scholar] [CrossRef]
- Condamine, F.L.; Nagalingum, N.S.; Marshall, C.R.; Morlon, H. Origin and diversification of living cycads: A cautionary tale on the impact of the branching prior in Bayesian molecular dating. BMC Evol. Biol. 2015, 15, 65. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.L. Cycads of the World; Smithsonian Institution Press: Washington, DC, USA, 1993. [Google Scholar]
- Bryant, L.M.; Krosch, M.N. Lines in the land: A review of evidence for eastern Australia’s major biogeographical barriers to closed forest taxa. Biol. J. Linn. Soc. 2016, 119, 238–264. [Google Scholar] [CrossRef]
- Stevenson, D.W.; Stanberg, L.; Calonje, M.A. The world list of cycads. In Cycad Biology and Conservation: The 9th International Congress on Cycad Biology. Memoirs of the New York Botanical Garden, 117; Li, N., Stevenson, D.W., Griffith, M.P., Eds.; New York Botanical Garden Press: New York, NY, USA, 2019. [Google Scholar]
- Kokubugata, G.; Kondo, K.; Wilson, G.W.; Randall, L.M.; van der Schans, A.; Morris, D.K. Comparison of karyotype and rDNA-distribution in somatic chromosomes of Bowenia species (Stangeriaceae, Cycadales). Aust. Syst. Bot. 2000, 13, 15–20. [Google Scholar] [CrossRef]
- Kokubugata, G.; Hill, K.D.; Wilson, G.W.; Kondo, K.; Randall, L.M. A comparison of chromosome number and karyotype in somatic chromosomes of Stangeriaceae (Cycadales). Edinb. J. Bot. 2001, 58, 475–481. [Google Scholar] [CrossRef][Green Version]
- Norstog, K.; Nicholls, T.J. The Biology of the Cycads.; Cornell University Press: Ithaca, NY, USA, 1997. [Google Scholar]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsiao, Y.; Oberprieler, R.G. Taxonomic Revision of the Genus Miltotranes Zimmerman, 1994 (Coleoptera: Curculionidae: Molytinae), the Bowenia-Pollinating Cycad Weevils in Australia, with Description of a New Species and Implications for the Systematics of Bowenia. Insects 2022, 13, 456. https://doi.org/10.3390/insects13050456
Hsiao Y, Oberprieler RG. Taxonomic Revision of the Genus Miltotranes Zimmerman, 1994 (Coleoptera: Curculionidae: Molytinae), the Bowenia-Pollinating Cycad Weevils in Australia, with Description of a New Species and Implications for the Systematics of Bowenia. Insects. 2022; 13(5):456. https://doi.org/10.3390/insects13050456
Chicago/Turabian StyleHsiao, Yun, and Rolf G. Oberprieler. 2022. "Taxonomic Revision of the Genus Miltotranes Zimmerman, 1994 (Coleoptera: Curculionidae: Molytinae), the Bowenia-Pollinating Cycad Weevils in Australia, with Description of a New Species and Implications for the Systematics of Bowenia" Insects 13, no. 5: 456. https://doi.org/10.3390/insects13050456
APA StyleHsiao, Y., & Oberprieler, R. G. (2022). Taxonomic Revision of the Genus Miltotranes Zimmerman, 1994 (Coleoptera: Curculionidae: Molytinae), the Bowenia-Pollinating Cycad Weevils in Australia, with Description of a New Species and Implications for the Systematics of Bowenia. Insects, 13(5), 456. https://doi.org/10.3390/insects13050456