
Melanaphis sorghi (Hemiptera: Aphididae) Clonal Diversity in the United States and Brazil

 ,
, {kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
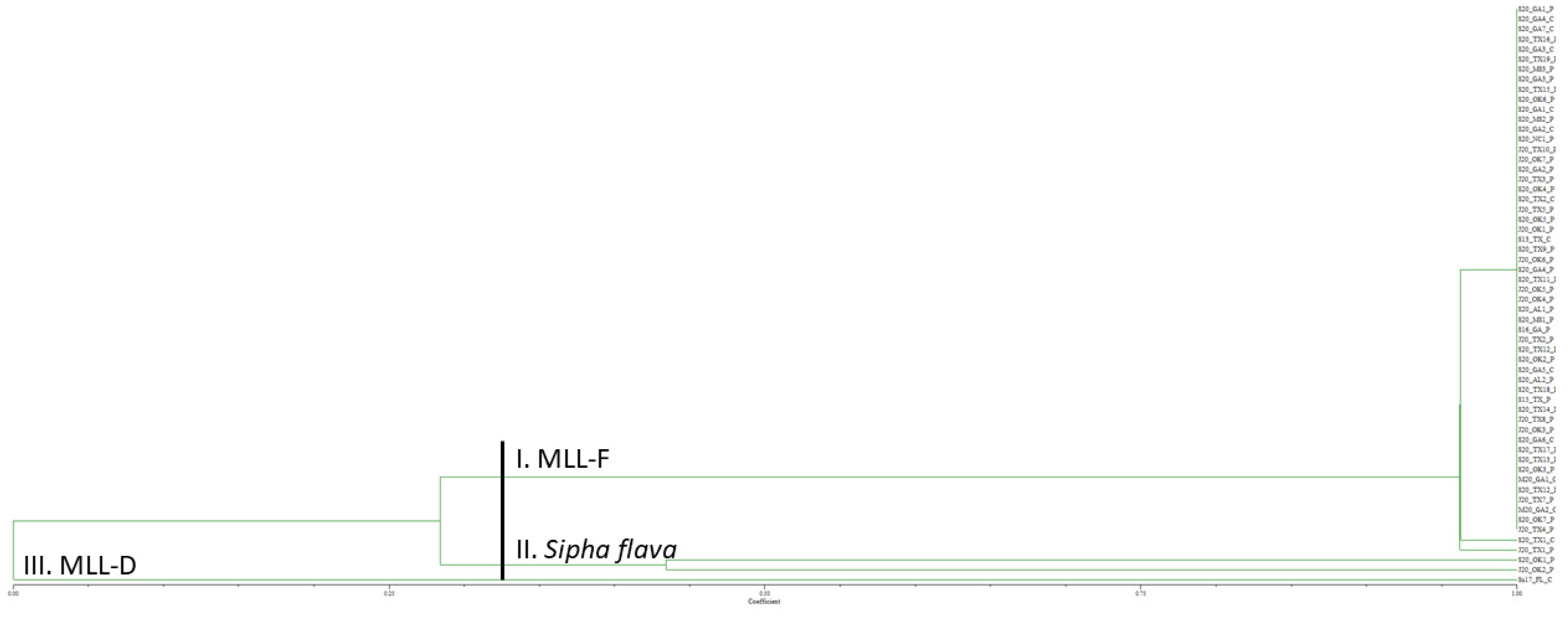
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Armstrong, J.S.; Rooney, W.L.; Peterson, G.C.; Villenueva, R.T.; Brewer, M.J.; Sekula-Ortiz, D. Sugarcane aphid (Hemiptera: Aphididae): Host range and sorghum resistance including cross-resistance from greenbug sources. J. Econ. Entomol. 2015, 108, 576–582. [Google Scholar] [CrossRef] [PubMed]
- Bowling, R.D.; Brewer, M.J.; Kerns, D.L.; Gordy, J.; Seiter, N.; Elliott, N.E.; Buntin, G.D.; Way, M.O.; Royer, T.A.; Biles, S.; et al. Sugarcane aphid (Hemiptera: Aphididae): A new pest on sorghum in North America. J. Integr. Pest Manag. 2016, 7, 12. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, J.S.; Mbulwe, L.; Sekula-Ortiz, D.; Villanueva, R.T.; Rooney, W.L. Resistance to Melanaphis sacchari (Hemiptera: Aphididae) in forage and grain sorghums. J. Econ. Entomol. 2017, 110, 259–265. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Armstrong, J.S.; Harris-Shultz, K.R.; Ni, X.; Wang, H.; Knoll, J.E.; Anderson, W.F. Utilizing biodemographic indices to identify perennial bioenergy grasses as sugarcane aphid (Hemiptera: Aphididae) host plants. Trends Entomol. 2019, 15, 1–14. [Google Scholar]
- Paudyal, S.; Armstrong, J.S.; Harris-Shultz, K.R.; Wang, H.; Giles, K.L.; Rott, P.C.; Payton, M.E. Evidence of host plant specialization among the U.S. sugarcane aphid (Hemiptera: Aphididae) genotypes. Trends Entomol. 2019, 15, 47–58. [Google Scholar]
- Brewer, M.J.; Gordy, J.W.; Kerns, D.L.; Woolley, J.B.; Rooney, W.L.; Bowling, R.D. Sugarcane aphid population growth, plant injury, and natural enemies on selected grain sorghum hybrids in Texas and Louisiana. J. Econ. Entomol. 2017, 110, 2109–2118. [Google Scholar] [CrossRef]
- Singh, B.U.; Padmaja, P.G.; Seetharama, N. Biology and management of the sugarcane aphid, Melanaphis sacchari (Zehntner) (Homoptera: Aphididae), in sorghum: A review. Crop Prot. 2004, 23, 739–755. [Google Scholar] [CrossRef]
- Brewer, M.J.; Bowling, R.; Michaud, J.P.; Jacobson, A.L. 2016, Sugarcane Aphid: A new Pest in North America. ENTO-056. Texas A&M AgriLife Extension Service, College Station, TX. Available online: https://agrilifecdn.tamu.edu/sca/files/2017/03/Sugarcane-Aphid-a-New-Sorghum-Pest-in-North-America.pdf (accessed on 18 April 2022).
- Peterson, G.C.; Armstrong, J.S.; Pendleton, B.B.; Stelter, M.; Brewer, M.J. Registration of RTx3410 through RTx3428 sorghum germplasm resistant to sugarcane aphid [Melanaphis sacchari (Zehntner)]. J. Plant Regist. 2018, 12, 391–398. [Google Scholar] [CrossRef]
- Villanueva, R.T.; Brewer, M.; Way, M.O.; Biles, S.; Sekula-Ortiz, D.; Bynum, E.; Swart, J.; Crumley, C.; Knutson, A.; Porter, P.; et al. Sugarcane Aphid: A New Pest of Sorghum. Texas A&M Agrilife Extension, Ento-035. 2014. Available online: http://denton.agrilife.org/files/2013/08/ENTO-035-The-Sugarcane-Aphid-2014.pdf (accessed on 25 April 2022).
- Nibouche, S.; Fartek, B.; Mississipi, S.; Delatte, H.; Reynaud, B.; Costet, L. Low genetic diversity in Melanaphis sacchari aphid populations at the worldwide scale. PLoS ONE 2014, 9, e106067. [Google Scholar] [CrossRef]
- Harris-Shultz, K.; Ni, X.; Wadl, P.A.; Wang, X.; Wang, H.; Huang, F.; Flanders, K.; Seiter, N.; Kerns, D.; Meagher, R.; et al. Microsatellite markers reveal a predominant sugarcane aphid (Homoptera: Aphididae) clone is found on sorghum in seven states and one territory of the USA. Crop Sci. 2017, 57, 2064–2072. [Google Scholar] [CrossRef]
- Nibouche, S.; Costet, L.; Holt, J.R.; Jacobson, A.; Pekarcik, A.; Sadeyen, J.; Armstrong, J.S.; Peterson, G.C.; McLaren, N.; Medina, R.F. Invasion of sorghum in the Americas by a new sugarcane aphid (Melanaphis sacchari) superclone. PLoS ONE 2018, 13, e0196124. [Google Scholar] [CrossRef] [PubMed]
- Harris-Shultz, K.; Ni, X. A sugarcane aphid (Hemiptera: Aphididae) ‘super-clone’ remains on U.S. sorghum and Johnsongrass and feeds on Giant Miscanthus. J. Entomol. Sci. 2021, 56, 43–52. [Google Scholar] [CrossRef]
- Nibouche, S.; Costet, L.; Medina, R.F.; Holt, J.R.; Sadeyen, J.; Zoogones, A.-S.; Brown, P.; Blackman, R.L. Morphometric and molecular discrimination of the sugarcane aphid, Melanaphis sacchari, (Zehntner, 1897) and the sorghum aphid Melanaphis sorghi (Theobald, 1904). PLoS ONE 2021, 16, e0241881. [Google Scholar] [CrossRef] [PubMed]
- Paudyal, S.; Armstrong, J.S.; Giles, K.L.; Payton, M.E.; Opit, G.P.; Limaje, A. Categories of resistance to sugarcane aphid (Hemiptera: Aphididae) among sorghum genotypes. J. Econ. Entomol. 2019, 112, 1932–1940. [Google Scholar] [CrossRef]
- MyFields. Pest Profile. 2021. Available online: https://www.myfields.info/pests/sugarcane-aphid (accessed on 18 April 2022).
- Harris-Shultz, K.; Armstrong, S.; Jacobson, A. Invasive cereal aphids of North America: Biotypes, genetic variation, management, and lessons learned. Trends Entomol. 2019, 15, 99–122. [Google Scholar]
- Harris-Shultz, K.; Ni, X.; Wang, H.; Knoll, J.E.; Anderson, W.F. Use of benzimidazole agar plates to assess fall armyworm (Lepidoptera: Noctuidae) feeding on excised maize and sorghum leaves. Florida Entomol. 2015, 98, 394–397. [Google Scholar] [CrossRef]
- Nowierski, R.M.; Zeng, Z.; Scharen, A. Age-specific life table modeling of the Russian wheat aphid (Homoptera: Aphididae) on barley grown in benzimidazole agar. Environ. Entomol. 1995, 24, 1284–1290. [Google Scholar] [CrossRef]
- Molecular Ecology Resources Primer Development Consortium; Andris, M.; Aradottir, G.I.; Arnau, G.; Audzijonyte, A.; Bess, E.C.; Bonadonna, F.; Bourdel, G.; Bried, J.; Bugbee, G.; et al. Permanent genetic resources added to Molecular Ecology Resources Database 1 June 2010–31 July 2010. Mol. Ecol. Resour. 2010, 10, 1106–1108. [Google Scholar] [CrossRef]
- Rohlf, F.J. NTSYSpc: Numerical Taxonomy System; Version 2.20; Exeter Publ.: Setauket, NY, USA, 2008. [Google Scholar]
- Nei, M.; Li, W.H. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA 1979, 76, 5269–5273. Available online: http://www.jstor.org/stable/70430 (accessed on 18 April 2022). [CrossRef]
- Hall, D.G. The sugarcane aphid, Melanaphis sacchari, in Florida sugarcane. J. Am. Soc. Sugar Cane Technol. 1987, 7, 26–29. [Google Scholar]
- White, W.H.; Reagan, T.E.; Hall, D.G. Melanaphis sacchari (Homoptera: Aphididae), a sugarcane pest new to Louisiana. Fla. Entomol. 2001, 84, 435–436. [Google Scholar] [CrossRef]
- Harrington, R. Aphid layer (letter). Antenna Bull. R. Entomol. Soc. 1994, 18, 50. [Google Scholar]
- Lushai, G.; De Barro, P.J.; David, O.; Sherratt, T.N.; Maclean, N. Genetic variation within a parthenogenetic lineage. Insect Mol. Biol. 1998, 7, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Forneck, A.; Walker, M.A.; Blaich, R. Ecological and genetic aspects of grape phylloxera (Daktulosphaira vitifoliae Fitch) performance on rootstock hosts. Bull. Entomol. Res. 2001, 91, 445–451. [Google Scholar] [CrossRef]
- Loxdale, H.D.; Lushai, G. Rapid changes in clonal lines: The death of a ‘sacred cow’. Biol. J. Linn. Soc. Lond. 2003, 79, 3–16. [Google Scholar] [CrossRef]
- Figueroa, C.C.; Fuentes-Contreras, E.; Molina-Montenegro, M.A.; Ramirez, C.C. Biological and genetic features of introduced aphid populations in agroecosystems. Curr. Opin. Insect Sci. 2018, 26, 63–68. [Google Scholar] [CrossRef]
- Colares, F.; Michaud, J.P.; Bain, C.L.; Torres, J.B. Recruitment of aphidophagous arthropods to sorghum plants infested with Melanaphis sacchari and Schizaphis graminum (Hemiptera: Aphididae). Biol. Control 2015, 90, 16–24. [Google Scholar] [CrossRef]
- Szczepaniec, A. Interactive effects of crop variety, insecticide seed treatment, and planting date on population dynamics of sugarcane aphid (Melanaphis sacchari) and their predators in late-colonized sorghum. Crop Prot. 2018, 109, 72–79. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harris-Shultz, K.; Armstrong, J.S.; Carvalho, G., Jr.; Segundo, J.P.; Ni, X. Melanaphis sorghi (Hemiptera: Aphididae) Clonal Diversity in the United States and Brazil. Insects 2022, 13, 416. https://doi.org/10.3390/insects13050416
Harris-Shultz K, Armstrong JS, Carvalho G Jr., Segundo JP, Ni X. Melanaphis sorghi (Hemiptera: Aphididae) Clonal Diversity in the United States and Brazil. Insects. 2022; 13(5):416. https://doi.org/10.3390/insects13050416
Chicago/Turabian StyleHarris-Shultz, Karen, John Scott Armstrong, Geraldo Carvalho, Jr., Jurandir Pereira Segundo, and Xinzhi Ni. 2022. "Melanaphis sorghi (Hemiptera: Aphididae) Clonal Diversity in the United States and Brazil" Insects 13, no. 5: 416. https://doi.org/10.3390/insects13050416
APA StyleHarris-Shultz, K., Armstrong, J. S., Carvalho, G., Jr., Segundo, J. P., & Ni, X. (2022). Melanaphis sorghi (Hemiptera: Aphididae) Clonal Diversity in the United States and Brazil. Insects, 13(5), 416. https://doi.org/10.3390/insects13050416