
Multimodal Information Processing and Associative Learning in the Insect Brain


{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Sensory Processing and Perception across Multiple Modalities
3. Bimodal Processing and Learning in Popular Insect Models
3.1. Honeybees (Apidae)
3.2. Ants (Formicidae)
3.3. Flies (Drosophilidae)
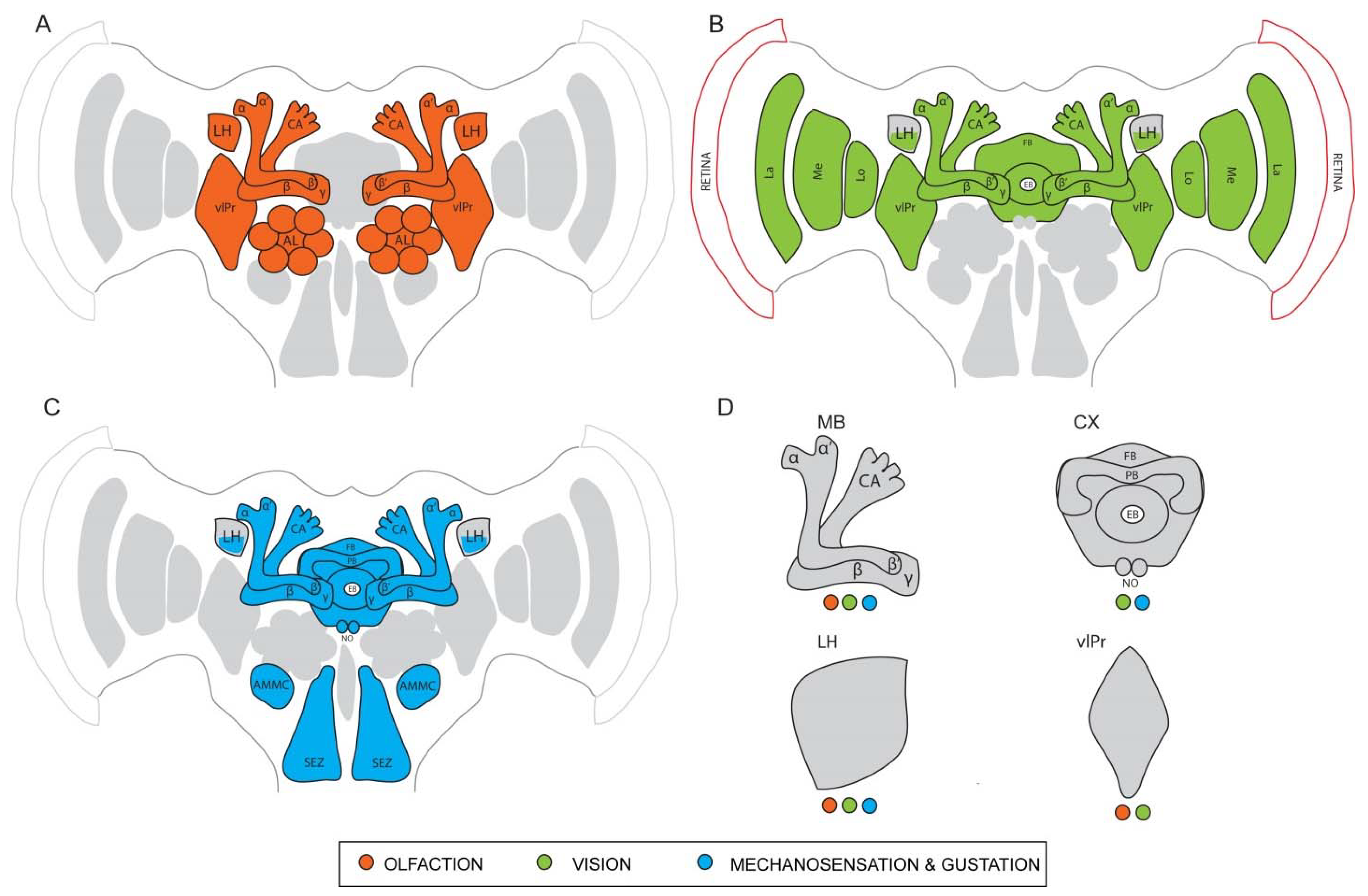
4. Neuronal Substrates and Brain Centers Underlying Unimodal and Multimodal Processing
4.1. The Insect Olfactory System—A High-End Chemosensor
4.2. The Insect Visual System—A Thousand Tiny Eyes Working as One
4.3. Gustation and Mechanosensation
4.4. The Mushroom Bodies
4.5. The Central Complex
4.6. The Lateral Horn
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Nicholas, S.; Supple, J.; Leibbrandt, R.; Gonzalez-Bellido, P.T.; Nordström, K. Integration of Small-and Wide-Field Visual Features in Target-Selective Descending Neurons of Both Predatory and Nonpredatory Dipterans. J. Neurosci. 2018, 38, 10725–10733. [Google Scholar] [CrossRef] [PubMed]
- Kulahci, I.G.; Dornhaus, A.; Papaj, D.R. Multimodal Signals Enhance Decision Making in Foraging Bumble-Bees. Proc. R. Soc. B Biol. Sci. 2008, 275, 797–802. [Google Scholar] [CrossRef] [PubMed]
- Giurfa, M. Visual Learning in Social Insects: From Simple Associations to Higher-Order Problem Solving BT-Sensory Perception: Mind and Matter; Barth, F.G., Giampieri-Deutsch, P., Klein, H.-D., Eds.; Springer: Vienna, Austria, 2012; pp. 109–133. [Google Scholar] [CrossRef]
- Fleischer, J.; Pregitzer, P.; Breer, H.; Krieger, J. Access to the Odor World: Olfactory Receptors and Their Role for Signal Transduction in Insects. Cell. Mol. Life Sci. 2018, 75, 485–508. [Google Scholar] [CrossRef] [PubMed]
- Masse, N.Y.; Turner, G.C.; Jefferis, G.S.X.E. Olfactory Information Processing in Drosophila. Curr. Biol. 2009, 19, R700–R713. [Google Scholar] [CrossRef]
- Shanbhag, S.; Müller, B.; Steinbrecht, R. Atlas of Olfactory Organs of Drosophila melanogaster 1. Types, External Organization, Innervation. Int. J. Insect Morphol. Embryol. 1999, 28, 377–397. [Google Scholar] [CrossRef]
- Avargués-Weber, A.; Deisig, N.; Giurfa, M. Visual Cognition in Social Insects. Annu. Rev. Entomol. 2011, 56, 423–443. [Google Scholar] [CrossRef]
- Stowers, J.R.; Hofbauer, M.; Bastien, R.; Griessner, J.; Higgins, P.; Farooqui, S.; Fischer, R.M.; Nowikovsky, K.; Haubensak, W.; Couzin, I.D.; et al. Virtual Reality for Freely Moving Animals. Nat. Methods 2017, 14, 995–1002. [Google Scholar] [CrossRef]
- Lafon, G.; Howard, S.R.; Paffhausen, B.H.; Avarguès-Weber, A.; Giurfa, M. Motion Cues from the Background Influence Associative Color Learning of Honey Bees in a Virtual-Reality Scenario. Sci. Rep. 2021, 11, 21127. [Google Scholar] [CrossRef]
- Kaushik, P.K.; Olsson, S.B. Using Virtual Worlds to Understand Insect Navigation for Bio-Inspired Systems. Curr. Opin. Insect Sci. 2020, 42, 97–104. [Google Scholar] [CrossRef]
- Fairhall, S.L.; MacAluso, E. Spatial Attention Can Modulate Audiovisual Integration at Multiple Cortical and Subcortical Sites. Eur. J. Neurosci. 2009, 29, 1247–1257. [Google Scholar] [CrossRef]
- Gottfried, J.A.; Dolan, R.J. The Nose Smells What the Eye Sees: Crossmodal Visual Facilitation of Human Olfactory Perception. Neuron 2003, 39, 375–386. [Google Scholar] [CrossRef]
- Wallace, M.T.; Meredith, M.A.; Stein, B.E. Integration of Multiple Sensory Modalities in Cat Cortex. Exp. Brain Res. 1992, 91, 484–488. [Google Scholar] [CrossRef] [PubMed]
- McMeniman, C.J.; Corfas, R.A.; Matthews, B.J.; Ritchie, S.A.; Vosshall, L.B. Multimodal Integration of Carbon Dioxide and Other Sensory Cues Drives Mosquito Attraction to Humans. Cell 2014, 156, 1060–1071. [Google Scholar] [CrossRef] [PubMed]
- Bailey, W.J.; Ager, E.I.; O’Brien, E.K.; Watson, D.L. Searching by Visual and Acoustic Cues among Bushcrickets (Orthoptera: Tettigoniidae): Will Females Remain Faithful to a Male Who Stops Calling? Physiol. Entomol. 2003, 28, 209–214. [Google Scholar] [CrossRef]
- Oh, S.M.; Jeong, K.; Seo, J.T.; Moon, S.J. Multisensory Interactions Regulate Feeding Behavior in Drosophila. Proc. Natl. Acad. Sci. USA 2021, 118, e2004523118. [Google Scholar] [CrossRef] [PubMed]
- Nurme, K.; Merivee, E.; Must, A.; Sibul, I.; Muzzi, M.; Di Giulio, A.; Williams, I.; Tooming, E. Responses of the Antennal Bimodal Hygroreceptor Neurons to Innocuous and Noxious High Temperatures in the Carabid Beetle, Pterostichus Oblongopunctatus. J. Insect Physiol. 2015, 81, 1–13. [Google Scholar] [CrossRef]
- Balkenius, A.; Kelber, A. Colour Preferences Influences Odour Learning in the Hawkmoth, Macroglossum Stellatarum. Naturwissenschaften 2006, 93, 255–258. [Google Scholar] [CrossRef]
- Raguso, R.A.; Willis, M.A. Synergy between Visual and Olfactory Cues in Nectar Feeding by Naïve Hawkmoths, Manduca Sexta. Anim. Behav. 2002, 64, 685–695. [Google Scholar] [CrossRef]
- Balkenius, A.; Bisch-Knaden, S.; Hansson, B. Interaction of Visual and Odour Cues in the Mushroom Body of the Hawkmoth Manduca Sexta. J. Exp. Biol. 2009, 212, 535–541. [Google Scholar] [CrossRef]
- Raguso, R.A. Flowers as Sensory Billboards: Progress towards an Integrated Understanding of Floral Advertisement. Curr. Opin. Plant Biol. 2004, 7, 434–440. [Google Scholar] [CrossRef]
- Cognigni, P.; Felsenberg, J.; Waddell, S. Do the Right Thing: Neural Network Mechanisms of Memory Formation, Expression and Update in Drosophila. Curr. Opin. Neurobiol. 2018, 49, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Aso, Y.; Hattori, D.; Yu, Y.; Johnston, R.M.; Iyer, N.A.; Ngo, T.T.B.; Dionne, H.; Abbott, L.F.; Axel, R.; Tanimoto, H.; et al. The Neuronal Architecture of the Mushroom Body Provides a Logic for Associative Learning. Elife 2014, 3, e04577. [Google Scholar] [CrossRef]
- Fiala, A. Olfaction and Olfactory Learning in Drosophila: Recent Progress. Curr. Opin. Neurobiol. 2007, 17, 720–726. [Google Scholar] [CrossRef] [PubMed]
- De Belle, J.S.; Heisenberg, M. Associative Odor Learning in Drosophila Abolished by Chemical Ablation of Mushroom Bodies. Science 1994, 263, 692–695. [Google Scholar] [CrossRef] [PubMed]
- Heisenberg, M. Mushroom Body Memoir: From Maps to Models. Nat. Rev. Neurosci. 2003, 4, 266–275. [Google Scholar] [CrossRef] [PubMed]
- Buatois, A.; Pichot, C.; Schultheiss, P.; Sandoz, J.C.; Lazzari, C.R.; Chittka, L.; Avarguès-Weber, A.; Giurfa, M. Associative Visual Learning by Tethered Bees in a Controlled Visual Environment. Sci. Rep. 2017, 7, 12903. [Google Scholar] [CrossRef]
- Schnaitmann, C.; Vogt, K.; Triphan, T.; Tanimoto, H. Appetitive and Aversive Visual Learning in Freely Moving Drosophila. Front. Behav. Neurosci. 2010, 4, 10. [Google Scholar] [CrossRef]
- Witthöft, W. Absolute Anzahl Und Verteilung Der Zellen Im Him Der Honigbiene. Z. Morphol. Tiere 1967, 61, 160–184. [Google Scholar] [CrossRef]
- Menzel, R. The Honeybee as a Model for Understanding the Basis of Cognition. Nat. Rev. Neurosci. 2012, 13, 758–768. [Google Scholar] [CrossRef]
- Sun, X.; Yue, S.; Mangan, M. How the Insect Central Complex Could Coordinate Multimodal Navigation. Elife 2021, 10, e73077. [Google Scholar] [CrossRef]
- Kantsa, A.; Raguso, R.A.; Dyer, A.G.; Sgardelis, S.P.; Olesen, J.M.; Petanidou, T. Community-Wide Integration of Floral Colour and Scent in a Mediterranean Scrubland. Nat. Ecol. Evol. 2017, 1, 1502–1510. [Google Scholar] [CrossRef] [PubMed]
- Kunze, J.; Gumbert, A. The Combined Effect of Color and Odor on Flower Choice Behavior of Bumble Bees in Flower Mimicry Systems. Behav. Ecol. 2001, 12, 447–456. [Google Scholar] [CrossRef]
- Balamurali, G.S.; Rose, S.; Somanathan, H.; Kodandaramaiah, U. Complex Multi-Modal Sensory Integration and Context Specificity in Colour Preferences of a Pierid Butterfly. J. Exp. Biol. 2020, 223, jeb223271. [Google Scholar] [CrossRef] [PubMed]
- Leonard, A.S.; Dornhaus, A.; Papaj, D.R. Flowers Help Bees Cope with Uncertainty: Signal Detection and the Function of Floral Complexity. J. Exp. Biol. 2011, 214, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Westerman, E.L.; Monteiro, A. Odour Influences Whether Females Learn to Prefer or to Avoid Wing Patterns of Male Butterflies. Anim. Behav. 2013, 86, 1139–1145. [Google Scholar] [CrossRef]
- Leonard, A.S.; Masek, P. Multisensory Integration of Colors and Scents: Insights from Bees and Flowers. J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 2014, 200, 463–474. [Google Scholar] [CrossRef]
- Gerber, B.; Smith, B.H. Visual Modulation of Olfactory Learning in Honeybees. J. Exp. Biol. 1998, 201, 2213–2217. [Google Scholar] [CrossRef]
- Couvillon, P.A.; Mateo, E.T.; Bitterman, M.E. Reward and Learning in Honeybees: Analysis of an Overshadowing Effect. Anim. Learn. Behav. 1996, 24, 19–27. [Google Scholar] [CrossRef][Green Version]
- Funayama, E.S.; Couvillon, P.A.; Bitterman, M.E. Compound Conditioning in Honeybees: Blocking Tests of the Independence Assumption. Anim. Learn. Behav. 1995, 23, 429–437. [Google Scholar] [CrossRef][Green Version]
- Mota, T.; Giurfa, M.; Sandoz, J.C. Color Modulates Olfactory Learning in Honeybees by an Occasion-Setting Mechanism. Learn. Mem. 2011, 18, 144–155. [Google Scholar] [CrossRef]
- Zhang, L.Z.; Zhang, S.W.; Wang, Z.L.; Yan, W.Y.; Zeng, Z.J. Cross-Modal Interaction between Visual and Olfactory Learning in Apis cerana. J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 2014, 200, 899–909. [Google Scholar] [CrossRef] [PubMed]
- Becker, M.C.; Rössler, W.; Strube-Bloss, M.F. UV Light Perception Is Modulated by the Odour Element of an Olfactory–Visual Compound in Restrained Honeybees. J. Exp. Biol. 2019, 222, jeb201483. [Google Scholar] [CrossRef] [PubMed]
- Devaud, J.M.; Papouin, T.; Carcaud, J.; Sandoz, J.C.; Grönewald, B.; Giurfa, M. Neural Substrate for Higher-Order Learning in an Insect: Mushroom Bodies Are Necessary for Configural Discriminations. Proc. Natl. Acad. Sci. USA 2015, 112, E5854–E5862. [Google Scholar] [CrossRef] [PubMed]
- Mansur, B.E.; Rodrigues, J.R.V.; Mota, T. Bimodal Patterning Discrimination in Harnessed Honey Bees. Front. Psychol. 2018, 9, 1529. [Google Scholar] [CrossRef]
- Zhang, S.W.; Lehrer, M.; Srinivasan, M.V. Honeybee Memory: Navigation by Associative Grouping and Recall of Visual Stimuli. Neurobiol. Learn. Mem. 1999, 72, 180–201. [Google Scholar] [CrossRef]
- Srinivasan, M.V.; Zhang, S.W.; Zhu, H. Honeybees Link Sights to Smells. Nature 1998, 396, 637–638. [Google Scholar] [CrossRef]
- Reinhard, J.; Srinivasan, M.V.; Guez, D.; Zhang, S.W. Floral Scents Induce Recall of Navigational and Visual Memories in Honeybees. J. Exp. Biol. 2004, 207, 4371–4381. [Google Scholar] [CrossRef]
- Giurfa, M. Learning and Cognition in Insects. Wiley Interdiscip. Rev. Cogn. Sci. 2015, 6, 383–395. [Google Scholar] [CrossRef]
- Plath, J.A.; Entler, B.V.; Kirkerud, N.H.; Schlegel, U.; Galizia, C.G.; Barron, A.B. Different Roles for Honey Bee Mushroom Bodies and Central Complex in Visual Learning of Colored Lights in an Aversive Conditioning Assay. Front. Behav. Neurosci. 2017, 11, 98. [Google Scholar] [CrossRef]
- Freas, C.A.; Schultheiss, P. How to Navigate in Different Environments and Situations: Lessons from Ants. Front. Psychol. 2018, 9, 841. [Google Scholar] [CrossRef]
- Schwarz, S.; Wystrach, A.; Cheng, K. Ants’ Navigation in an Unfamiliar Environment Is Influenced by Their Experience of a Familiar Route. Sci. Rep. 2017, 7, 14161. [Google Scholar] [CrossRef] [PubMed]
- Wystrach, A.; Mangan, M.; Webb, B. Optimal Cue Integration in Ants. Proc. R. Soc. B Biol. Sci. 2015, 282, 20151484. [Google Scholar] [CrossRef] [PubMed]
- Zeil, J.; Narendra, A.; Stürzl, W. Looking and Homing: How Displaced Ants Decide Where to Go. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20130034. [Google Scholar] [CrossRef] [PubMed]
- Collett, T.S.; Collett, M. Path Integration in Insects. Curr. Opin. Neurobiol. 2000, 10, 757–762. [Google Scholar] [CrossRef]
- Andel, D.; Wehner, R. Path Integration in Desert Ants, Cataglyphis: How to Make a Homing Ant Run Away from Home. Proc. R. Soc. B Biol. Sci. 2004, 271, 1485–1489. [Google Scholar] [CrossRef]
- Lent, D.D.; Graham, P.; Collett, T.S. Visual Scene Perception in Navigating Wood Ants. Curr. Biol. 2013, 23, 684–690. [Google Scholar] [CrossRef]
- Buehlmann, C.; Woodgate, J.L.; Collett, T.S. On the Encoding of Panoramic Visual Scenes in Navigating Wood Ants. Curr. Biol. 2016, 26, 2022–2027. [Google Scholar] [CrossRef]
- Schmid-Hempel, P.; Schmid-Hempel, R. Life Duration and Turnover of Foragers in the Ant Cataglyphis bicolor (Hymenoptera, Formicidae). Insectes Soc. 1984, 31, 345–360. [Google Scholar] [CrossRef]
- Narendra, A.; Si, A.; Sulikowski, D.; Cheng, K. Learning, Retention and Coding of Nest-Associated Visual Cues by the Australian Desert Ant, Melophorus bagoti. Behav. Ecol. Sociobiol. 2007, 61, 1543–1553. [Google Scholar] [CrossRef]
- Bisch-Knaden, S.; Wehner, R. Landmark Memories Are More Robust When Acquired at the Nest Site than En Route: Experiments in Desert Ants. Naturwissenschaften 2003, 90, 127–130. [Google Scholar] [CrossRef]
- Shams, L.; Seitz, A.R. Benefits of Multisensory Learning. Trends Cogn. Sci. 2008, 12, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Buehlmann, C.; Graham, P.; Hansson, B.S.; Knaden, M. Desert Ants Locate Food by Combining High Sensitivity to Food Odors with Extensive Crosswind Runs. Curr. Biol. 2014, 24, 960–964. [Google Scholar] [CrossRef] [PubMed]
- Wolf, H.; Wehner, R.; Institut, Z.; Zürich, U.; Zürich, C. Pinpointing Food Sources: Olfactory And Anemotactic Orientation In Desert Ants, Cataglyphis fortis. J. Exp. Biol. 2000, 868, 857–868. [Google Scholar] [CrossRef] [PubMed]
- Collett, M.; Cardé, R.T. Navigation: Many Senses Make Efficient Foraging Paths. Curr. Biol. 2014, 24, 362–364. [Google Scholar] [CrossRef][Green Version]
- Steck, K.; Hansson, B.S.; Knaden, M. Smells like Home: Desert Ants, Cataglyphis fortis, Use Olfactory Landmarks to Pinpoint the Nest. Front. Zool. 2009, 6, 5. [Google Scholar] [CrossRef]
- Steck, K.; Knaden, M.; Hansson, B.S. Do Desert Ants Smell the Scenery in Stereo? Anim. Behav. 2010, 79, 939–945. [Google Scholar] [CrossRef]
- Steck, K.; Hansson, B.S.; Knaden, M. Desert Ants Benefit from Combining Visual and Olfactory Landmarks. J. Exp. Biol. 2011, 214, 1307–1312. [Google Scholar] [CrossRef]
- Yan, H.; Opachaloemphan, C.; Mancini, G.; Yang, H.; Gallitto, M.; Mlejnek, J.; Leibholz, A.; Haight, K.; Ghaninia, M.; Huo, L.; et al. An Engineered Orco Mutation Produces Aberrant Social Behavior and Defective Neural Development in Ants. Cell 2017, 170, 736–747.e9. [Google Scholar] [CrossRef]
- McKenzie, S.K.; Fetter-Pruneda, I.; Ruta, V.; Kronauer, D.J.C. Transcriptomics and Neuroanatomy of the Clonal Raider Ant Implicate an Expanded Clade of Odorant Receptors in Chemical Communication. Proc. Natl. Acad. Sci. USA 2016, 113, 14091–14096. [Google Scholar] [CrossRef]
- Trible, W.; Olivos-Cisneros, L.; McKenzie, S.K.; Saragosti, J.; Chang, N.C.; Matthews, B.J.; Oxley, P.R.; Kronauer, D.J.C. Orco Mutagenesis Causes Loss of Antennal Lobe Glomeruli and Impaired Social Behavior in Ants. Cell 2017, 170, 727–735.e10. [Google Scholar] [CrossRef]
- Tolwinski, N.S. Introduction: Drosophila-A Model System for Developmental Biology. J. Dev. Biol. 2017, 5, 9. [Google Scholar] [CrossRef] [PubMed]
- Cardé, R.T.; Willis, M.A. Navigational Strategies Used by Insects to Find Distant, Wind-Borne Sources of Odor. J. Chem. Ecol. 2008, 34, 854–866. [Google Scholar] [CrossRef]
- Sadanandappa, M.K.; Sathyanarayana, S.H.; Kondo, S.; Bosco, G. Neuropeptide F Signaling Regulates Parasitoid-Specific Germline Development and Egg-Laying in Drosophila. PLoS Genet. 2021, 17, e1009456. [Google Scholar] [CrossRef] [PubMed]
- Halfwerk, W.; Varkevisser, J.; Simon, R.; Mendoza, E.; Scharff, C.; Riebel, K. Toward Testing for Multimodal Perception of Mating Signals. Front. Ecol. Evol. 2019, 7, 2013–2019. [Google Scholar] [CrossRef]
- Krstic, D.; Boll, W.; Noll, M. Sensory Integration Regulating Male Courtship Behavior in Drosophila. PLoS ONE 2009, 4, e4457. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Trona, F.; Khallaf, M.A.; Schuh, E.; Knaden, M.; Hansson, B.S.; Sachse, S. Electrical Synapses Mediate Synergism between Pheromone and Food Odors in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2017, 114, E9962–E9971. [Google Scholar] [CrossRef]
- Griffith, L.C.; Ejima, A. Multimodal Sensory Integration of Courtship Stimulating Cues in Drosophila melanogaster: Contextual Effects on Chemosensory Cues. Ann. N. Y. Acad. Sci. 2009, 1170, 394–398. [Google Scholar] [CrossRef]
- Ohyama, T.; Schneider-Mizell, C.M.; Fetter, R.D.; Aleman, J.V.; Franconville, R.; Rivera-Alba, M.; Mensh, B.D.; Branson, K.M.; Simpson, J.H.; Truman, J.W.; et al. A Multilevel Multimodal Circuit Enhances Action Selection in Drosophila. Nature 2015, 520, 633–639. [Google Scholar] [CrossRef]
- Gepner, R.; Skanata, M.M.; Bernat, N.M.; Kaplow, M.; Gershow, M. Computations Underlying Drosophila Photo- Taxis, Odor-Taxis, and Multi-Sensory Integration. Elife 2015, 4, e6229. [Google Scholar] [CrossRef]
- Gershow, M.; Berck, M.; Mathew, D.; Luo, L.; Kane, E.A.; Carlson, J.R.; Samuel, A.D.T. Controlling Airborne Cues to Study Small Animal Navigation. Nat. Methods 2012, 9, 290–296. [Google Scholar] [CrossRef]
- Gomez-Marin, A.; Louis, M. Multilevel Control of Run Orientation in Drosophila Larval Chemotaxis. Front. Behav. Neurosci. 2014, 8, 38. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Marin, A.; Stephens, G.J.; Louis, M. Active Sampling and Decision Making in Drosophila Chemotaxis. Nat. Commun. 2011, 2, 441. [Google Scholar] [CrossRef]
- Lehmann, F.O.; Dickinson, M.H. The Changes in Power Requirements and Muscle Efficiency during Elevated Force Production in the Fruit Fly Drosophila melanogaster. J. Exp. Biol. 1997, 200, 1133–1143. [Google Scholar] [CrossRef] [PubMed]
- Sherman, A.; Dickinson, M.H. Summation of Visual and Mechanosensory Feedback in Drosophila Flight Control. J. Exp. Biol. 2004, 207, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Currier, T.A.; Nagel, K.I. Multisensory Control of Orientation in Tethered Flying Drosophila. Curr. Biol. 2018, 28, 3533–3546.e6. [Google Scholar] [CrossRef] [PubMed]
- Frye, M.A.; Dickinson, M.H. Motor Output Reflects the Linear Superposition of Visual and Olfactory Inputs in Drosophila. J. Exp. Biol. 2004, 207, 123–131. [Google Scholar] [CrossRef]
- Chow, D.M.; Frye, M.A. Context-Dependent Olfactory Enhancement of Optomotor Flight Control in Drosophila. J. Exp. Biol. 2008, 211, 2478–2485. [Google Scholar] [CrossRef]
- Cheng, K.Y.; Colbath, R.A.; Frye, M.A. Olfactory and Neuromodulatory Signals Reverse Visual Object Avoidance to Approach in Drosophila. Curr. Biol. 2019, 29, 2058–2065.e2. [Google Scholar] [CrossRef]
- Cheng, K.Y.; Frye, M.A. Odour Boosts Visual Object Approach in Flies. Biol. Lett. 2021, 17, 20200770. [Google Scholar] [CrossRef]
- Brembs, B.; Heisenberg, M. Conditioning with Compound Stimuli in Drosophila melanogaster in the Flight Simulator. J. Exp. Biol. 2001, 204, 2849–2859. [Google Scholar] [CrossRef]
- Guo, J.; Guo, A. Neuroscience: Crossmodal Interactions between Olfactory and Visual Learning in Drosophila. Science 2005, 309, 307–310. [Google Scholar] [CrossRef] [PubMed]
- Stein, B.E.; Meredith, M.A. The Merging of the Senses. In The Merging of the Senses; The MIT Press: Cambridge, MA, USA, 1993; p. 211. [Google Scholar]
- Zhao, B.; Sun, J.; Zhang, X.; Mo, H.; Niu, Y.; Li, Q.; Wang, L.; Zhong, Y. Long-Term Memory Is Formed Immediately without the Need for Protein Synthesis-Dependent Consolidation in Drosophila. Nat. Commun. 2019, 10, 4550. [Google Scholar] [CrossRef] [PubMed]
- Tully, T.; Quinn, W.G. Classical Conditioning and Retention in Normal and Mutant Drosophila melanogaster. J. Comp. Physiol. A 1985, 157, 263–277. [Google Scholar] [CrossRef] [PubMed]
- Wessnitzer, J.; Webb, B. Multimodal Sensory Integration in Insects—Towards Insect Brain Control Architectures. Bioinspir. Biomim. 2006, 1, 63–75. [Google Scholar] [CrossRef]
- Cruse, H. What Mechanisms Coordinate Leg Movement in Walking Arthropods? Trends Neurosci. 1990, 13, 15–21. [Google Scholar] [CrossRef]
- Burdohan, J.A.; Comer, C.M. Cellular Organization of an Antennal Mechanosensory Pathway in the Cockroach, Periplaneta americana. J. Neurosci. 1996, 16, 5830–5843. [Google Scholar] [CrossRef] [PubMed]
- Homberg, U.; Christensen, T.A.; Hildebrand, J.G. Structure and Function of the Deutocerebrum in Insects. Annu. Rev. Entomol. 1989, 34, 477–501. [Google Scholar] [CrossRef]
- Rajashekhar, K.P.; Singh, R.N. Neuroarchitecture of the Tritocerebrum of Drosophila melanogaster. J. Comp. Neurol. 1994, 349, 633–645. [Google Scholar] [CrossRef]
- Berck, M.E.; Khandelwal, A.; Claus, L.; Hernandez-Nunez, L.; Si, G.; Tabone, C.J.; Li, F.; Truman, J.W.; Fetter, R.D.; Louis, M.; et al. The Wiring Diagram of a Glomerular Olfactory System. Elife 2016, 5, e14859. [Google Scholar] [CrossRef]
- Wei, H.; Lam, K.C.; Kazama, H. Odors Drive Feeding through Gustatory Receptor Neurons in Drosophila. bioRxiv 2022. [Google Scholar] [CrossRef]
- Brand, H.; Perrimon, N. Targeted Gene Expression as a Means of Altering Cell Fates and Generating Dominant Phenotypes. Development 1993, 118, 401–415. [Google Scholar] [CrossRef] [PubMed]
- Fenno, L.; Yizhar, O.; Deisseroth, K. The Development and Application of Optogenetics. Annu. Rev. Neurosci. 2011, 34, 389–412. [Google Scholar] [CrossRef] [PubMed]
- Luan, H.; Diao, F.; Scott, R.L.; White, B.H. The Drosophila Split Gal4 System for Neural Circuit Mapping. Front. Neural Circuits 2020, 14, 72. [Google Scholar] [CrossRef]
- Li, F.; Lindsey, J.; Marin, E.C.; Otto, N.; Dreher, M.; Dempsey, G.; Stark, I.; Bates, A.S.; Pleijzier, M.W.; Schlegel, P.; et al. The Connectome of the Adult Drosophila Mushroom Body Provides Insights into Function. Elife 2020, 9, e62576. [Google Scholar] [CrossRef]
- Scheffer, L.; Xu, C.S.; Januszewski, M.; Lu, Z.; Takemura, S.; Hayworth, K.; Huang, G.; Shinomiya, K.; Maitin-Shepard, J.; Berg, S.; et al. A Connectome and Analysis of the Adult Drosophila Central Brain. bioRxiv 2020, 1–83. [Google Scholar] [CrossRef]
- Dolan, M.J.; Frechter, S.; Bates, A.S.; Dan, C.; Huoviala, P.; Roberts, R.J.; Schlegel, P.; Dhawan, S.; Tabano, R.; Dionne, H.; et al. Neurogenetic Dissection of the Drosophila Lateral Horn Reveals Major Outputs, Diverse Behavioural Functions, and Interactions with the Mushroom Body. Elife 2019, 8, e43079. [Google Scholar] [CrossRef] [PubMed]
- Takemura, S.Y.; Aso, Y.; Hige, T.; Wong, A.; Lu, Z.; Xu, C.S.; Rivlin, P.K.; Hess, H.; Zhao, T.; Parag, T.; et al. A Connectome of a Learning and Memory Center in the Adult Drosophila Brain. Elife 2017, 6, e26975. [Google Scholar] [CrossRef]
- Zheng, Z.; Lauritzen, J.S.; Perlman, E.; Robinson, C.G.; Nichols, M.; Milkie, D.; Torrens, O.; Price, J.; Fisher, C.B.; Sharifi, N.; et al. A Complete Electron Microscopy Volume of the Brain of Adult Drosophila melanogaster. Cell 2018, 174, 730–743.e22. [Google Scholar] [CrossRef]
- Kreher, S.A.; Mathew, D.; Kim, J.; Carlson, J.R. Translation of Sensory Input into Behavioral Output via an Olfactory System. Neuron 2008, 59, 110–124. [Google Scholar] [CrossRef]
- Hansson, B.S.; Stensmyr, M.C. Evolution of Insect Olfaction. Neuron 2011, 72, 698–711. [Google Scholar] [CrossRef]
- Smith, D.P. Odor and Pheromone Detection in Drosophila melanogaster. Pflugers Arch. Eur. J. Physiol. 2007, 454, 749–758. [Google Scholar] [CrossRef]
- Gomez-Diaz, C.; Martin, F.; Garcia-Fernandez, J.M.; Alcorta, E. The Two Main Olfactory Receptor Families in Drosophila, ORs and IRs: A Comparative Approach. Front. Cell. Neurosci. 2018, 12, 253. [Google Scholar] [CrossRef] [PubMed]
- Benton, R. On the ORigin of Smell: Odorant Receptors in Insects. Cell. Mol. Life Sci. 2006, 63, 1579–1585. [Google Scholar] [CrossRef] [PubMed]
- Vosshall, L.B.; Wong, A.M.; Axel, R. An Olfactory Sensory Map in the Fly Brain. Cell 2000, 102, 147–159. [Google Scholar] [CrossRef]
- Pellegrino, M.; Nakagawa, T.; Vosshall, L.B. Single Sensillum Recordings in the Insects Drosophila melanogaster and Anopheles gambiae. J. Vis. Exp. 2010, 36, e1725. [Google Scholar] [CrossRef]
- Grabe, V.; Sachse, S. Fundamental Principles of the Olfactory Code. BioSystems 2018, 164, 94–101. [Google Scholar] [CrossRef]
- Wilson, R.I.; Mainen, Z.F. Early Events in Olfactory Processing. Annu. Rev. Neurosci. 2006, 29, 163–201. [Google Scholar] [CrossRef]
- Caron, S.J.C.; Ruta, V.; Abbott, L.F.; Axel, R. Random Convergence of Olfactory Inputs in the Drosophila Mushroom Body. Nature 2013, 497, 113–117. [Google Scholar] [CrossRef]
- Das Chakraborty, S.; Sachse, S. Olfactory Processing in the Lateral Horn of Drosophila. Cell Tissue Res. 2021, 383, 113–123. [Google Scholar] [CrossRef]
- Land, M.F. Visual Acuity in Insects. Annu. Rev. Entomol. 1997, 42, 147–177. [Google Scholar] [CrossRef]
- Bausenwein, B.; Dittrich, A.P.M.; Fischbach, K.F. The Optic Lobe of Drosophila melanogaster—II. Sorting of Retinotopic Pathways in the Medulla. Cell Tissue Res. 1992, 267, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Carthew, R.W. Pattern Formation in the Drosophila Eye. Curr. Opin. Genet. Dev. 2007, 17, 309–313. [Google Scholar] [CrossRef] [PubMed]
- Morante, J.; Desplan, C. The Color-Vision Circuit in the Medulla of Drosophila. Curr. Biol. 2008, 18, 553–565. [Google Scholar] [CrossRef]
- Schnaitmann, C.; Garbers, C.; Wachtler, T.; Tanimoto, H. Color Discrimination with Broadband Photoreceptors. Curr. Biol. 2013, 23, 2375–2382. [Google Scholar] [CrossRef] [PubMed]
- Schnaitmann, C.; Haikala, V.; Abraham, E.; Oberhauser, V.; Thestrup, T.; Griesbeck, O.; Reiff, D.F. Color Processing in the Early Visual System of Drosophila. Cell 2018, 172, 318–330.e18. [Google Scholar] [CrossRef]
- Salcedo, E.; Huber, A.; Henrich, S.; Chadwell, L.V.; Chou, W.H.; Paulsen, R.; Britt, S.G. Blue- and Green-Absorbing Visual Pigments of Drosophila: Ectopic Expression and Physiological Characterization of the R8 Photoreceptor Cell- Specific Rh5 and Rh6 Rhodopsins. J. Neurosci. 1999, 19, 10716–10726. [Google Scholar] [CrossRef] [PubMed]
- Hofbauer, A.; Campos-Ortega, J.A. Proliferation Pattern and Early Differentiation of the Optic Lobes in Drosophila melanogaster. Roux’s Arch. Dev. Biol. 1990, 198, 264–274. [Google Scholar] [CrossRef]
- Mu, L.; Ito, K.; Bacon, J.P.; Strausfeld, N.J. Optic Glomeruli and Their Inputs in Drosophila Share an Organizational Ground Pattern with the Antennal Lobes. J. Neurosci. 2012, 32, 6061–6071. [Google Scholar] [CrossRef]
- Schnaitmann, C.; Pagni, M.; Reiff, D.F. Color Vision in Insects: Insights from Drosophila. J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 2020, 206, 183–198. [Google Scholar] [CrossRef]
- Clark, D.A.; Bursztyn, L.; Horowitz, M.A.; Schnitzer, M.J.; Clandinin, T.R. Defining the Computational Structure of the Motion Detector in Drosophila. Neuron 2011, 70, 1165–1177. [Google Scholar] [CrossRef]
- Behnia, R.; Desplan, C. Visual Circuits in Flies: Beginning to See the Whole Picture The Eye and the Optic Lobe HHS Public Access. Curr. Opin. Neurobiol. 2015, 34, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Omoto, J.J.; Keleş, M.F.; Nguyen, B.C.M.; Bolanos, C.; Lovick, J.K.; Frye, M.A.; Hartenstein, V. Visual Input to the Drosophila Central Complex by Developmentally and Functionally Distinct Neuronal Populations. Curr. Biol. 2017, 27, 1098–1110. [Google Scholar] [CrossRef] [PubMed]
- Ling, F.; Dahanukar, A.; Weiss, L.A.; Kwon, J.Y.; Carlson, J.R. The Molecular and Cellular Basis of Taste Coding in the Legs of Drosophila. J. Neurosci. 2014, 34, 7148–7164. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.C.D.; Dahanukar, A. Recent Advances in the Genetic Basis of Taste Detection in Drosophila. Cell. Mol. Life Sci. 2020, 77, 1087–1101. [Google Scholar] [CrossRef]
- Tuthill, J.C.; Wilson, R.I. Mechanosensation and Adaptive Motor Control in Insects. Curr. Biol. 2016, 26, R1022–R1038. [Google Scholar] [CrossRef]
- Patella, P.; Wilson, R.I. Functional Maps of Mechanosensory Features in the Drosophila Brain. Curr. Biol. 2018, 28, 1189–1203.e5. [Google Scholar] [CrossRef]
- Suver, M.P.; Matheson, A.M.M.; Sarkar, S.; Damiata, M.; Schoppik, D.; Nagel, K.I. Encoding of Wind Direction by Central Neurons in Drosophila. Neuron 2019, 102, 828–842.e7. [Google Scholar] [CrossRef]
- Dujardin, F. Mémoire Sur Le Système Nerveux Des Insectes. Ann. Sci. Nat. Zool. 1850, 14, 195–206. [Google Scholar]
- Strausfeld, N.J.; Hansen, L.; Li, Y.; Gomez, R.S.; Ito, K. Evolution, Discovery, and Interpretations of Arthropod Mushroom Bodies. Learn. Mem. 1998, 5, 11–37. [Google Scholar] [CrossRef]
- Rybak, J.; Menzel, R. Anatomy of the Mushroom Bodies in the Honey Bee Brain: The Neuronal Connections of the Alpha-lobe. J. Comp. Neurol. 1993, 334, 444–465. [Google Scholar] [CrossRef]
- Vogt, K.; Aso, Y.; Hige, T.; Knapek, S.; Ichinose, T.; Friedrich, A.B.; Turner, G.C.; Rubin, G.M.; Tanimoto, H. Direct Neural Pathways Convey Distinct Visual Information to Drosophila Mushroom Bodies. Elife 2016, 5, e14009. [Google Scholar] [CrossRef] [PubMed]
- Morante, J.; Desplan, C. The Color Vision Circuit in Drosophila. J. Neurogenet. 2006, 20, 181–182. [Google Scholar]
- Yagi, R.; Mabuchi, Y.; Mizunami, M.; Tanaka, N.K. Convergence of Multimodal Sensory Pathways to the Mushroom Body Calyx in Drosophila melanogaster. Sci. Rep. 2016, 6, 29481. [Google Scholar] [CrossRef] [PubMed]
- Gronenberg, W. Subdivisions of Hymenopteran Mushroom Body Calyces by Their Afferent Supply. J. Comp. Neurol. 2001, 435, 474–489. [Google Scholar] [CrossRef]
- Schröter, U.; Menzel, R. A New Ascending Sensory Tract to the Calyces of the Honeybee Mushroom Body, the Subesophageal-Calycal Tract. J. Comp. Neurol. 2003, 465, 168–178. [Google Scholar] [CrossRef]
- Li, J.; Mahoney, B.D.; Jacob, M.S.; Caron, S.J.C. Visual Input into the Drosophila melanogaster Mushroom Body. Cell Rep. 2020, 32, 108138. [Google Scholar] [CrossRef]
- Mobbs, P.G.; Young, J.Z. The Brain of the Honeybee Apis mellifera. I. The Connections and Spatial Organization of the Mushroom Bodies. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1982, 298, 309–354. [Google Scholar] [CrossRef]
- Mizunami, M.; Iwasaki, M.; Nishikawa, M.; Okada, R. Modular Structures in the Mushroom Body of the Cockroach. Neurosci. Lett. 1997, 229, 153–156. [Google Scholar] [CrossRef]
- Okada, R.; Ikeda, J.; Mizunami, M. Sensory Responses and Movement-Related Activities in Extrinsic Neurons of the Cockroach Mushroom Bodies. J. Comp. Physiol. A Sens. Neural Behav. Physiol. 1999, 185, 115–129. [Google Scholar] [CrossRef]
- Strube-Bloss, M.F.; Rössler, W. Multimodal Integration and Stimulus Categorization in Putative Mushroom Body Output Neurons of the Honeybee. R. Soc. Open Sci. 2018, 5, 171785. [Google Scholar] [CrossRef]
- Liu, L.; Wolf, R.; Ernst, R.; Heisenberg, M. Context Generalization in Drosophila Visual Learning Requires the Mushroom Bodies. Nature 1999, 400, 753–756. [Google Scholar] [CrossRef]
- Vogt, K.; Schnaitmann, C.; Dylla, K.V.; Knapek, S.; Aso, Y.; Rubin, G.M.; Tanimoto, H. Shared Mushroom Body Circuits Underlie Visual and Olfactory Memories in Drosophila. Elife 2014, 3, e02395. [Google Scholar] [CrossRef] [PubMed]
- Aso, Y.; Sitaraman, D.; Ichinose, T.; Kaun, K.R.; Vogt, K.; Belliart-Guérin, G.; Plaçais, P.Y.; Robie, A.A.; Yamagata, N.; Schnaitmann, C.; et al. Mushroom Body Output Neurons Encode Valence and Guide Memory-Based Action Selection in Drosophila. Elife 2014, 3, e04580. [Google Scholar] [CrossRef] [PubMed]
- Kirkhart, C.; Scott, K. Gustatory Learning and Processing in the Drosophila Mushroom Bodies. J. Neurosci. 2015, 35, 5950–5958. [Google Scholar] [CrossRef] [PubMed]
- Barth, M.; Heisenberg, M. Vision Affects Mushroom Bodies and Central Complex in Drosophila melanogaster. Learn. Mem. 1997, 4, 219–229. [Google Scholar] [CrossRef]
- Chow, D.M.; Theobald, J.C.; Frye, M.A. An Olfactory Circuit Increases the Fidelity of Visual Behavior. J. Neurosci. 2011, 31, 15035–15047. [Google Scholar] [CrossRef]
- Li, J.; Mahoney, B.D.; Jacob, M.S.; Caron, S.J.C. Two Parallel Pathways Convey Distinct Visual Information to the Drosophila Mushroom Body. bioRxiv 2020. [Google Scholar] [CrossRef]
- Power, M.E. The Brain of Drosophila melanogaster. J. Morphol. 1943, 72, 517–559. [Google Scholar] [CrossRef]
- Homberg, U. Evolution of the Central Complex in the Arthropod Brain with Respect to the Visual System. Arthropod Struct. Dev. 2008, 37, 347–362. [Google Scholar] [CrossRef]
- Strauss, R. The Central Complex and the Genetic Dissection of Locomotor Behaviour. Curr. Opin. Neurobiol. 2002, 12, 633–638. [Google Scholar] [CrossRef]
- Martin, J.P.; Guo, P.; Mu, L.; Harley, C.M.; Ritzmann, R.E. Central-Complex Control of Movement in the Freely Walking Cockroach. Curr. Biol. 2015, 25, 2795–2803. [Google Scholar] [CrossRef]
- Homberg, U. In Search of the Sky Compass in the Insect Brain. Naturwissenschaften 2004, 91, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Strausfeld, N.J. Chapter 24 A Brain Region in Insects That Supervises Walking. In Peripheral and Spinal Mechanisms in the Neural Control of Movement; Binder, M.D., Ed.; Elsevier: Amsterdam, The Netherlands, 1999; Volume 123, pp. 273–284. [Google Scholar] [CrossRef]
- Currier, T.A.; Matheson, A.M.M.; Nagel, K.I. Encoding and Control of Orientation to Airflow by a Set of Drosophila Fan-Shaped Body Neurons. Elife 2020, 9, e61510. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Peng, Y.; Sun, J.; Zhang, F.; Zhang, X.; Wang, L.; Li, Q.; Zhong, Y. Fan-Shaped Body Neurons in the Drosophila Brain Regulate Both Innate and Conditioned Nociceptive Avoidance. Cell Rep. 2018, 24, 1573–1584. [Google Scholar] [CrossRef]
- Hardcastle, B.J.; Omoto, J.J.; Kandimalla, P.; Nguyen, B.C.M.; Keleş, M.F.; Boyd, N.K.; Hartenstein, V.; Frye, M.A. A Visual Pathway for Skylight Polarization Processing in Drosophila. Elife 2021, 10, e63225. [Google Scholar] [CrossRef]
- Kim, S.S.; Rouault, H.; Druckmann, S.; Jayaraman, V. Ring Attractor Dynamics in the Drosophila Central Brain. Science 2017, 356, 849–853. [Google Scholar] [CrossRef]
- Seelig, J.D.; Jayaraman, V. Neural Dynamics for Landmark Orientation and Angular Path Integration. Nature 2015, 521, 186–191. [Google Scholar] [CrossRef]
- Pfeiffer, K.; Homberg, U. Organization and Functional Roles of the Central Complex in the Insect Brain. Annu. Rev. Entomol. 2014, 59, 165–184. [Google Scholar] [CrossRef]
- Pickard, S.C.; Quinn, R.D.; Szczecinski, N.S. A Dynamical Model Exploring Sensory Integration in the Insect Central Complex Substructures. Bioinspir. Biomim. 2020, 15, 026003. [Google Scholar] [CrossRef]
- Brembs, B. Mushroom Bodies Regulate Habit Formation in Drosophila. Curr. Biol. 2009, 19, 1351–1355. [Google Scholar] [CrossRef]
- Ofstad, T.A.; Zuker, C.S.; Reiser, M.B. Visual Place Learning in Drosophila melanogaster. Nature 2011, 474, 204–209. [Google Scholar] [CrossRef] [PubMed]
- Heisenberg, M.; Borst, A.; Wagner, S.; Byers, D. Drosophila Mushroom Body Mutants Are Deficient in Olfactory Learning: Research Papers. J. Neurogenet. 1985, 2, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Bouhouche, A.; Vaysse, G.; Corbiegre, M. Immunocytochemical and Learning Studies of a Drosophila melanogaster Neurological Mutant, No-BridgeKS49 as an Approach to the Possible Role of the Central Complex. J. Neurogenet. 1993, 9, 105–121. [Google Scholar] [CrossRef] [PubMed]
- Collett, M.; Collett, T.S. How Does the Insect Central Complex Use Mushroom Body Output for Steering? Curr. Biol. 2018, 28, R733–R734. [Google Scholar] [CrossRef]
- Hulse, B.K.; Haberkern, H.; Franconville, R.; Turner-evans, D.B.; Parekh, R.; Hermundstad, A.M.; Rubin, G.M.; Jayaraman, V. A Connectome of the Drosophila Central Complex Reveals Network Motifs Suitable for Flexible Navigation and Context-Dependent Action Selection. Elife 2021, 10, e66039. [Google Scholar] [CrossRef]
- Jefferis, G.S.X.E.; Potter, C.J.; Chan, A.M.; Marin, E.C.; Rohlfing, T.; Maurer, C.R.; Luo, L. Comprehensive Maps of Drosophila Higher Olfactory Centers: Spatially Segregated Fruit and Pheromone Representation. Cell 2007, 128, 1187–1203. [Google Scholar] [CrossRef]
- Schultzhaus, J.N.; Saleem, S.; Iftikhar, H.; Carney, G.E. The Role of the Drosophila Lateral Horn in Olfactory Information Processing and Behavioral Response. J. Insect Physiol. 2017, 98, 29–37. [Google Scholar] [CrossRef]
- Gupta, N.; Stopfer, M. Functional Analysis of a Higher Olfactory Center, the Lateral Horn. J. Neurosci. 2012, 32, 8138–8148. [Google Scholar] [CrossRef]
- Strutz, A.; Soelter, J.; Baschwitz, A.; Farhan, A.; Grabe, V.; Rybak, J.; Knaden, M.; Schmuker, M.; Hansson, B.S.; Sachse, S. Decoding Odor Quality and Intensity in the Drosophila Brain. Elife 2014, 3, e04147. [Google Scholar] [CrossRef]
- Frechter, S.; Bates, A.S.; Tootoonian, S.; Dolan, M.J.; Manton, J.; Jamasb, A.R.; Kohl, J.; Bock, D.; Jefferis, G. Functional and Anatomical Specificity in a Higher Olfactory Centre. Elife 2019, 8, e44590. [Google Scholar] [CrossRef]
- Ramdya, P.; Lichocki, P.; Cruchet, S.; Frisch, L.; Tse, W.; Floreano, D.; Benton, R. Mechanosensory Interactions Drive Collective Behaviour in Drosophila. Nature 2015, 519, 233–236. [Google Scholar] [CrossRef] [PubMed]
- Thistle, R.; Cameron, P.; Ghorayshi, A.; Dennison, L.; Scott, K. Contact Chemoreceptors Mediate Male-Male Repulsion and Male-Female Attraction during Drosophila Courtship. Cell 2012, 149, 1140–1151. [Google Scholar] [CrossRef] [PubMed]
- Clemens, J.; Girardin, C.C.; Coen, P.; Guan, X.J.; Dickson, B.J.; Murthy, M. Connecting Neural Codes with Behavior in the Auditory System of Drosophila. Neuron 2015, 87, 1332–1343. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, E.; Seki, H.; Asai, T.; Morimoto, T.; Miyakawa, H.; Ito, K.; Kamikouchi, A. Organization of Projection Neurons and Local Neurons of the Primary Auditory Center in the Fruit Fly Drosophila melanogaster. J. Comp. Neurol. 2016, 524, 1099–1164. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, A.A.M.; Hansson, B.S.; Sachse, S. Third-Order Neurons in the Lateral Horn Enhance Bilateral Contrast of Odor Inputs Through Contralateral Inhibition in Drosophila. Front. Physiol. 2019, 10, 851. [Google Scholar] [CrossRef]
- Zhou, C.; Franconville, R.; Vaughan, A.G.; Robinett, C.C.; Jayaraman, V.; Baker, B.S. Central Neural Circuitry Mediating Courtship Song Perception in Male Drosophila. Elife 2015, 4, e8477. [Google Scholar] [CrossRef]
- Zhu, Y. The Drosophila Visual System: From Neural Circuits to Behavior. Cell Adhes. Migr. 2013, 7, 333–344. [Google Scholar] [CrossRef]
- Lai, J.S.Y.; Lo, S.J.; Dickson, B.J.; Chiang, A.S. Auditory Circuit in the Drosophila Brain. Proc. Natl. Acad. Sci. USA 2012, 109, 2607–2612. [Google Scholar] [CrossRef]
- Eschbach, C.; Fushiki, A.; Winding, M.; Afonso, B.; Andrade, I.V.; Cocanougher, B.T.; Eichler, K.; Gepner, R.; Si, G.; Valdes-Aleman, J.; et al. Circuits for Integrating Learned and Innate Valences in the Insect Brain. Elife 2021, 10, e62567. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thiagarajan, D.; Sachse, S. Multimodal Information Processing and Associative Learning in the Insect Brain. Insects 2022, 13, 332. https://doi.org/10.3390/insects13040332
Thiagarajan D, Sachse S. Multimodal Information Processing and Associative Learning in the Insect Brain. Insects. 2022; 13(4):332. https://doi.org/10.3390/insects13040332
Chicago/Turabian StyleThiagarajan, Devasena, and Silke Sachse. 2022. "Multimodal Information Processing and Associative Learning in the Insect Brain" Insects 13, no. 4: 332. https://doi.org/10.3390/insects13040332
APA StyleThiagarajan, D., & Sachse, S. (2022). Multimodal Information Processing and Associative Learning in the Insect Brain. Insects, 13(4), 332. https://doi.org/10.3390/insects13040332