
Different Distribution Patterns of Hoverflies (Diptera: Syrphidae) and Bees (Hymenoptera: Anthophila) Along Altitudinal Gradients in Dolomiti Bellunesi National Park (Italy)




Abstract
:Simple Summary
Abstract
1. Introduction
- To evaluate the altitudinal distribution of two important pollinator taxa, i.e., Hymenoptera Anthophila (bees) and Diptera Syrphidae (hoverflies), also including the phenological patterns;
- To compare the effects of landscape and altitude as main factors affecting species distribution.
2. Materials and Methods
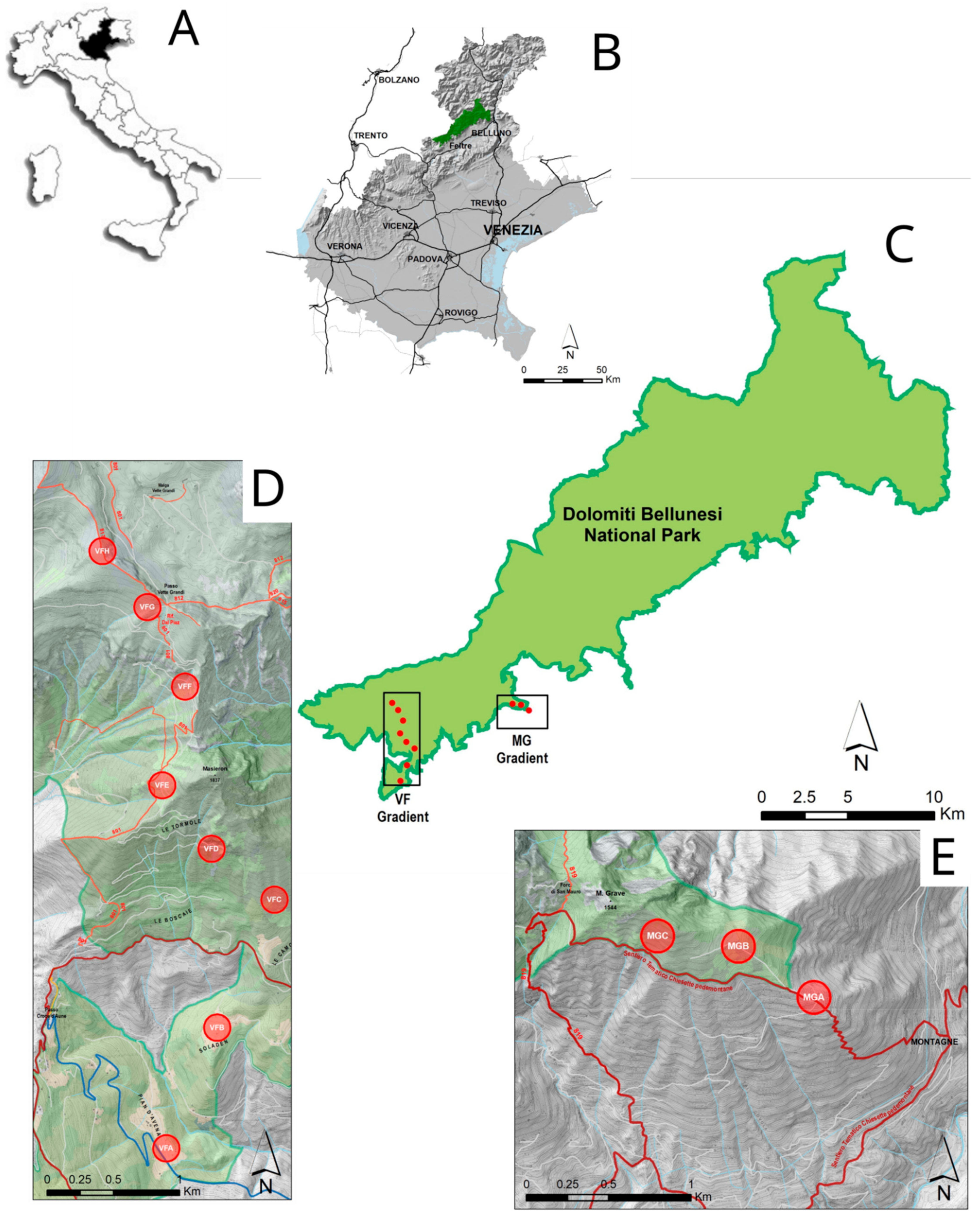
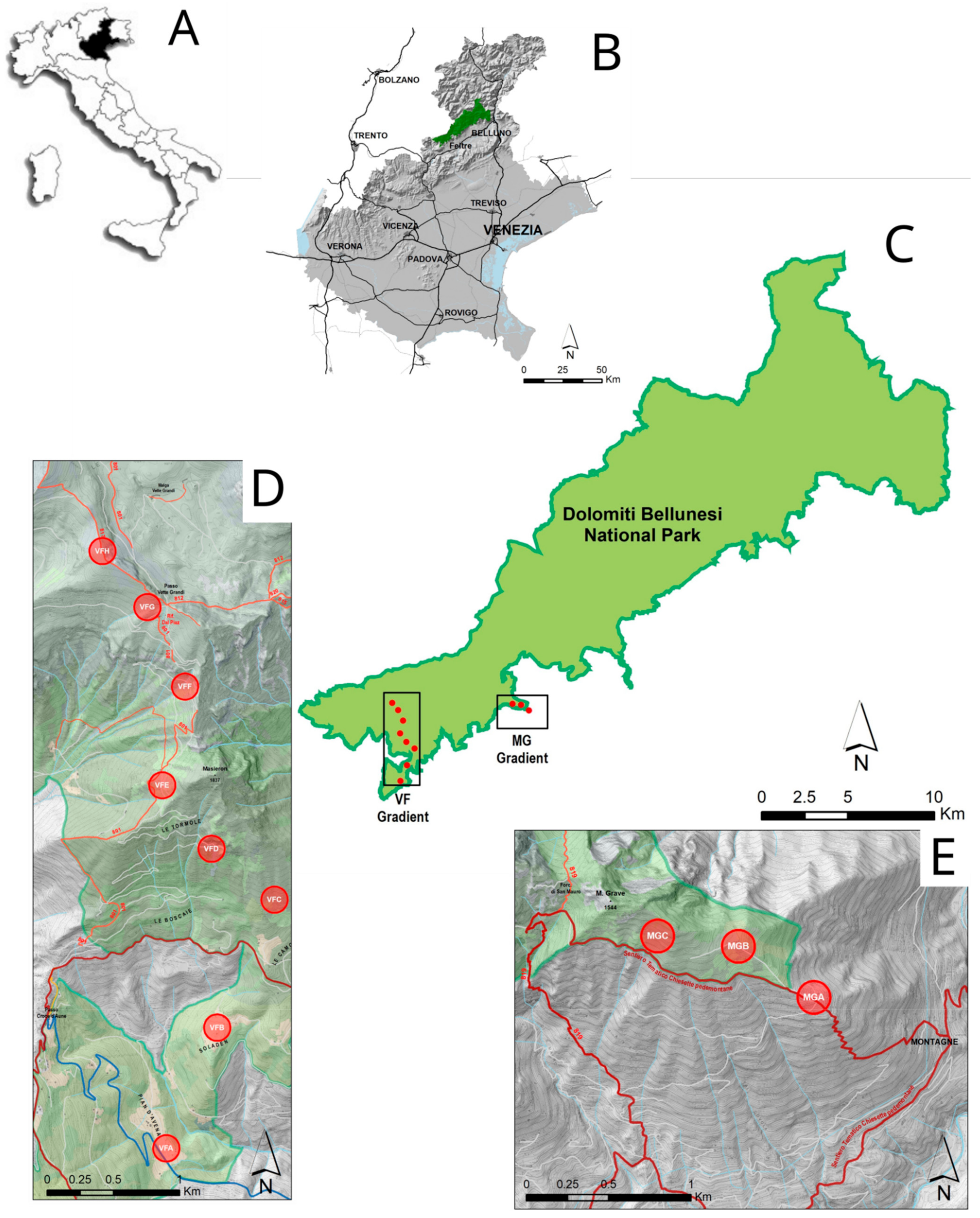
2.1. Study Site
- Vette Feltrine (VF): This gradient starts at 780 until 2130 m a.s.l.; 8 circular plots (100 m radius) were selected at around 200 m altitude difference (Table 1).
- Mount Grave (MG): This gradient is shorter than previous one, starting at 1080 m and ending at 1375 m; 3 circular plots were selected at altitude differences of around 200 and 100 m (Table 1).
2.2. Sampling Design
- -
- -
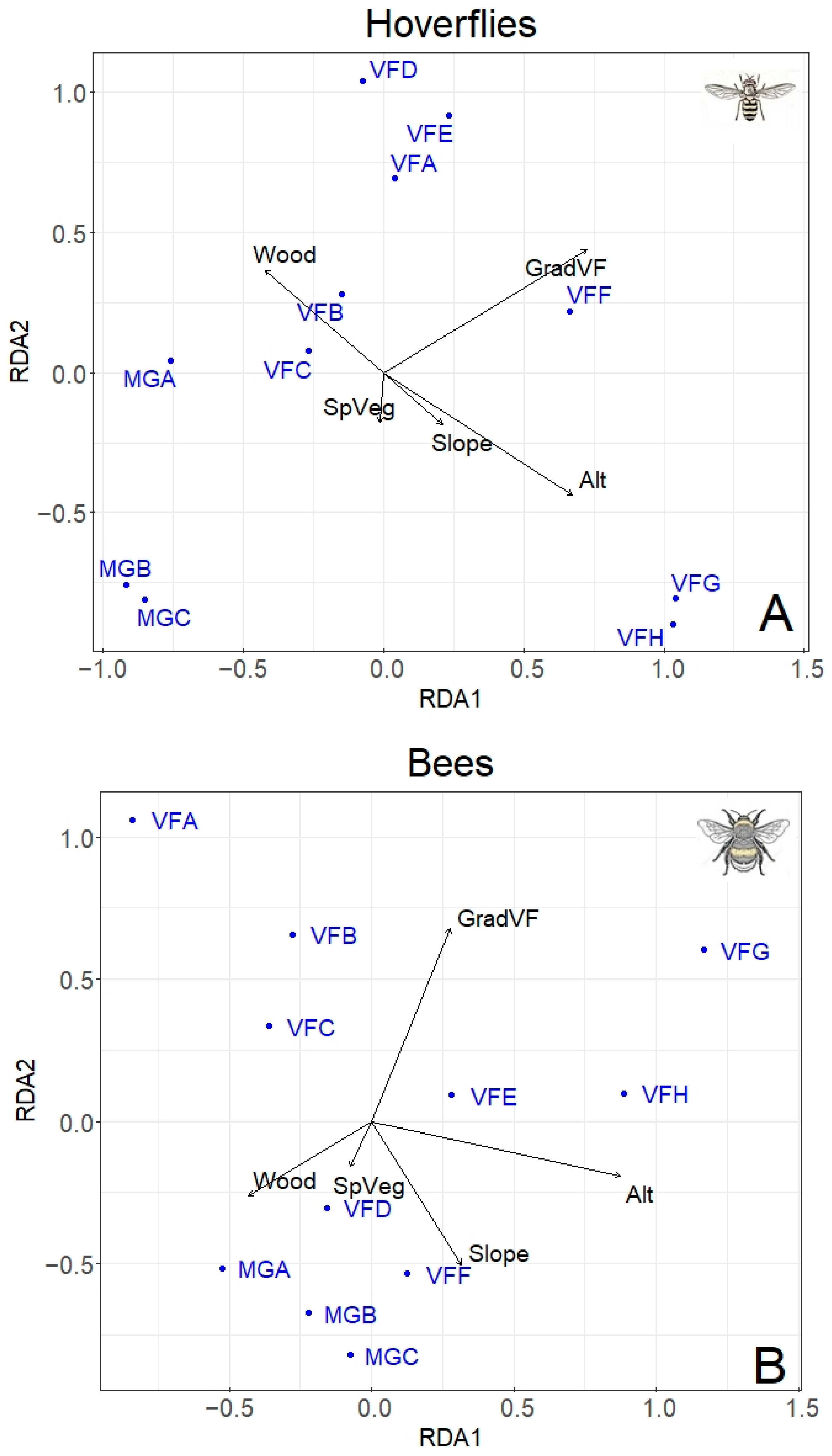
2.3. Statistical Analysis
- Temperature was correlated with altitude;
- Percentage of grassland was inversely correlated with percentage of woods;
- Percentage of scree was correlated with altitude;
- Percentage of scrubs was correlated with percentage of grassland;
- Slope was correlated with plant species pollinated by insects;
- Total plant species was correlated with plant species pollinated by insects.
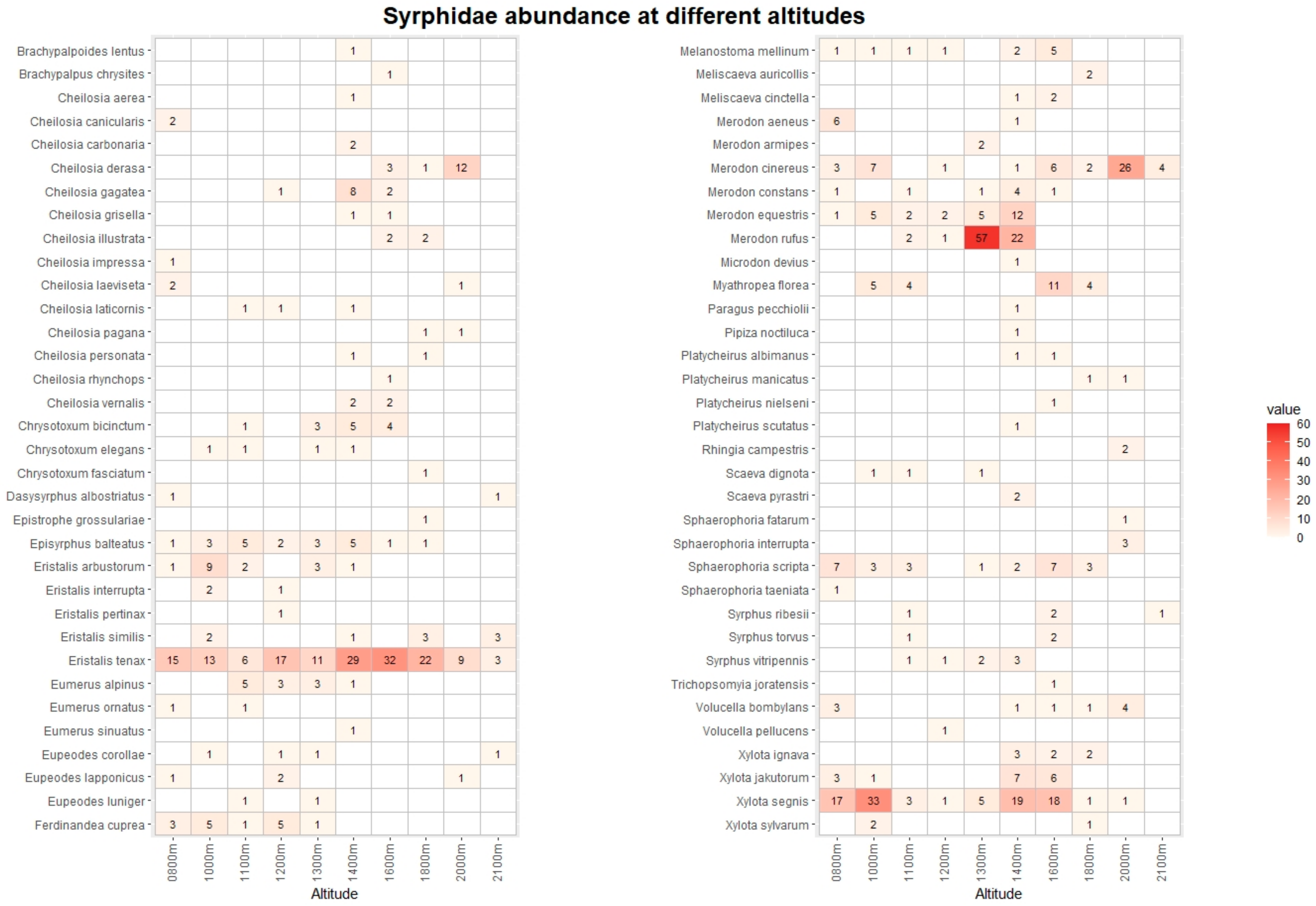
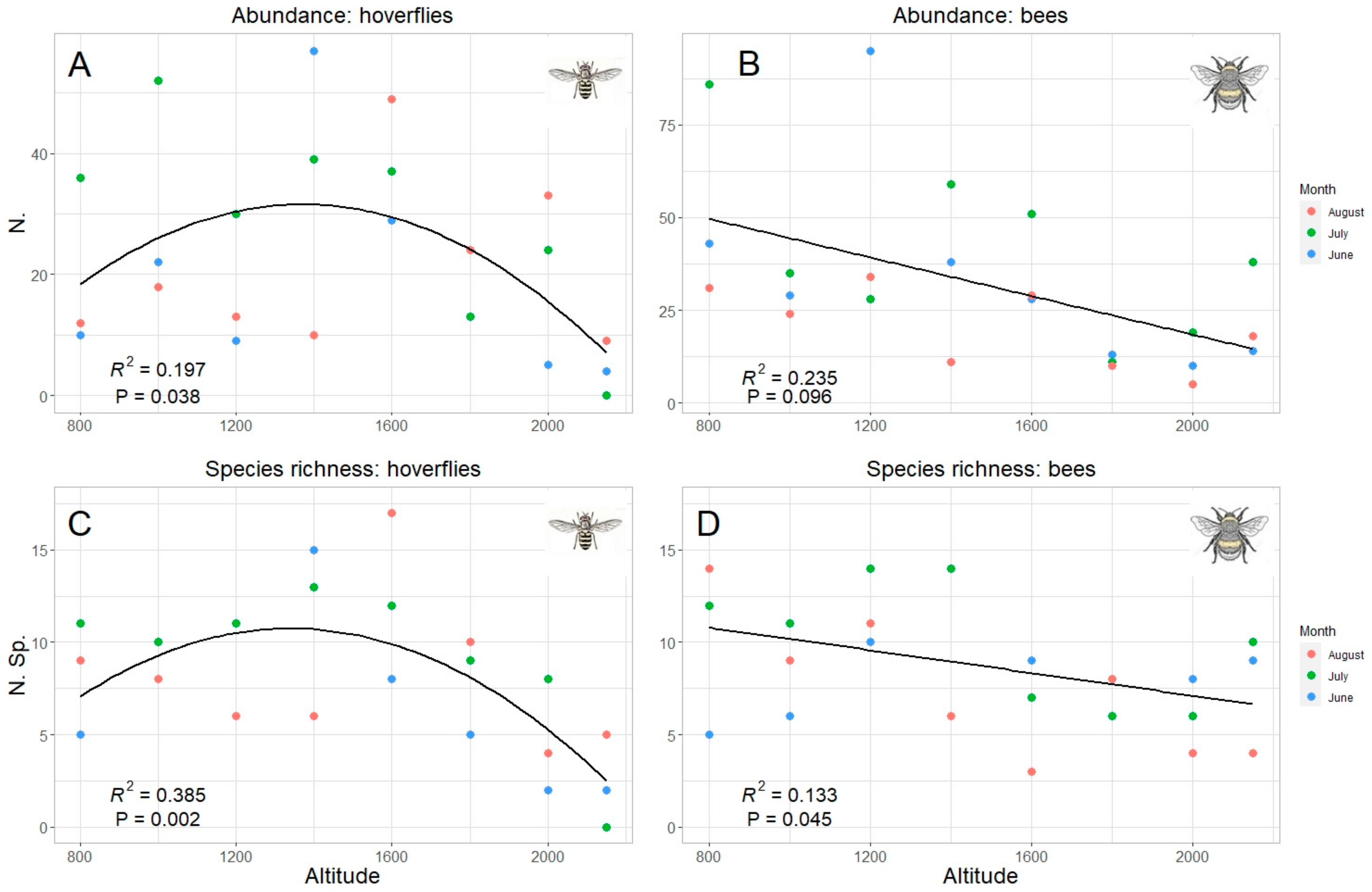
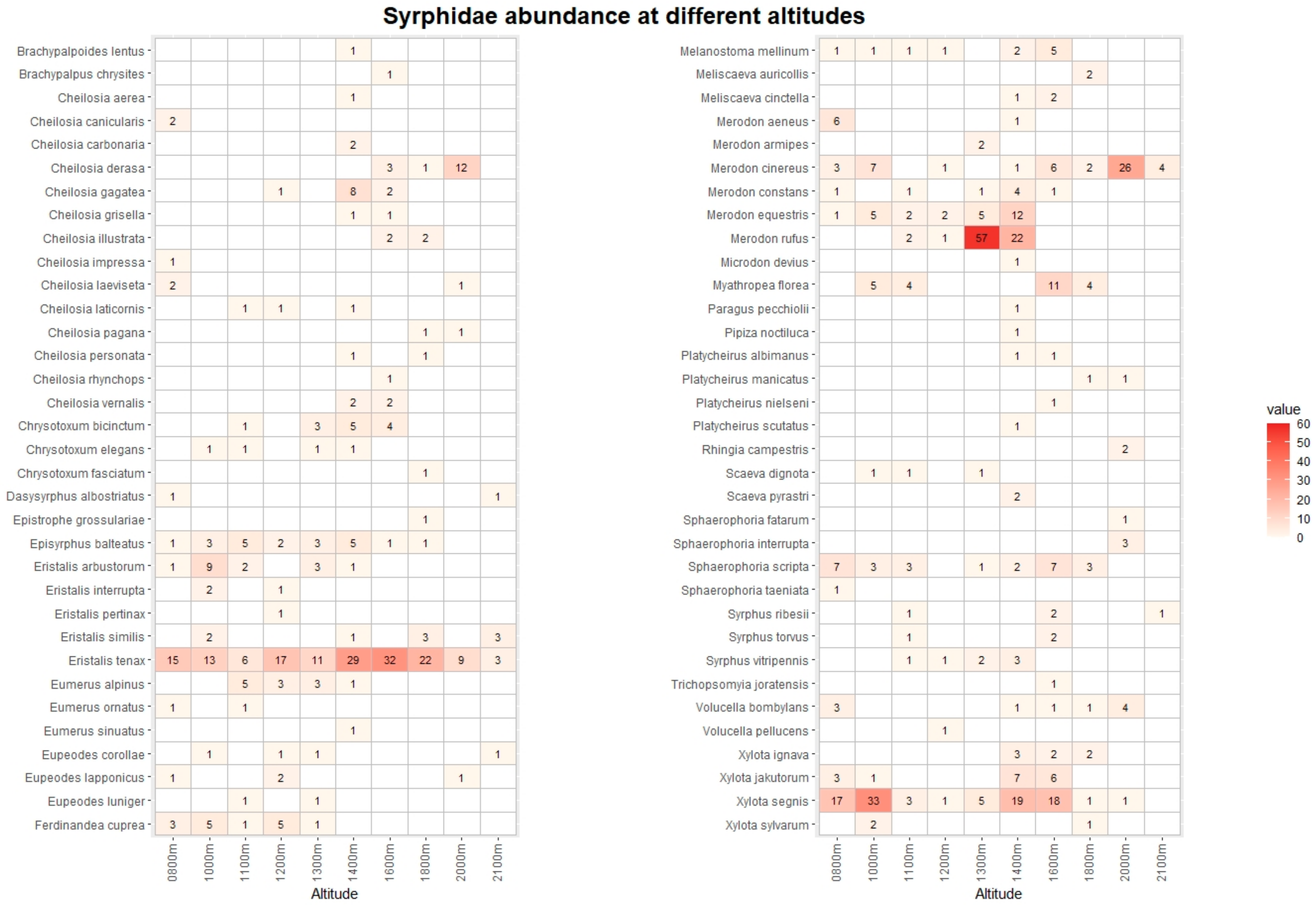
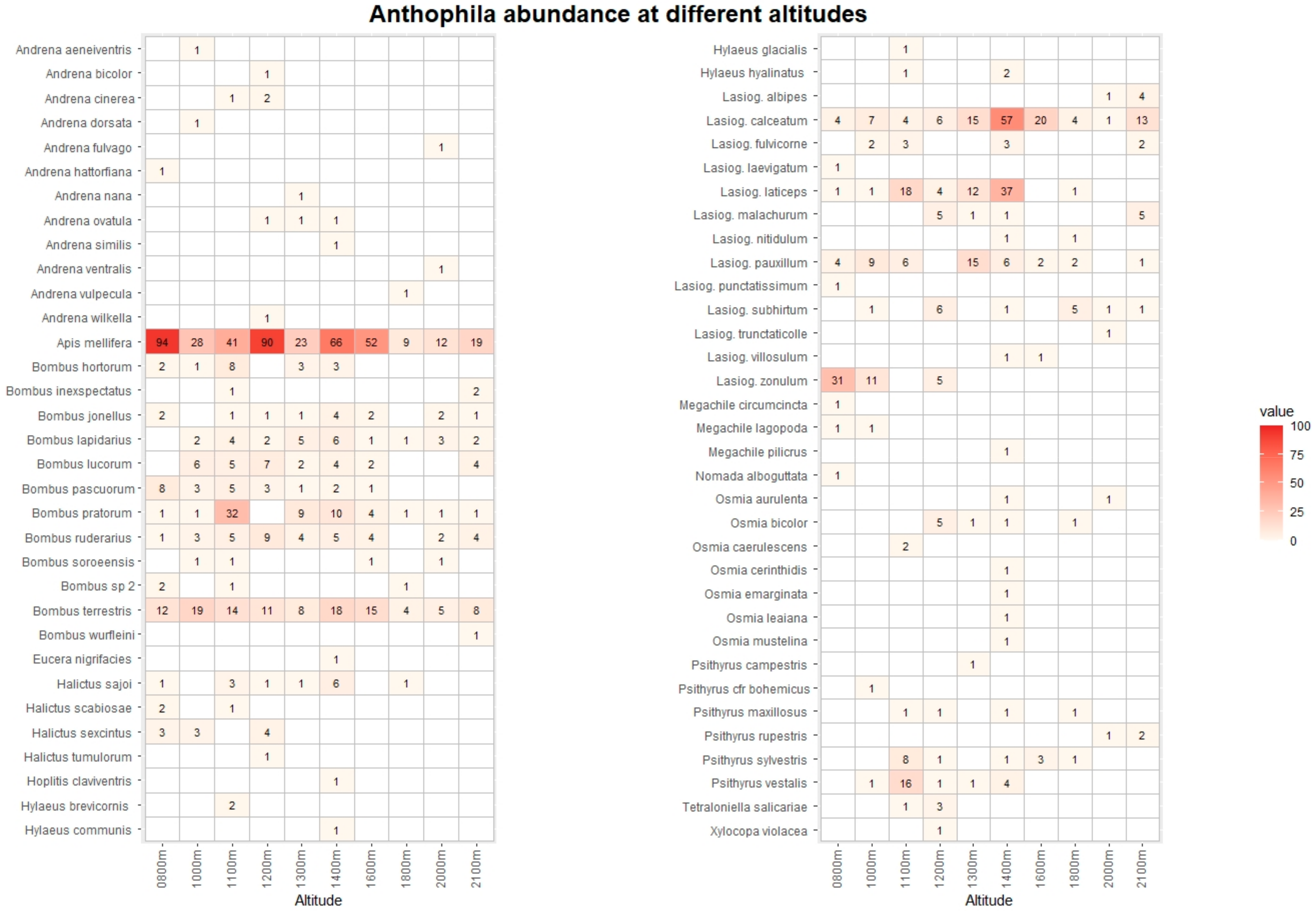
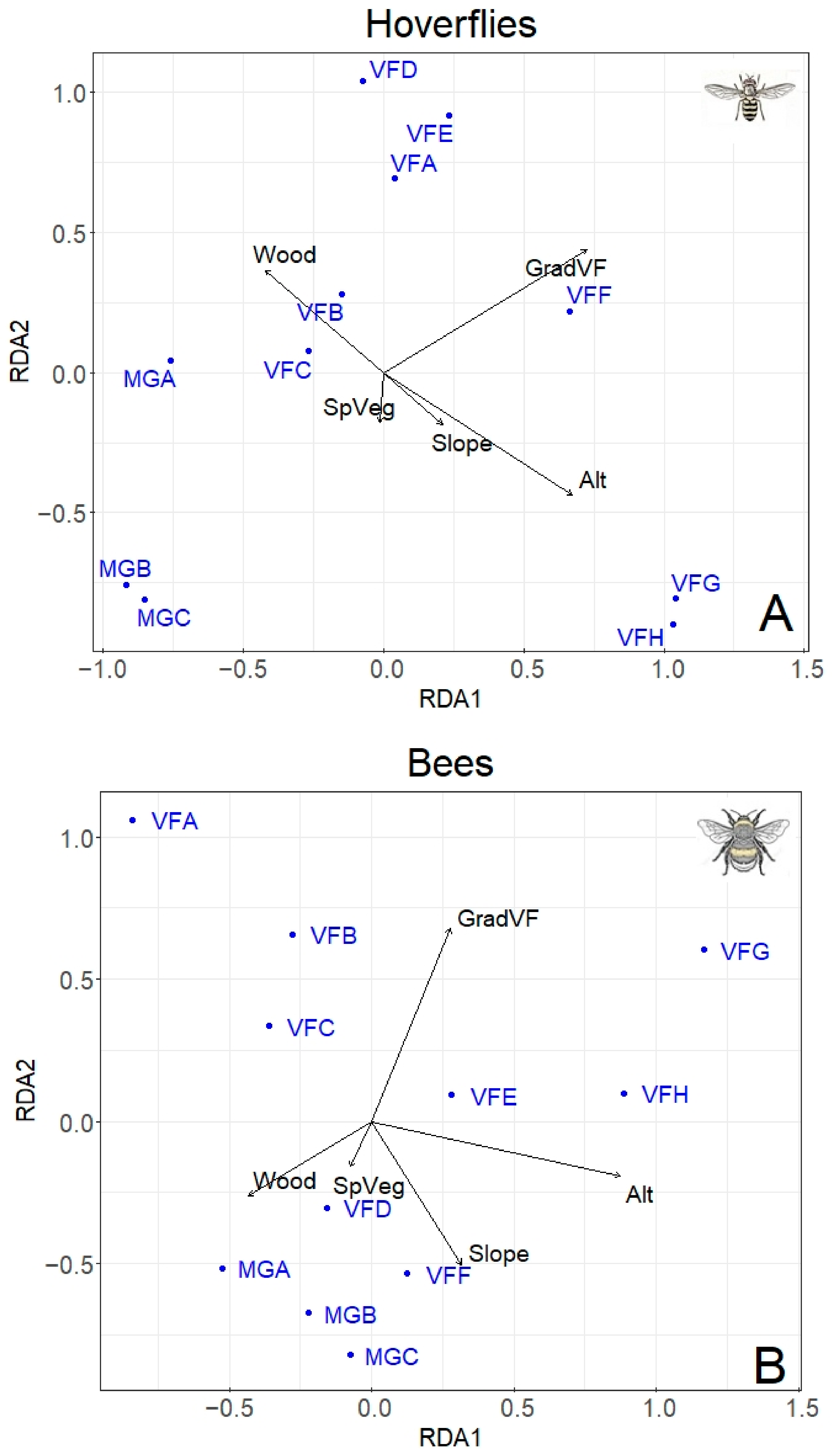
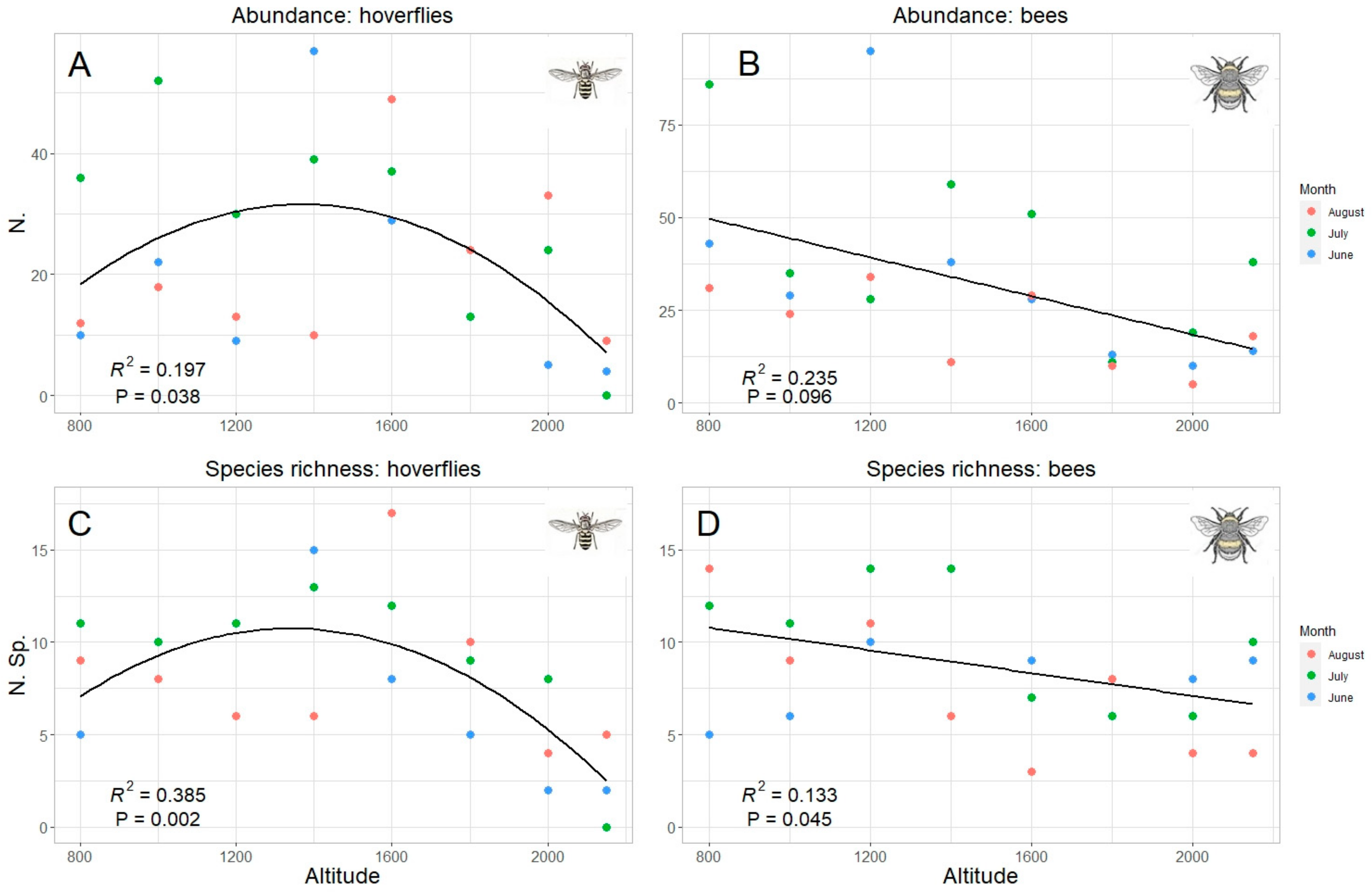
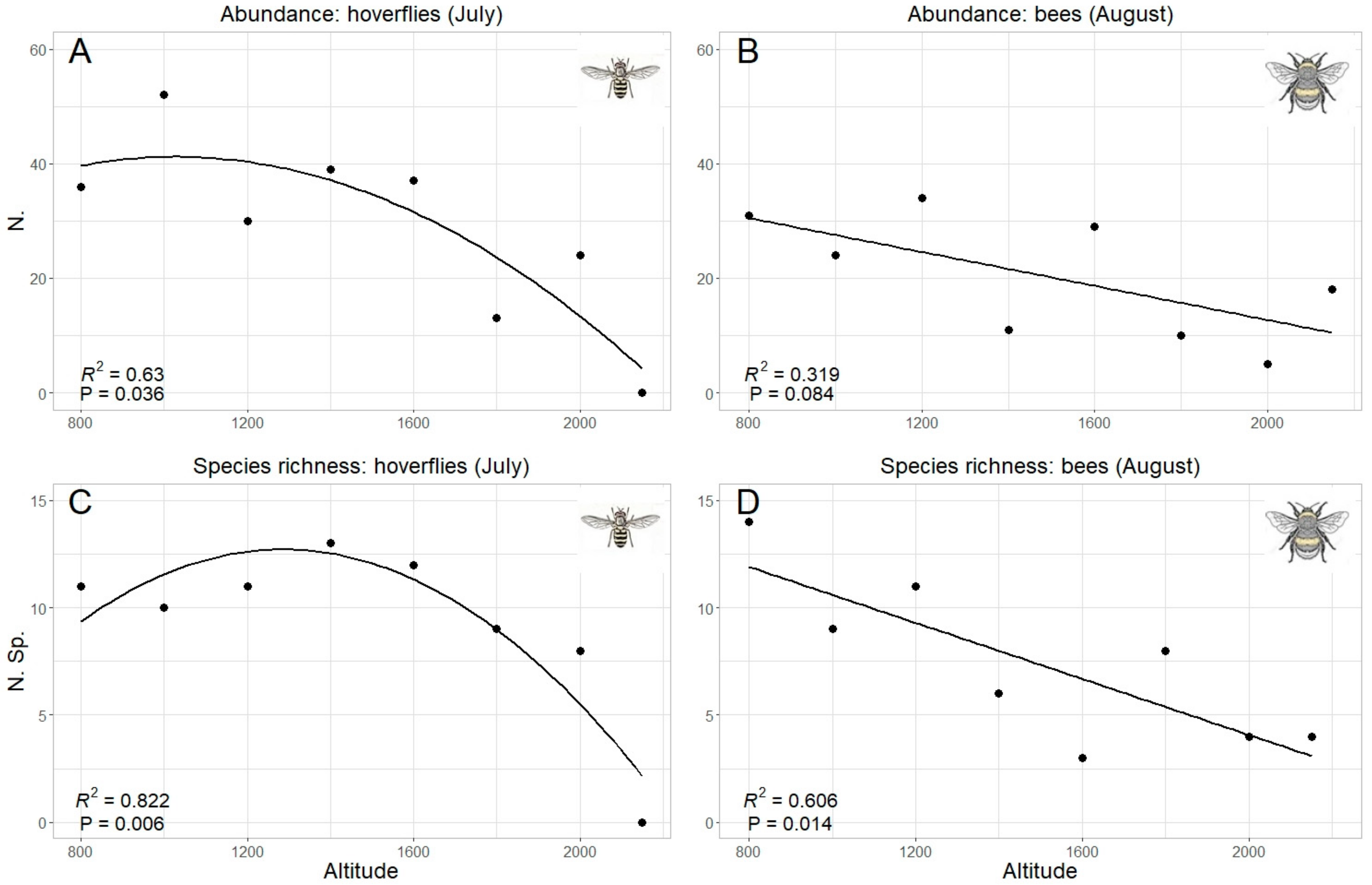
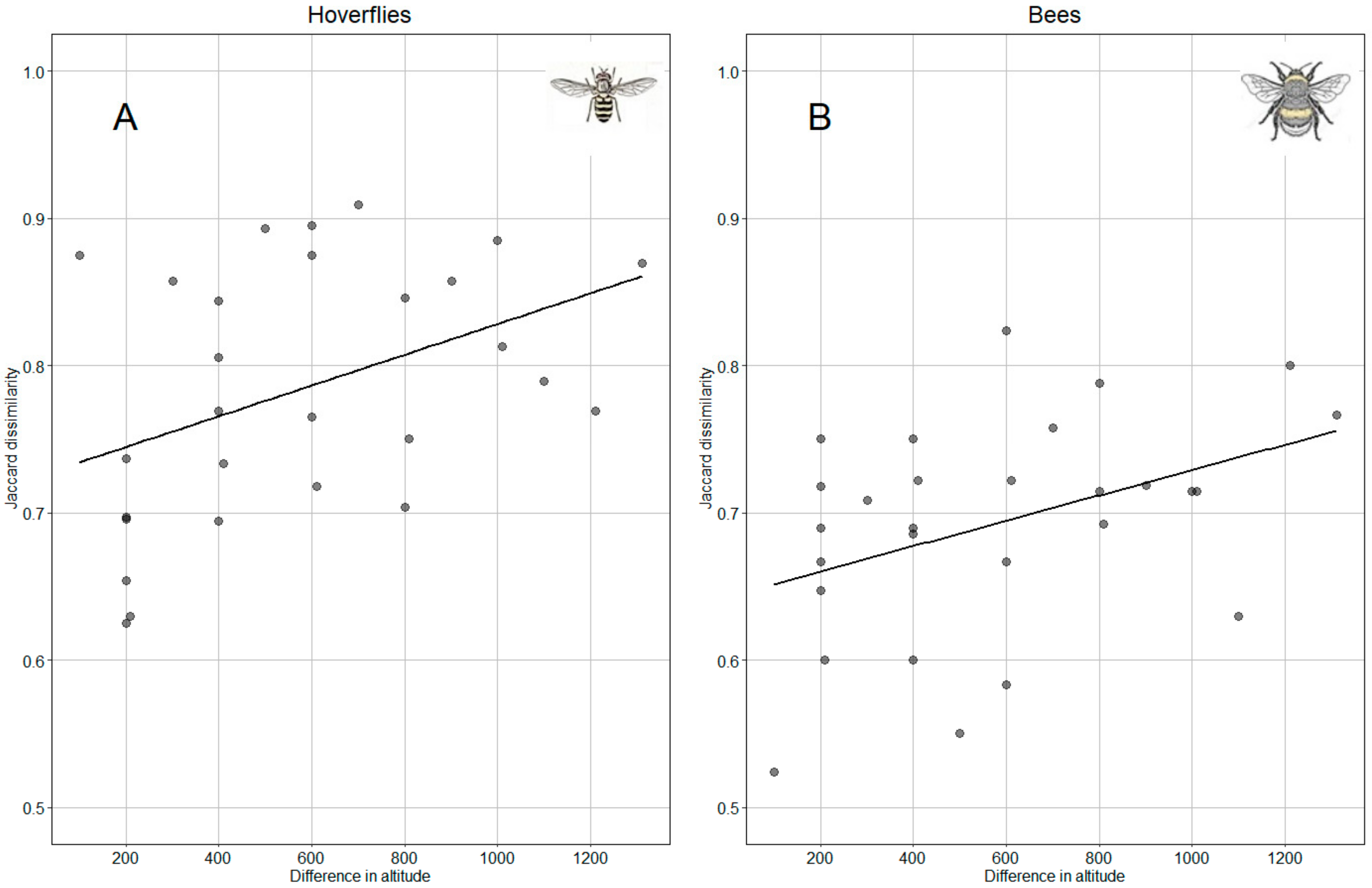
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ghazoul, J. Qualifying pollinator decline evidence. Science 2015, 348, 981–982. [Google Scholar] [CrossRef]
- Shivanna, K.R.; Tandon, R.; Koul, M. ‘Global Pollinator Crisis’ and its impact on crop productivity and sustenance of plant diversity. In Reproductive Ecology of Flowering Plants: Patterns and Processes; Tandon, R., Shivanna, K.R., Koul, M., Eds.; Springer: Singapore, 2020; pp. 395–413. [Google Scholar]
- Ollerton, J. Pollinators & Pollination: Nature and Society; Pelagic Publishing: Exeter, UK, 2021; 286p. [Google Scholar]
- Kosior, A.; Celary, W.; Olejniczak, P.; Fijał, J.; Król, W.; Solarz, W.; Płonka, P. The decline of the bumble bees and cuckoo bees (Hymenoptera: Apidae: Bombini) of Western and Central Europe. Oryx 2007, 41, 79–88. [Google Scholar] [CrossRef] [Green Version]
- Grixti, J.C.; Wong, L.T.; Cameron, S.A.; Favret, C. Decline of bumble bees (Bombus) in the North American Midwest. Biol. Conserv. 2009, 142, 75–84. [Google Scholar] [CrossRef]
- Williams, P.H.; Osborne, J.L. Bumblebee vulnerability and conservation world-wide. Apidologie 2009, 40, 367–387. [Google Scholar] [CrossRef] [Green Version]
- Cameron, S.A.; Lozier, J.D.; Strange, J.P.; Koch, J.B.; Cordes, N.; Solter, L.F.; Griswold, T.L. Patterns of widespread decline in North American bumble bees. Proc. Natl. Acad. Sci. USA 2011, 108, 662–667. [Google Scholar] [CrossRef] [Green Version]
- Bartomeus, I.; Ascher, J.S.; Gibbs, J.; Danforth, B.N.; Wagner, D.L.; Hedtke, S.M.; Winfree, R. Historical changes in northeastern US bee pollinators related to shared ecological traits. Proc. Natl. Acad. Sci. USA 2013, 110, 4656–4660. [Google Scholar] [CrossRef] [Green Version]
- Nieto, A.; Roberts, S.P.M.; Kemp, J.; Rasmont, P.; Kuhlmann, M.; Criado, M.G.; Biesmeijer, J.C.; Bogusch, P.; Dathe, H.H.; De la Rúa, P.; et al. European Red List of Bees; Publication Office of the European Union: Luxembourg, 2014; 84p. [Google Scholar]
- Van Swaay, C.A.M.; Cuttelod, A.; Collins, S.; Maes, D.; Munguira, M.L.; Settele, J.; Verovnik, R.; Verstrael, T.; Warren, M.S.; Wiemers, M.; et al. European Red List of Butterflies; Publications Office of the European Union: Luxembourg, 2010; 47p. [Google Scholar]
- Biesmeijer, J.C.; Roberts, S.P.M.; Reemer, M.; Ohlemüller, R.; Edwards, M.; Peeters, T.; Schaffers, A.P.; Potts, S.G.; Kleukers, R.; Thomas, C.D.; et al. Parallel Declines in Pollinators and Insect-Pollinated Plants in Britain and the Netherlands. Science 2006, 313, 351–354. [Google Scholar] [CrossRef]
- Speight, M.C.D. Species accounts of European Syrphidae, 2020. In Syrph the Net, the Database of European Syrphidae (Diptera); Syrph the Net Publications: Dublin, Ireland, 2020; Volume 104, 314p. [Google Scholar]
- IPBES. The assessment report of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services on pollinators, pollination and food production. In Secretariat of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services; Potts, S.G., Imperatriz-Fonseca, V.L., Ngo, H.T., Eds.; NERC Open Research Archive: Bonn, Germany, 2016. [Google Scholar]
- Godfray, H.C.J.; Blacquière, T.; Field, L.; Hails, R.S.; Potts, S.G.; Raine, N.E.; Vanbergen, A.; McLean, A.R. A restatement of recent advances in the natural science evidence base concerning neonicotinoid insecticides and insect pollinators. Proc. R. Soc. B Boil. Sci. 2015, 282, 20151821. [Google Scholar] [CrossRef] [Green Version]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef]
- Wright, G.A.; Softley, S.; Earnshaw, H. Low doses of neonicotinoid pesticides in food rewards impair short-term olfactory memory in foraging-age honeybees. Sci. Rep. 2015, 5, 15322. [Google Scholar] [CrossRef] [Green Version]
- Stanley, D.A.; Garratt, M.P.D.; Wickens, J.; Wickens, V.; Potts, S.G.; Raine, N. Neonicotinoid pesticide exposure impairs crop pollination services provided by bumblebees. Nature 2015, 528, 548–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klatt, B.K.; Rundlöf, M.; Smith, H.G. Maintaining the Restriction on neonicotinoids in the European union–benefits and risks to bees and pollination services. Front. Ecol. Evol. 2016, 4, 4. [Google Scholar] [CrossRef] [Green Version]
- Kathage, J.; Castañera, P.; Alonso-Prados, J.L.; Gómez-Barbero, M.; Rodríguez-Cerezo, E. The impact of restrictions on neonicotinoid and fipronil insecticides on pest management in maize, oilseed rape and sunflower in eight European Union regions. Pest. Manag. Sci. 2018, 74, 88–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Di Pasquale, G.; Salignon, M.; Le Conte, Y.; Belzunces, L.P.; Decourtye, A.; Kretzschmar, A.; Suchail, S.; Brunet, J.-L.; Alaux, C. Influence of pollen nutrition on honey bee health: Do pollen quality and diversity matter? PLoS ONE 2013, 8, e72016. [Google Scholar] [CrossRef] [Green Version]
- Winfree, R. The conservation and restoration of wild bees. Ann. N. Y. Acad. Sci. 2010, 1195, 169–197. [Google Scholar] [CrossRef]
- Willmer, P. Ecology: Pollinator–Plant synchrony tested by climate change. Curr. Biol. 2012, 22, R131–R132. [Google Scholar] [CrossRef] [Green Version]
- Iler, A.M.; Inouye, D.W.; Høye, T.T.; Miller-Rushing, A.J.; Burkle, L.A.; Johnston, E.B. Maintenance of temporal synchrony between syrphid flies and floral resources despite differential phenological responses to climate. Glob. Chang. Biol. 2013, 19, 2348–2359. [Google Scholar] [CrossRef]
- Benadi, G.; Hovestadt, T.; Poethke, H.-J.; Blüthgen, N. Specialization and phenological synchrony of plant-pollinator interactions along an altitudinal gradient. J. Anim. Ecol. 2013, 83, 639–650. [Google Scholar] [CrossRef]
- Rixen, C.; Wipf, S.; Frei, E.R.; Stöckli, V. Faster, higher, more? Past, present and future dynamics of alpine and arctic flora under climate change. Alp. Bot. 2014, 124, 77–79. [Google Scholar] [CrossRef] [Green Version]
- Elsen, P.; Tingley, M. Global mountain topography and the fate of montane species under climate change. Nat. Clim. Chang. 2015, 5, 772–776. [Google Scholar] [CrossRef]
- Inouye, D.W. Effects of climate change on alpine plants and their pollinators. Ann. N. Y. Acad. Sci. 2020, 1469, 26–37. [Google Scholar] [CrossRef]
- Engler, R.; Randin, C.F.; Thuiller, W.; Dullinger, S.; Zimmermann, N.E.; Araújo, M.B.; Pearman, P.B.; Le Lay, G.; Piedallu, C.; Albert, C.H.; et al. 21st century climate change threatens mountain flora unequally across Europe. Glob. Chang. Biol. 2011, 17, 2330–2341. [Google Scholar] [CrossRef]
- Pyke, G.H.; Thomson, J.D.; Inouye, D.W.; Miller, T.J. Effects of climate change on phenologies and distributions of bumble bees and the plants they visit. Ecosphere 2016, 7, e01267. [Google Scholar] [CrossRef] [Green Version]
- Biella, P.; Bogliani, G.; Cornalba, M.; Manino, A.; Neumayer, J.; Porporato, M.; Rasmont, P.; Milanesi, P. Distribution patterns of the cold adapted bumblebee Bombus alpinus in the Alps and hints of an uphill shift (Insecta: Hymenoptera: Apidae). J. Insect Conserv. 2017, 21, 357–366. [Google Scholar] [CrossRef]
- Dullinger, S.; Gattringer, A.; Thuiller, W.; Moser, D.; Zimmermann, N.; Guisan, A.; Willner, W.; Plutzar, C.; Leitner, M.; Mang, T.; et al. Extinction debt of high-mountain plants under twenty-first-century climate change. Nat. Clim. Chang. 2012, 2, 619–622. [Google Scholar] [CrossRef]
- Brown, J.H.; Lomolino, M.V. Biogeography; Sinauer Associates, Inc. Publishers: Sunderland, MA, USA, 1998. [Google Scholar]
- Cushman, J.H.; Lawton, J.H.; Manly, B.F.J. Latitudinal patterns in European ant assemblages: Variation in species richness and body size. Oecologia 1993, 95, 30–37. [Google Scholar] [CrossRef]
- Hawkins, B.A.; Lawton, J.H. Latitudinal gradients in butterfly body sizes: Is there a general pattern? Oecologia 1995, 102, 31–36. [Google Scholar] [CrossRef]
- Hawkins, B.A.; De Vries, P.J. Altitudinal gradients in the body sizes of Costa Rican Butterflies. Acta Oecol. 1996, 17, 185–194. [Google Scholar]
- Malo, J.E.; Baonza, J. Are there predictable clines in plant-pollinator interactions along altitudinal gradients? The example of Cytisus scoparius (L.) Link in the Sierra de Guadarrama (Central Spain). Divers. Distrib. 2002, 8, 365–371. [Google Scholar] [CrossRef]
- Arroyo, M.T.K.; Primack, R.; Armesto, J. Community studies in pollination ecology in the high temperate Andes of central Chile. I. pollination mechanisms and altitudinal variation. Am. J. Bot. 1982, 69, 82–97. [Google Scholar] [CrossRef]
- Inouye, D.W.; Pyke, G.H. Pollination biology in the Snowy Mountains of Australia: Comparisons with montane Colorado, USA. Austral. Ecol. 1988, 13, 191–205. [Google Scholar] [CrossRef]
- Warren, S.D.; Booth, G.M.; Harper, K.T. Elevational Distribution of Insect Pollinators. Am. Midl. Nat. 1988, 120, 325–330. [Google Scholar] [CrossRef]
- McCall, C.; Primack, R.B. Influence of flower characteristics, weather, time of day, and season on insect visitation rates in three plant communities. Am. J. Bot. 1992, 79, 434–442. [Google Scholar] [CrossRef]
- Kearns, C.A. Anthophilous fly distribution across an elevation gradient. Am. Midl. Nat. 1992, 127, 172–182. [Google Scholar] [CrossRef]
- Forister, M.L.; McCall, A.C.; Sanders, N.J.; Fordyce, J.A.; Thorne, J.H.; O’Brien, J.; Waetjen, D.P.; Shapiro, A.M. Compounded effects of climate change and habitat alteration shift patterns of butterfly diversity. Proc. Natl. Acad. Sci. USA 2010, 107, 2088–2092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Körner, C. The use of ‘altitude’ in ecological research. Trends Ecol. Evol. 2007, 22, 569–574. [Google Scholar] [CrossRef]
- Burgett, M.; Rucker, R.R.; Thurman, W.N. Economics and Honey Bee Pollination Markets. Am. Bee J. 2004, 144, 269–271. [Google Scholar]
- Calderone, N.W. Insect pollinated crops, insect pollinators and US agriculture: Trend analysis of aggregate data for the period 1992–2009. PLoS ONE 2012, 7, e37235. [Google Scholar] [CrossRef] [Green Version]
- Losey, J.E.; Vaughan, M. The economic value of ecological services provided by insects. BioScience 2006, 56, 311–323. [Google Scholar] [CrossRef] [Green Version]
- Galliot, J.-N.; Brunel, D.; Bérard, A.; Chauveau, A.; Blanchetête, A.; Lanore, L.; Farruggia, A. Investigating a flower-insect forager network in a mountain grassland community using pollen DNA barcoding. J. Insect Conserv. 2017, 21, 827–837. [Google Scholar] [CrossRef]
- Lucas, A.; Bodger, O.; Brosi, B.; Ford, C.R.; Forman, D.W.; Greig, C.; Hegarty, M.; Neyland, P.J.; de Vere, N. Generalisation and specialisation in hoverfly (Syrphidae) grassland pollen transport networks revealed by DNA metabarcoding. J. Anim. Ecol. 2018, 87, 1008–1021. [Google Scholar] [CrossRef] [Green Version]
- Lucas, A.; Bodger, O.; Brosi, B.J.; Ford, C.R.; Forman, D.W.; Greig, C.; Hegarty, M.; Jones, L.; Neyland, P.J.; de Vere, N. Floral resource partitioning by individuals within generalised hoverfly pollination networks revealed by DNA metabarcoding. Sci. Rep. 2018, 8, 5133. [Google Scholar] [CrossRef] [Green Version]
- Dunn, L.; Lequerica, M.; Reid, C.R.; Latty, T. Dual ecosystem services of syrphid flies (Diptera: Syrphidae): Pollinators and biological control agents. Pest. Manag. Sci. 2020, 76, 1973–1979. [Google Scholar] [CrossRef]
- Westhoff, V.; Van Der Maarel, E. The Braun-Blanquet Approach. In Classification of Plant Communities, 2nd ed.; Whittaker, R.H., Ed.; Springer: Dordrecht, The Netherlands, 1978; pp. 287–399. [Google Scholar]
- Westphal, C.; Bommarco, R.; Carré, G.; Lamborn, E.; Morison, N.; Petanidou, T.; Potts, S.G.; Roberts, S.P.M.; Szentgyörgyi, H.; Tscheulin, T.; et al. Measuring bee diversity in different European habitats and biogeographical regions. Ecol. Monogr. 2008, 78, 653–671. [Google Scholar] [CrossRef] [Green Version]
- Moreira, E.F.; Santos, R.L.D.S.; Penna, U.L.; Angel-Coca, C.; de Oliveira, F.F.; Viana, B.F. Are pan traps colors complementary to sample community of potential pollinator insects? J. Insect Conserv. 2016, 20, 583–596. [Google Scholar] [CrossRef]
- O’Connor, R.S.; Kunin, W.E.; Garratt, M.P.D.; Potts, S.G.; Roy, H.E.; Andrews, C.; Jones, C.M.; Peyton, J.M.; Savage, J.; Harvey, M.C.; et al. Monitoring insect pollinators and flower visitation: The effectiveness and feasibility of different survey methods. Methods Ecol. Evol. 2019, 10, 2129–2140. [Google Scholar] [CrossRef]
- Hutchinson, L.A.; Oliver, T.H.; Breeze, T.D.; O’Connor, R.S.; Potts, S.G.; Roberts, S.P.M.; Garratt, M.P.D. Inventorying and monitoring crop pollinating bees: Evaluating the effectiveness of common sampling methods. Insect Conserv. Divers. 2021, 2021, 12557. [Google Scholar] [CrossRef]
- Krahner, A.; Schmidt, J.; Maixner, M.; Porten, M.; Schmitt, T. Evaluation of four different methods for assessing bee diversity as ecological indicators of agro-ecosystems. Ecol. Indic. 2021, 125, 107573. [Google Scholar] [CrossRef]
- Prendergast, K.S.; Menz, M.H.M.; Dixon, K.W.; Bateman, P.W. The relative performance of sampling methods for native bees: An empirical test and review of the literature. Ecosphere 2020, 11, 03076. [Google Scholar] [CrossRef]
- Droege, S.; Tepedino, V.J.; Lebuhn, G.; Link, W.; Minckley, R.L.; Chen, Q.; Conrad, C. Spatial patterns of bee captures in North American bowl trapping surveys. Insect Conserv. Divers. 2010, 3, 15–23. [Google Scholar] [CrossRef]
- Barkalov, A.V.; Ståhls, G. Revision of the Palaearctic bare-eyed and black-legged species of the genus Cheilosia Meigen (Diptera, Syrphidae). Acta Zool. Fenn. 1997, 208, 1–74. [Google Scholar]
- Van Veen, M. Hoverflies of Northwest Europe: Identification Keys to the Syrphidae; KNNV Publishing: Utrecht, The Netherlands, 2010; 256p. [Google Scholar]
- Speight, M.C.D.; Sarthou, J.-P. StN keys for the identification of the European species of various genera of Syrphidae. In Syrph the Net, the Database of European Syrphidae (Diptera); Syrph the Net Publications: Dublin, Ireland, 2017; Volume 99, 139p. [Google Scholar]
- Intoppa, F.; Piazza, M.G.; Bolchi Serini, G.; Cornalba, M. I bombi. Guida al riconoscimento delle specie italiane. In CRA-Unità di Ricerca di Apicoltura e Bachicoltura; CSR: Roma, Italy, 2009; Volume 174. [Google Scholar]
- Michez, D.; Rasmont, P.; Terzo, M.; Vereecken, N.J. Bees of Europe. In Hymenoptera of Europe—1; N.A.P.: Verrières-le-Buisson, France, 2019; p. 545. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplifcation of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Villalta, I.; Ledet, R.; Baude, M.; Genoud, D.; Bouget, C.; Cornillon, M.; Moreau, S.; Courtial, B.; Lopez-Vaamonde, C. A DNA barcode-based survey of wild urban bees in the Loire Valley, France. Sci. Rep. 2021, 11, 4770. [Google Scholar] [CrossRef]
- Legendre, P.; Anderson, M.J. Distance-based redundancy analysis: Testing multispecies responses in multifactorial ecological experiments. Ecol. Monogr. 1999, 69, 1–24. [Google Scholar] [CrossRef]
- Legendre, P.; Gallagher, E.D. Ecologically meaningful transformations for ordination of species data. Oecologia 2001, 129, 271–280. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Package “Vegan”. 2020. Available online: https://cran.r-project.org/web/packages/vegan/vegan.pdf (accessed on 31 January 2022).
- Nekola, J.C.; White, P.S. The distance decay of similarity in biogeography and ecology. J. Biogeogr. 1999, 26, 867–878. [Google Scholar] [CrossRef] [Green Version]
- Andrade, J.; Estévez-Pérez, M. Statistical comparison of the slopes of two regression lines: A tutorial. Anal. Chim. Acta 2014, 838, 1–12. [Google Scholar] [CrossRef]
- Baselga, A. Partitioning the turnover and nestedness components of beta diversity. Glob. Ecol. Biogeogr. 2010, 19, 134–143. [Google Scholar] [CrossRef]
- Soininen, J.; Heino, J.; Wang, J. A meta-analysis of nestedness and turnover components of beta diversity across organisms and ecosystems. Glob. Ecol. Biogeogr. 2018, 27, 96–109. [Google Scholar] [CrossRef] [Green Version]
- Baselga, A.; Orme, C.D.L. Betapart: An R package for the study of beta diversity. Methods Ecol. Evol. 2012, 3, 808–812. [Google Scholar] [CrossRef]
- Harrell, F.E.; Dupont, C. Package “Hmisc”. 2021. Available online: https://cran.r-project.org/web/packages/Hmisc/Hmisc.pdf (accessed on 31 January 2022).
- Baselga, A.; Orme, D.; Villeger, S.; De Bortoli, J.; Leprieur, F.; Logez, M.; Henriques-Silva, R. Package “Betapart”. 2021. Available online: https://cran.r-project.org/web/packages/betapart/betapart.pdf (accessed on 15 January 2022).
- Wickham, H.; Chang, W.; Henry, L.; Pedersen, T.L.; Takahashi, K.; Wilke, C.; Woo, K. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA; Available online: http://ggplot2.tidyverse.org (accessed on 31 January 2022).
- Romig, T. Syrphidae—Part 2. In Diptera Stelviana: A Dipterological Perspective on a Changing Alpine Landscape; Ziegler, J., Ed.; Studia Dipterologica Supplement 21; Ampyx-Verlag: Halle/Salle, Germany, 2016; Volume 2. [Google Scholar]
- Speight, M.C.D.; Castella, E.; Obrdlik, P. Use of the Syrph the Net database 2000. In Syrph the Net, the Database of European Syrphidae; Syrph the Net Publications: Dublin, Ireland, 2000; Volume 25. [Google Scholar]
- Ssymank, A. Habitat use by Syrphidae (Diptera) in the valley of the river Strom—Part 2: Comparison of wet grassland and its succession stages. Volucella 2007, 8, 165–184. [Google Scholar]
- Hickman, J.M.; Wratten, S.D.; Jepson, P.C.; Frampton, C.M. Effect of hunger on yellow water trap catches of hoverfly (Diptera: Syrphidae) adults. Agric. For. Entomol. 2001, 3, 35–40. [Google Scholar] [CrossRef]
- Pei, C.K.; Hovick, T.J.; Duquette, C.A.; Limb, R.F.; Harmon, J.P.; Geaumont, B.A. Two common bee-sampling methods reflect different assemblages of the bee (Hymenoptera: Apoidea) community in mixed-grass prairie systems and are dependent on surrounding floral resource availability. J. Insect Conserv. 2022, 26, 69–83. [Google Scholar] [CrossRef]
- Rahbek, C. The elevational gradient of species richness: A uniform pattern? Ecography 1995, 18, 200–205. [Google Scholar] [CrossRef]
- Rahbek, C. The role of spatial scale and the perception of large-scale species-richness patterns. Ecol. Lett. 2005, 8, 224–239. [Google Scholar] [CrossRef]
- Colwell, R.K.; Gotelli, N.J.; Ashton, L.; Beck, J.; Brehm, G.; Fayle, T.M.; Fiedler, K.; Forister, M.L.; Kessler, M.; Kitching, R.L.; et al. Midpoint attractors and species richness: Modelling the interaction between environmental drivers and geometric constraints. Ecol. Lett. 2016, 19, 1009–1022. [Google Scholar] [CrossRef] [Green Version]
- Peters, M.; Hemp, A.; Appelhans, T.; Behler, C.; Classen, A.; Detsch, F.; Ensslin, A.; Ferger, S.W.; Frederiksen, S.B.; Gebert, F.; et al. Predictors of elevational biodiversity gradients change from single taxa to the multi-taxa community level. Nat. Commun. 2016, 7, 13736. [Google Scholar] [CrossRef] [Green Version]
- Widhiono, I.; Sudiana, E.; Darsono, D. Diversity of wild bees along elevational gradient in an agricultural area in central Java, Indonesia. Psyche A J. Entomol. 2017, 2017, 2968414. [Google Scholar] [CrossRef] [Green Version]
- Hackenberger, D.; Jarić, D.; Krcmar, S. Distribution of tabanids (Diptera: Tabanidae) along a two-sided altitudinal transect. Environ. Entomol. 2009, 38, 1600–1607. [Google Scholar] [CrossRef]
- Srisuka, W.; Sulin, C.; Aupalee, K.; Phankaen, T.; Taai, K.; Thongsahuan, S.; Saeung, A.; Takaoka, H. Community structure, biodiversity and spatiotemporal distribution of the black flies (Diptera: Simuliidae) using malaise traps on the highest mountain in Thailand. Insects 2021, 12, 504. [Google Scholar] [CrossRef]
- Słowińska, I.; Jaskuła, R. Distributional Patterns of Aquatic Empididae (Diptera) along an Elevational Diversity Gradient in a Low Mountain Range: An Example from Central Europe. Insects 2021, 12, 165. [Google Scholar] [CrossRef]
- Perillo, L.N.; Neves, F.; Antonini, Y.; Martins, R.P. Compositional changes in bee and wasp communities along Neotropical mountain altitudinal gradient. PLoS ONE 2017, 12, e0182054. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Longino, J.T.; Colwell, R.K.; O’Donnell, S. Elevational Patterns of Diversity and Abundance of Eusocial Paper Wasps (Vespidae) in Costa Rica. Biotropica 2009, 41, 338–346. [Google Scholar] [CrossRef]
- Finnie, S.; Sam, K.; Leponce, M.; Basset, Y.; Drew, D.; Schutze, M.K.; Dahl, C.; Damag, M.; Dilu, M.; Gewa, B.; et al. Assemblages of fruit flies (Diptera: Tephritidae) along an elevational gradient in the rainforests of Papua New Guinea. Insect Conserv. Divers. 2021, 14, 348–355. [Google Scholar] [CrossRef]
- Wolda, H. Altitude, habitat and tropical insect diversity. Biol. J. Linn. Soc. 1987, 30, 313–323. [Google Scholar] [CrossRef]
- McCoy, E.D. The Distribution of Insects along Elevational Gradients. Oikos 1990, 58, 313–322. [Google Scholar] [CrossRef]
- Beck, J.; Altermatt, F.; Hagmann, R.; Lang, S. Seasonality in the altitude–diversity pattern of Alpine moths. Basic Appl. Ecol. 2010, 11, 714–722. [Google Scholar] [CrossRef]
- Baumann, K.; Keune, J.; Wolters, V.; Jauker, F. Distribution and pollination services of wild bees and hoverflies along an altitudinal gradient in mountain hay meadows. Ecol. Evol. 2021, 11, 11345–11351. [Google Scholar] [CrossRef]
- Sanders, N.; Moss, J.; Wagner, D. Patterns of ant species richness along elevational gradients in an arid ecosystem. Glob. Ecol. Biogeogr. 2003, 12, 93–102. [Google Scholar] [CrossRef] [Green Version]
- Hodkinson, I.D. Terrestrial insects along elevation gradients: Species and community responses to altitude. Biol. Rev. 2005, 80, 489–513. [Google Scholar] [CrossRef] [Green Version]
- Rohde, K. Latitudinal gradients in species diversity: The search for the primary cause. Oikos 1992, 65, 514–527. [Google Scholar] [CrossRef] [Green Version]
- Rahbek, C.; Borregaard, M.K.; Colwell, R.K.; Dalsgaard, B.; Holt, B.G.; Morueta-Holme, N.; Nogues-Bravo, D.; Whittaker, R.J.; Fjeldså, J. Humboldt’s enigma: What causes global patterns of mountain biodiversity? Science 2019, 365, 1108–1113. [Google Scholar] [CrossRef]
- Hawkins, B.A.; Field, R.; Cornell, H.V.; Currie, D.J.; Guégan, J.-F.; Kaufman, D.M.; Kerr, J.T.; Mittelbach, G.G.; Oberdorff, T.; O’Brien, E.M.; et al. Energy, water, and broad-scale geographic patterns of species richness. Ecology 2003, 84, 3105–3117. [Google Scholar] [CrossRef] [Green Version]
- Hurlbert, A.H.; Stegen, J. When should species richness be energy limited, and how would we know? Ecol. Lett. 2014, 17, 401–413. [Google Scholar] [CrossRef]
- Belmaker, J.; Jetz, W. Relative roles of ecological and energetic constraints, diversification rates and region history on global species richness gradients. Ecol. Lett. 2015, 18, 563–571. [Google Scholar] [CrossRef]
- Novotny, V.; Drozd, P.; Miller, S.E.; Kulfan, M.; Janda, M.; Basset, Y.; Weiblen, G.D. Why are there so many species of herbivorous insects in tropical rainforests? Science 2006, 313, 1115–1118. [Google Scholar] [CrossRef] [Green Version]
- Romdal, T.S.; Grytnes, J.-A. An indirect area effect on elevational species richness patterns. Ecography 2007, 30, 440–448. [Google Scholar] [CrossRef]
- Brehm, G.; Colwell, R.K.; Kluge, J. The role of environment and mid-domain effect on moth species richness along a tropical elevational gradient. Glob. Ecol. Biogeogr. 2007, 16, 205–219. [Google Scholar] [CrossRef]
- Colwell, R.K.; Rahbek, C.; Gotelli, N.J. The mid-domain effect and species richness patterns: What have we learned so far? Am. Nat. 2004, 163, E1–E23. [Google Scholar] [CrossRef] [Green Version]
- Zapata, F.A.; Gaston, K.J.; Chown, S.L. Mid-domain models of species richness gradients: Assumptions, methods and evidence. J. Anim. Ecol. 2003, 72, 677–690. [Google Scholar] [CrossRef] [PubMed]
- Viterbi, R.; Cerrato, C.; Bassano, B.; Bionda, R.; Von Hardenberg, A.; Provenzale, A.; Bogliani, G. Patterns of biodiversity in the northwestern Italian Alps: A multi-taxa approach. Community Ecol. 2013, 14, 18–30. [Google Scholar] [CrossRef]
- Bishop, T.; Robertson, M.P.; van Rensburg, B.; Parr, C. Contrasting species and functional beta diversity in montane ant assemblages. J. Biogeogr. 2015, 42, 1776–1786. [Google Scholar] [CrossRef] [PubMed]
- Fontana, V.; Guariento, E.; Hilpold, A.; Niedrist, G.; Steinwandter, M.; Spitale, D.; Nascimbene, J.; Tappeiner, U.; Seeber, J. Species richness and beta diversity patterns of multiple taxa along an elevational gradient in pastured grasslands in the European Alps. Sci. Rep. 2020, 10, 12516. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot | Acronym | Gradient | Geographical Coordinates | Altitude (m) | Slope (°) | Type of Habitat (%) | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Wood | Grassl. | Scree | Scrub | Other | ||||||
| Pian d’Avena | VFA | Vette F. | 46.053101 11.841331 | 780 | 4 | 27 | 48 | 0 | 5 | 20 |
| Soladen | VFB | Vette F. | 46.060963 11.84648 | 956 | 5 | 3 | 90 | 0 | 7 | 0 |
| Camogne | VFC | Vette F. | 46.07009 11.853111 | 1210 | 26 | 1 | 72 | 0 | 27 | 0 |
| Camogne Alte | VFD | Vette F. | 46.073335 11.846008 | 1415 | 20 | 15 | 40 | 19 | 23 | 3 |
| Cimetta | VFE | Vette F. | 46.077279 11.842918 | 1635 | 17 | 38 | 42 | 0 | 17 | 3 |
| Col Cesta | VFF | Vette F. | 46.084766 11.844764 | 1790 | 35 | 13 | 40 | 15 | 26 | 6 |
| Rifugio Dal Piaz | VFG | Vette F. | 46.090258 11.841481 | 1990 | 15 | 0 | 97 | 2 | 0 | 1 |
| Vette Grandi | VFH | Vette F. | 46.09371 11.837018 | 2130 | 22 | 0 | 37 | 46 | 16 | 1 |
| Casera al Pos | MGA | Grave M. | 46.087296 11.939371 | 1075 | 7.5 | 54 | 27 | 0 | 16 | 3 |
| Al Pian | MGB | Grave M. | 46.090228 11.934328 | 1280 | 21 | 28 | 55 | 0 | 16 | 1 |
| Monte Grave | MGC | Grave M. | 46.091523 11.928041 | 1375 | 20 | 7 | 81 | 0 | 12 | 0 |
| Hoverflies | Bees | |||
|---|---|---|---|---|
| Abundance | Species Richness | Abundance | Species Richness | |
| Total | Quadratic R2 = 0.19 p = 0.042 * | Quadratic R2 = 0.385 p = 0.002 ** | Linear R2 = 0.241 p = 0.009 ** | Linear R2 = 0.137 p = 0.042 * |
| June | Quadratic R2 = 0.238 p = 0.22 | Quadratic R2 = 0.443 p = 0.1 | Linear R2 = 0.238 p = 0.12 | Quadratic R2 = 0.285 p = 0.19 |
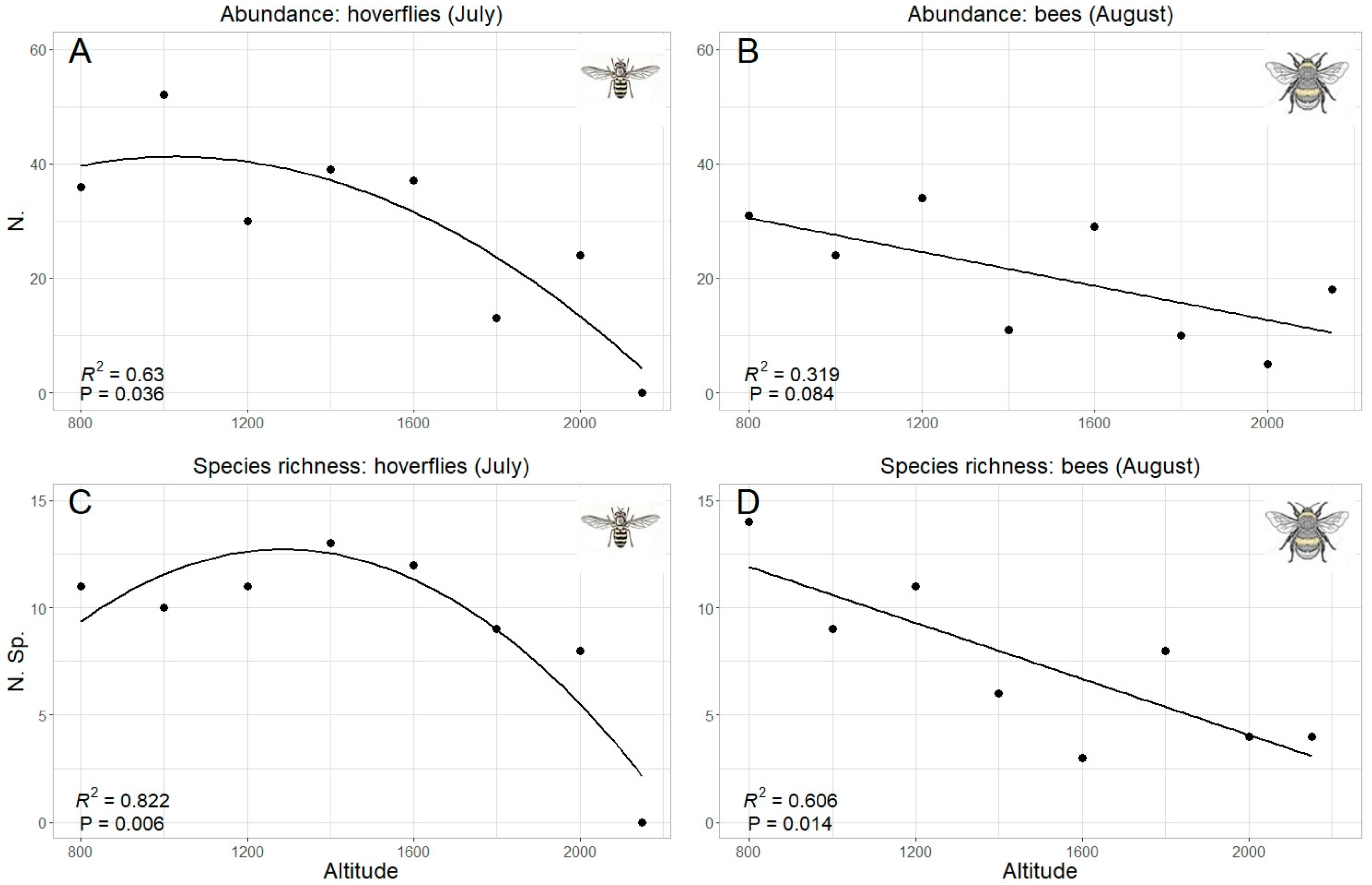
| July | Quadratic R2 = 0.614 p = 0.04 * | Quadratic R2 = 0.767 p = 0.011 * | Linear R2 = 0.234 p = 0.13 | Linear R2 = 0.288 p = 0.098 |
| August | Quadratic R2 = 0.07 p= 0.51 | Quadratic R2 = 0.16 p = 0.62 | Linear R2 = 0.335 p = 0.078 | Linear R2 = 0.606 p = 0.014 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sommaggio, D.; Zanotelli, L.; Vettorazzo, E.; Burgio, G.; Fontana, P. Different Distribution Patterns of Hoverflies (Diptera: Syrphidae) and Bees (Hymenoptera: Anthophila) Along Altitudinal Gradients in Dolomiti Bellunesi National Park (Italy). Insects 2022, 13, 293. https://doi.org/10.3390/insects13030293
Sommaggio D, Zanotelli L, Vettorazzo E, Burgio G, Fontana P. Different Distribution Patterns of Hoverflies (Diptera: Syrphidae) and Bees (Hymenoptera: Anthophila) Along Altitudinal Gradients in Dolomiti Bellunesi National Park (Italy). Insects. 2022; 13(3):293. https://doi.org/10.3390/insects13030293
Chicago/Turabian StyleSommaggio, Daniele, Livia Zanotelli, Enrico Vettorazzo, Giovanni Burgio, and Paolo Fontana. 2022. "Different Distribution Patterns of Hoverflies (Diptera: Syrphidae) and Bees (Hymenoptera: Anthophila) Along Altitudinal Gradients in Dolomiti Bellunesi National Park (Italy)" Insects 13, no. 3: 293. https://doi.org/10.3390/insects13030293
APA StyleSommaggio, D., Zanotelli, L., Vettorazzo, E., Burgio, G., & Fontana, P. (2022). Different Distribution Patterns of Hoverflies (Diptera: Syrphidae) and Bees (Hymenoptera: Anthophila) Along Altitudinal Gradients in Dolomiti Bellunesi National Park (Italy). Insects, 13(3), 293. https://doi.org/10.3390/insects13030293